
Abstract
We screened blood from 59 bats from French Guiana for Bartonella spp. PCRs were positive for 13.6% and culture was positive in one Noctilio albiventris and one Pteronotus parnellii, as well as in Ornithodoros hasei ticks collected from bats. Two isolated strains represent possible two new species.
Introduction
B
Materials and Methods
Blood samples were drawn from 59 apparently healthy bats (32 males and 27 females) in five areas near the coast of French Guiana: Saint-Jean-du-Maroni (05° 23′ 95″ N–54° 04′ 72″ W), Crique Malmanoury (05° 09′ 61″ N–52° 53′ 59″ W), Kourou (05° 13′ 96″ N–52° 45′ 19″ W), Cayenne (04° 54′ 65″ N–52° 18′ 54″ W), and Regina (04° 17′ 93″ N–52° 11′ 88″ W). Bats were trapped in January and February 2013 with hand nets (n = 16), mist nets (n = 26), or by hand (n = 17). Catches occurred in residential or unoccupied buildings, in culverts beneath a road, under bridges, or in caves. The species, age, sex, reproductive status, and morphological measurements of the bats were recorded. The protocol for capturing and sampling bat specimens (N°1688) was approved by the Animal Ethics Committee of Marseille (C2EA-14) and by the French authorities.
Blood samples were collected in dry tubes by intracardiac puncture of anesthetized bats (ketamine—10 mg/kg—intraperitoneal). After centrifugation, the serum was separated. The samples (cloths and sera) were kept refrigerated at +4°C during transportation to the laboratory. The DNA was extracted from 200 μL of mechanically disintegrated cloth and from ticks by the BioRobot EZ1 (QIAGEN, Courtaboeuf, France) device with a commercial DNA Extraction Kit (QIAamp DNA Mini Kit®) according to the manufacturer's instructions. All samples were screened using a Bartonella genus-specific real-time PCR with a 21-bp probe targeting the 16S–23S RNA intergenic spacer (Varagnol et al. 2009). The negative controls used consisted of DNA extracted from cell culture (L929) and distilled water. Positive controls included samples of Bartonella elizabethae (ATCC 49927) DNA.
All blood samples were plated on Columbia agar (BioMerieux, Marcy l'Etoile, France) and incubated at 37°C in a 5% CO2-enriched atmosphere for 6 weeks or until growth occurred as previously described (La Scola et al. 2003). Bacterial colonies were identified as Bartonella spp. based on their morphology. Conventional PCR (ITS, rpoB, ftsZ, gltA, and 16S) was performed on the resulting colonies to characterize isolated strains (La Scola et al. 2003). All obtained amplicons were directly sequenced; data were collected with an ABI PRISM 3130xl Genetic Analyzer capillary sequencer (Applied Biosystems, Foster City, CA). Sequences were edited and assembled using ChromasPro 1.34 (Technelysium Pty. Ltd., Tewantin, Australia). A BLAST search was performed for identification of the sequences obtained.
The Bayesian phylogenetic analysis (Ronquist and Huelsenbeck 2003) was performed on the TOPALi 2.5 software (Biomathematics and Statistics Scotland) with the integrated MrBayes application (
Results
In total, 59 bats from seven different species (seven genera) were captured; the most numerous were Noctilio albiventris (n = 22) and Pteronotus parnellii (n = 14) (Table 1). There were 11 immature bats, 48 adults, and 1 pregnant bat.
In total, 355 larvae from soft ticks Ornithodoros hasei taken from 12 bats (N. albiventris) were collected and samples from 105 ticks were analyzed. The ticks from the bats were removed and kept in 70% ethanol.
We detected Bartonella DNA in the blood of eight bats (13.6%, 8/59: 5 P. parnellii, 2 N. albiventris, and 1 Eumops auripendulus) and bacterial culture was positive in 2 insectivore bats: 1 N. albiventris (1 male/22—4.5%) and 1 P. parnellii (1 female/14—7.1%). Positive samples came from the area of Saint-Jean-du-Maroni and Malmanoury, west of Cayenne, respectively. Four O. hasei ticks from 4 N. albiventris captured in Saint-Jean-du-Maroni were also positive for Bartonella spp. We did not succeed in amplification of Bartonella ITS from these samples. PCR was performed on blood from one of these four bats and the results were negative.
We succeeded in isolating two bartonella strains from the blood of two adult bats: one (strain C65) from male N. albiventris and one (strain C102) from female P. parnellii. Morphologically, the colonies from both strains were consistent with other representatives of the genus Bartonella. However, four sequenced genes and ITS showed that neither of the two isolated strains belonged to a known Bartonella species (Table 2).
Based on 276-bp long homological portions of ITS out of total length 420-bp.
Based on 265-bp long homological portions of ITS out of total length 724-bp.
Discussion
In 2003, La Scola et al. proposed a multilocus sequence analysis based on four genes (16S rRNA, gltA, ftsZ, and rpoB) and one intergenic spacer (ITS) as a tool for the description of new Bartonella species (La Scola et al. 2003). Among these genetic markers, two, that is, gltA and rpoB, were particularly discriminatory, with new Bartonella isolates considered new species if they exhibited <96.0% and <95.4% sequence identity with other validly published species for the 327- and 825-bp fragments of the gltA and rpoB genes, respectively. According to these criteria and to the position of the two strains on the phylogenetic tree (Fig. 1), we suggest that both isolates may represent new species. Taking into consideration the fact that these strains are not yet completely described, we propose provisional names for these strains: Candidatus Bartonella rolaini for strain C65 and Candidatus Bartonella lascolai for strain C102 (Murray and Stackebrandt 1995).
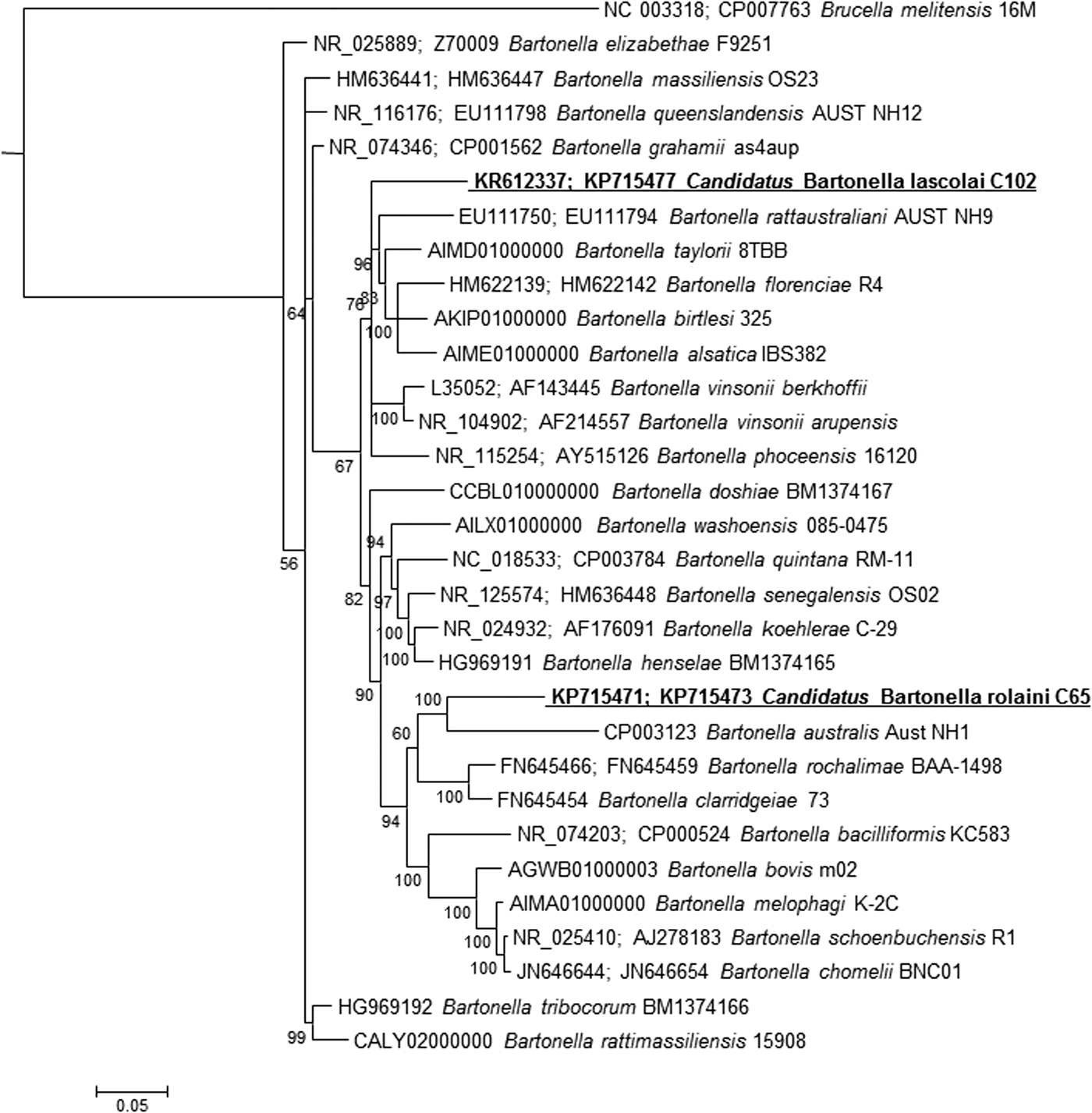
A consensus phylogenetic tree showing the relationships of the studied species of Bartonella based on concatenated 16S and gltA gene sequence comparisons. GenBank accession numbers (or the only genome accession number) are indicated when the sequences originated from GenBank (16S; gltA). The sequences were aligned using ClustalW, and phylogenetic inferences were obtained using Bayesian phylogenetic analysis with TOPALi 2.5 software (Biomathematics and Statistics Scotland, Edinburgh, United Kingdom) within the integrated MrBayes application using the SYM substitution model. Numbers at the nodes are percentages of bootstrap values obtained by repeating the analysis 100 times to generate a majority consensus tree. The final set includes 2301-bp (1410 and 891 for 16S and gltA, respectively). The scale bar represents a 5% nucleotide sequence divergence.
Obtained sequences were submitted to the GenBank under the following accession numbers: KP715471–KP715475 for 16S, ftsZ, gltA, ITS, and rpoB de Bartonella sp. C65 and KP715476–KP715478 and KR612337 for ftsZ, gltA, ITS, and 16S of Bartonella sp. C102.
The new isolates described in our study are the first strains of Bartonella spp. bats from French Guiana. Bats are frequently parasitized by Nycteribiidae flies. All Bartonella are vector-borne hemoparasites; they are often detected or isolated from Nycteribiidae flies (Morse et al. 2012), suggesting that the bat flies are vectors of Bartonella species among bats. In our case, however, the bats were not parasitized by Nycteribiidae flies, and only soft ticks were found as ectoparasites. Bat-associated soft ticks in Europe were recently shown to harbor different presumably pathogenic bacteria such as Rickettsia, Borrelia, and Ehrlichia (Socolovschi et al. 2012). Soft ticks (Ornithodoros sonrai) from rodents in Senegal have already been shown to be Bartonella hosts with several strains recently described (Mediannikov et al. 2014). We have found Bartonella in ticks collected from bat that were Bartonella negative. It may be partially explained by the mobility of ticks from one host bat to another and by the possible fluctuations in Bartonella-induced bacteremia in bats. Our observations may, however, suggest the potential role of bat-associated soft ticks as Bartonella spp. vectors.
In our study, the prevalence of Bartonella-infected bats (13.6%—8/59) is greater than that described in Taiwan (11.1%—6/54), but less than those reported in Peru (24.1%—27/112), Kenya (30.2%—106/331), Guatemala (33%—39/118), Costa Rica (33.3%), Vietnam (35%—21/60), and Finland (37%—46/124) (Kosoy et al. 2010, Bai et al. 2011, 2012, Lin et al. 2012, Anh et al. 2015, Judson et al. 2015, Lilley et al. 2015). It appears that bacteremia is common in bats and depending on the species Bartonella spp. is adapted to its hosts in that it often does not cause symptoms. Our study shows for the first time that Noctilio spp. and Pteronotus spp. are carriers of Bartonella. In Kenya, 6 of 13 bat species were bacteremic and carriers of Bartonella spp. (Kosoy et al. 2010). In the Amazon in Peru, vampires (Desmodus rotundus) were found to be positive (Bai et al. 2012).
Conclusion
The pathogenicity of isolated strains of Bartonella in humans or other mammals is unknown, although captured bats were not apparently ill. The impact of Bartonella spp. in bats is also so far unknown. According to our results, insectivore bats play a possible role in the maintenance of Bartonella spp. in the environment and its soft ticks may play a role as vectors. Epidemiological surveillance of Bartonella in bats and their ectoparasites is required to better identify their role.
Footnotes
Acknowledgments
The authors thank the Foundation Méditerranée Infection for funding this study and the French Forces Medical Service for its support. The authors also thank G. Hyvert for his excellent assistance during the field work, and G. Ganay and C. Grimaldier for processing the samples.
Author Disclosure Statement
No competing financial interests exist.