
Abstract
Bluetongue virus (BTV) is an orbivirus transmitted by biting midges (Culicoides spp.) that can result in moderate to high morbidity and mortality primarily in sheep and white-tailed deer. Although only 5 serotypes of BTV are considered endemic to the United States, as many as 11 incursive serotypes have been detected in livestock and wildlife in the past 16 years. Introductions of serotypes, with unknown virulence and disease risk, are constant threats to US agriculture. One potential incursive serotype of particular concern is the European strain of BTV-8, which was introduced into Northern Europe in 2006 and caused unprecedented livestock disease and mortality during the 2006–2007 vector seasons. To assess disease risk of BTV-8 in a common white-faced American sheep breed, eight Polled Dorset yearlings were experimentally infected and monitored for clinical signs. Viremia and viral tissue distribution were detected and quantified by real-time qRT-PCR. Overall, clinical disease was moderate with no mortality. Viremia reached as high as 9.7 log10 particles/mL and persisted at 5 logs or higher through the end of the study (28 days). Virus distribution in tissues was extensive with the highest mean titers at the peak of viremia (day 8) in the kidney (8.38 log10 particles/mg) and pancreas (8.37 log10 particles/mg). Virus persisted in tissues of some sheep at 8 logs or higher by day 28. Results of this study suggest that should BTV-8 emerge in the United States, clinical disease in this common sheep breed would likely be similar in form, duration, and severity to what is typically observed in severe outbreaks of endemic serotypes, not the extraordinary disease levels seen in Northern Europe. In addition, a majority of exposed sheep would be expected to survive and act as significant BTV-8 reservoirs with high titer viremias for subsequent transmission to other livestock and wildlife populations.
Introduction
B
Five serotypes of BTV are considered endemic (domestic) in the United States (BTV-2, 10, 11, 13, 17); however, 11 incursive (exotic) serotypes have been detected, some repeatedly, in wildlife and livestock since 1999 (Ostlund 2015). This suggests introductions are relatively routine and consistent in North America and clearly illustrates the ability of this virus to successfully spread to new ecosystems and become endemic where susceptible animal populations and competent Culicoides spp. vectors exist. One potential incursive serotype of particular concern to the U.S. livestock industry, in terms of clinical disease threat and possible need for vaccine stockpiling, is BTV-8. In 2006, this virus emerged in previously BTV-free Northern Europe and, subsequently, overwintered, causing unprecedented livestock disease and mortality during the 2006–2007 Culicoides vector seasons (Elbers et al. 2008b; Wilson and Mellor 2009). It is not clear whether the rapid spread, disease severity, and mortality rates were due to its inherent virulence or the immunologically naive status of livestock populations or a combination of the two. A massive vaccine campaign, costing the European Union €231 million in 2008–2009 (Pinior et al. 2015), was thought to have successfully eradicated this serotype from the livestock population. However, as expected, cessation of vaccine requirements in 2010 and herd population turnover have resulted in a reemergence of BTV-8 in livestock (BVA 2015b), possibly from persisting virus in wildlife (BVA 2015a; Sailleau et al. 2015).
To predict outbreak severity for a specific incursive serotype, it is important to ascertain its virulence in the most susceptible, most widely distributed, domestic ruminant species in the United States, namely sheep. The American Polled Dorset sheep breed arose from the horned Dorset originally from England in 1860 and was registered as a distinct breed in 1956. This white-faced breed has become the second largest breed in total numbers in the U.S. sheep industry (OSU, 1997). They are, therefore, an economically important sentinel animal in bluetongue outbreak risk assessments. Considering the threat posed by incursion of BTV-8 into the United States, we sought to assess the disease severity of a potential BTV-8 outbreak in this common North American livestock sheep breed by assessing infection, viremia, and clinical disease following experimental inoculation of virus.
Materials and Methods
Virus inoculum
Virus (BTV-8/NET2007/01) was obtained from the Central Veterinary Institute of Wageningen University (Lelystad, The Netherlands). The BTV-8 inoculum was EDTA-blood harvested from a Holstein Frisian cow NL441689187 from Bavel, representing the first detected case of BTV-8 after overwintering (GenBank acc. no. GQ506451–GQ506460) (Backx et al. 2007). Due to rapid attenuation of the virus when passed in cells, qRT-PCR (Wilson et al. 2009) was previously used to determine a titer of 6 × 105 viral particles/mL for this unpassaged washed blood cell inoculum (Drolet et al. 2013).
Animal experiment
Eight female American Polled Dorset yearling sheep were sedated with xylazine (0.05–0.1 mg/kg) and injected with 1 mL BTV-8/NET2007/01 (6 × 105 virus particles/mL) bovine blood stock virus in the neck and inner left leg. A total of 10 0.1 mL intradermal injections were given in a 2.5 × 2.5 cm area for each of the two sites for a total per animal dose of 1.2 × 106 (6.08 log10) virus particles. Two negative control sheep were sham inoculated with BTV-negative bovine blood and housed with infected animals to examine direct contact transmission. On days 0–10, 14, 17, 21, 24, and 28, blood samples were taken for ELISA and qRT-PCR analysis, rectal temperatures were recorded, and clinical signs were scored using a scale from 0 to 3 (0 = absence; 1 = mild; 2 = moderate; 3 = severe). To examine viral distribution and tissue pathology at the anticipated peak of viremia (day 8), two animals were sedated as above and euthanized with pentobarbital (IV). Gross pathological findings were noted and samples of adrenal gland, lymph nodes (mesenteric, mandibular, popliteal, inguinal), pancreas, kidney, heart, liver, lung, intestine, and spleen were frozen for qRT-PCR testing or fixed in 10% buffered formalin (Sigma-Aldrich), embedded in paraffin wax, and sectioned (5 μm) for histological analysis. All animal work was performed under BSL-3 containment conditions at Colorado State University (Fort Collins, CO) in compliance with Institutional Animal Care and Use Committee guidelines.
Real-time qRT-PCR
Due to the inherent and rapid reduction in titer of BTV-8 when cultured, qRT-PCR is a more robust and repeatable assay for detection and quantitation of virus in the original inoculum, as well as in blood and tissue samples of infected animals (Chatzinasiou et al. 2010; Drolet et al. 2013; Veronesi et al. 2013). Viral RNA was extracted from blood using the MagMAX™ Blood RNA Isolation Kit (Ambion). For tissue RNA, 50 mg was homogenized in a microfuge tube by bead beating (TissueLyser; QIAGEN) in 500 μL PBS with two 4.5 mm stainless steel balls (MSC Industrial Supply). Total RNA was extracted from homogenates using the Total Viral RNA Isolation Kit (Ambion) and stored in 96-well plates at −80°C.
Blood and tissue RNA samples were tested in triplicate by qRT-PCR (Applied Biosystems 7500) as previously described (Wilson et al. 2009; Drolet et al. 2013). Viral RNA was detected using primer sequences specific for BTV Seg-5 (NS1) and Seg-10 (NS3) genes (Drolet et al. 2013), with β-actin as an internal control (Moniwa et al. 2007). The mean cycle threshold (Ct) values (triplicates) were compared to known titers of BTV stock virus to determine RNA concentrations. Virus particles estimated from RNA concentrations were calculated as previously described (Akita et al. 1992). Blood and tissue titers are reported as log10 virus particles/mL and log10 viral particles/mg, respectively.
Detection of infectious virus in blood and tissues
To confirm infectious virus in PCR-positive blood and tissue homogenates, cell culture flasks were inoculated and monitored for cytopathic effect (CPE). Day 8 blood samples from each sheep (1 mL each) were sonicated (Q700; Qsonica) for 3 min with 5 s pulses at 100 mA and centrifuged for 10 min at 10,000× g. Cleared supernatants (500 μL) were used to inoculate T-25 flasks (Corning) containing baby hamster kidney (BHK) cell monolayers grown in Dulbecco's modified Eagle's medium (DMEM) with 10% FBS, 100 U/mL penicillin, and 100 μg/mL streptomycin sulfate (Sigma-Aldrich). Flasks were incubated at 37°C for 3–5 days and examined for CPE. To determine the presence of infectious virus in blood and tissues at the end of the study (day 28), blood samples from the six surviving sheep, and 100 μL of cleared tissue homogenate supernatants from one randomly chosen animal (sheep 1), were similarly tested in CPE culture flasks.
For genetic comparison of replicating virus with the original BTV-8 inoculum, RNA was extracted from 1 mL aliquots of each day 8 CPE culture flask for qRT-PCR as above. In addition, viral RNA from the blood of one animal at day 8 (sheep 3) was used for VP2 gene sequencing to compare with the original inoculum (Maan et al. 2010; van Gennip et al. 2012). Four forward and three reverse primers were used for full VP2 gene coverage as previously described (Drolet et al. 2013).
Serology
Sheep were monitored for seroconversion by testing serum samples (days 0–10, 14, 17, 21, 24, and 28) for BTV antibody using a commercial competitive ELISA (Veterinary Diagnostic Technology). Optical densities (OD) were measured using a BioTek Synergy H4 Hybrid plate reader and used to calculate percent inhibition: 100-(OD490nm test serum/OD490nm internal negative control serum) × 100. Preinfection sera and diluent-only wells were used to determine negative serum values. Internal strong positive and weak positive kit controls were used to validate the assay.
Results
Clinical observations
Clinical signs of bluetongue disease in sheep were recorded on days 1–10, 14, 17, 21, 24, and 28. Febrile responses, as detected by rectal temperatures, peaked for most sheep from 7 to 9 days post infection (dpi), 1 to 3 days following initial detection of viremia (Fig. 1). Clinical signs (Fig. 2) were observed 7 to 14 dpi and included conjunctivitis, ocular and nasal discharge, facial edema, oral ulcers, excessive salivation, coronitis, diarrhea, and respiratory distress. At the peak of clinical disease (day 8), the most commonly observed and highest scoring clinical signs were ocular and nasal discharge, followed by conjunctivitis and facial edema (Fig. 3). Because sheep were housed five per room with communal food and water, individual scores for loss of appetite were not possible, but were noted in both rooms on days 8 and 9. Depression was also seen and was characterized by listlessness, lowered heads, and slow or diminished avoidance of human contact. Clinical signs of disease were not observed in the two sham inoculated negative control sheep housed with the inoculated sheep.

Febrile response (°C) as measured by rectal thermometer (red columns) and blood titers (virus particles log10/mL) as quantitated by real-time qRT-PCR (blue columns) in American Polled Dorset sheep inoculated with European BTV-8.
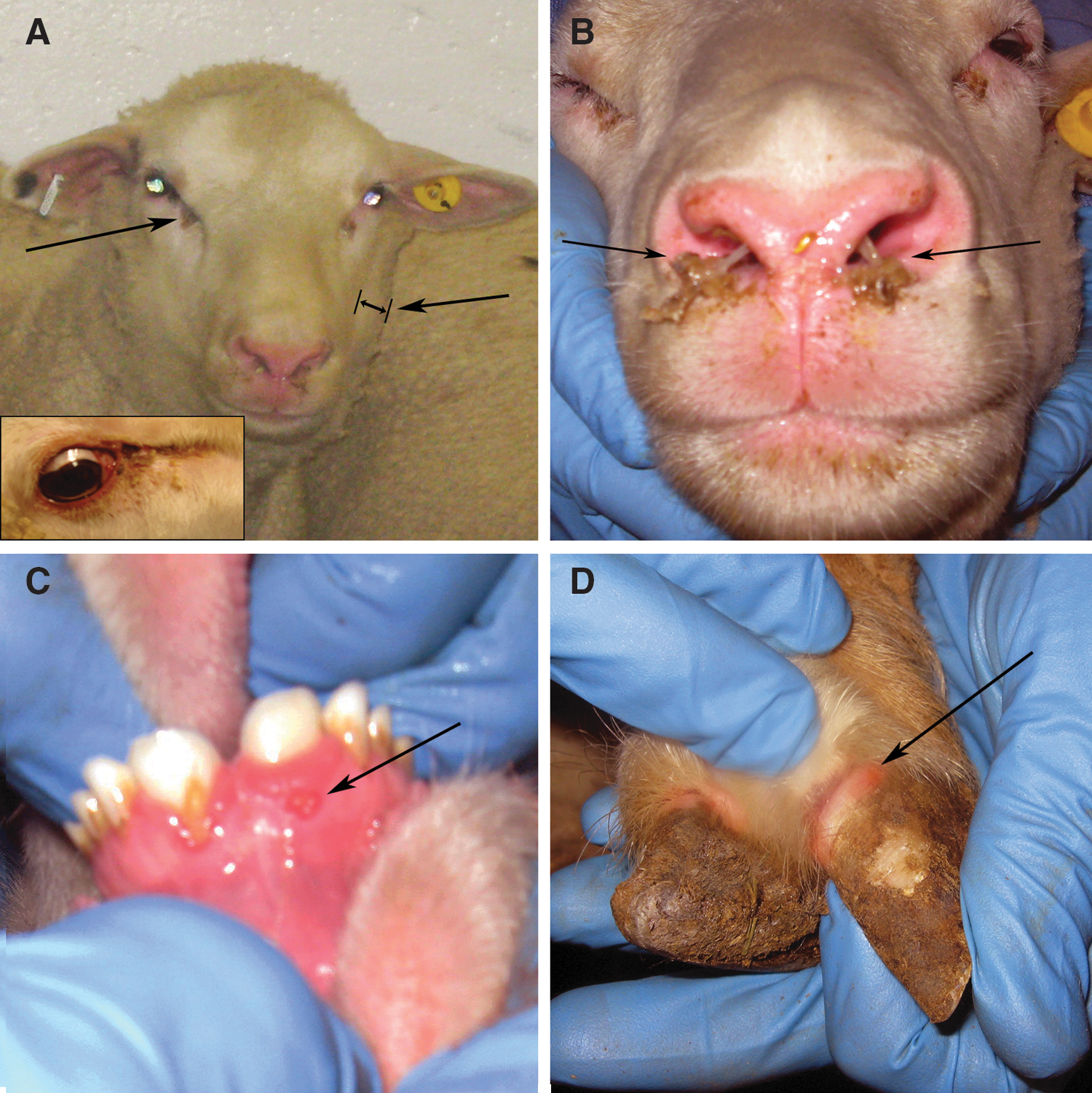
Clinical disease in American Polled Dorset sheep inoculated with BTV-8. Arrows indicate:
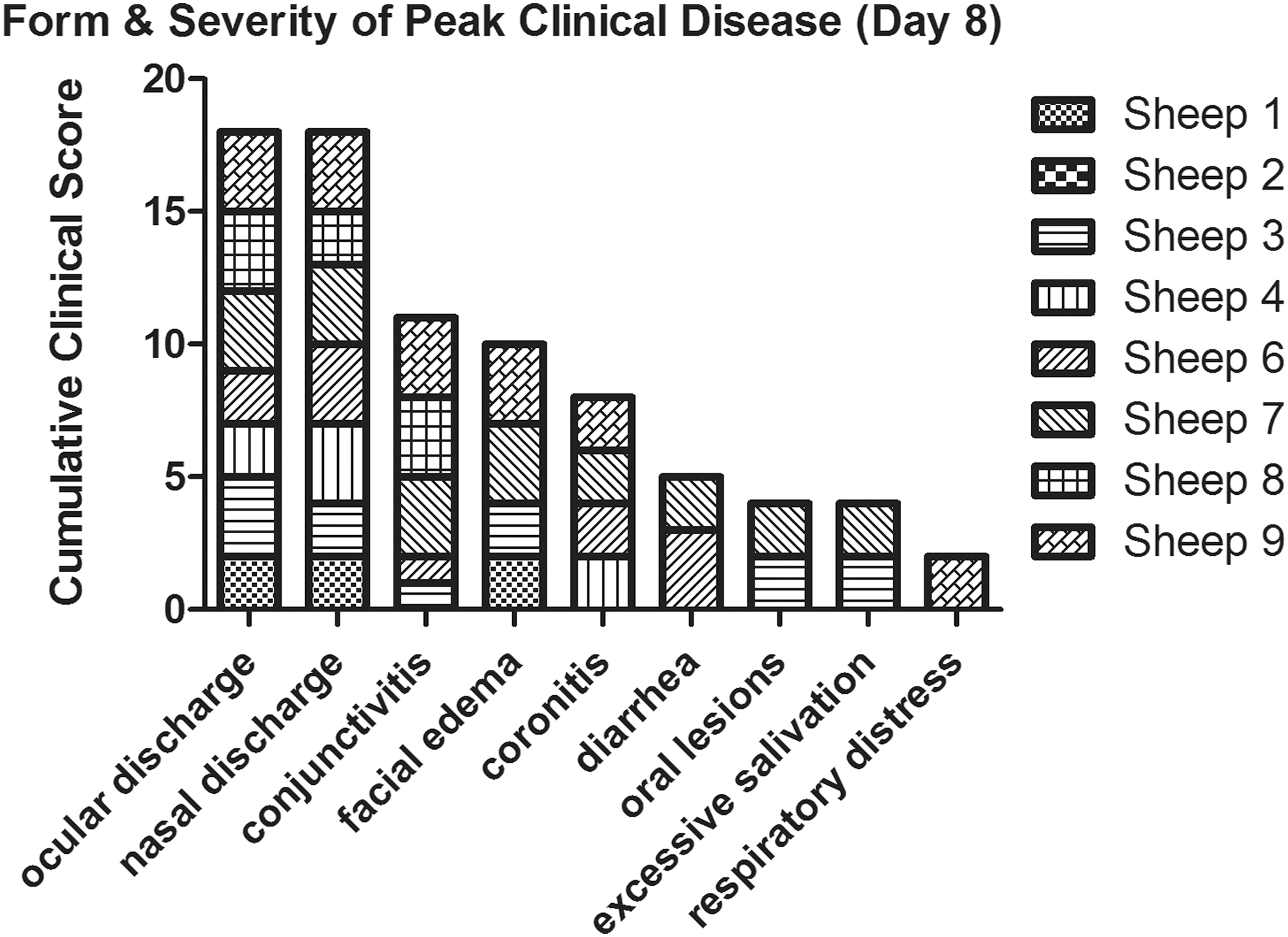
Clinical disease scores in sheep inoculated with BTV-8 at the peak of clinical disease (day 8). Individual clinical scores are shown cumulatively for each clinical sign observed.
Molecular analysis
Viremia in inoculated sheep was detected by real-time qRT-PCR by 6 or 7 dpi and peaked from 7 to 14 dpi (Fig. 1) reaching as high as 9.7 log10 virus particles/mL (sheep 2). At the end of the study (day 28), all sheep remained viremic with titers ranging from 5.3 to 6.9 log10 virus particles/mL (Fig. 1B). Analysis of necropsy tissue samples showed widespread dissemination of virus. In the day 8 sheep, virus was detected in all tissues tested (Fig. 4A) with the highest titers seen in the inguinal lymph node (draining node closest to the inner leg inoculation site) at 7.08 log10 particles/mg. At 28 dpi, virus was detected in all tissues except the inguinal lymph nodes with the highest titers seen in kidney (9.45 logs; sheep 1), lung (9.29 logs; sheep 1), and mandibular lymph node (8.67 logs; sheep 8) (Fig. 4B). No virus was detected in the blood or tissues of the negative control sheep, confirming the inability of this virus to transmit by direct contact.
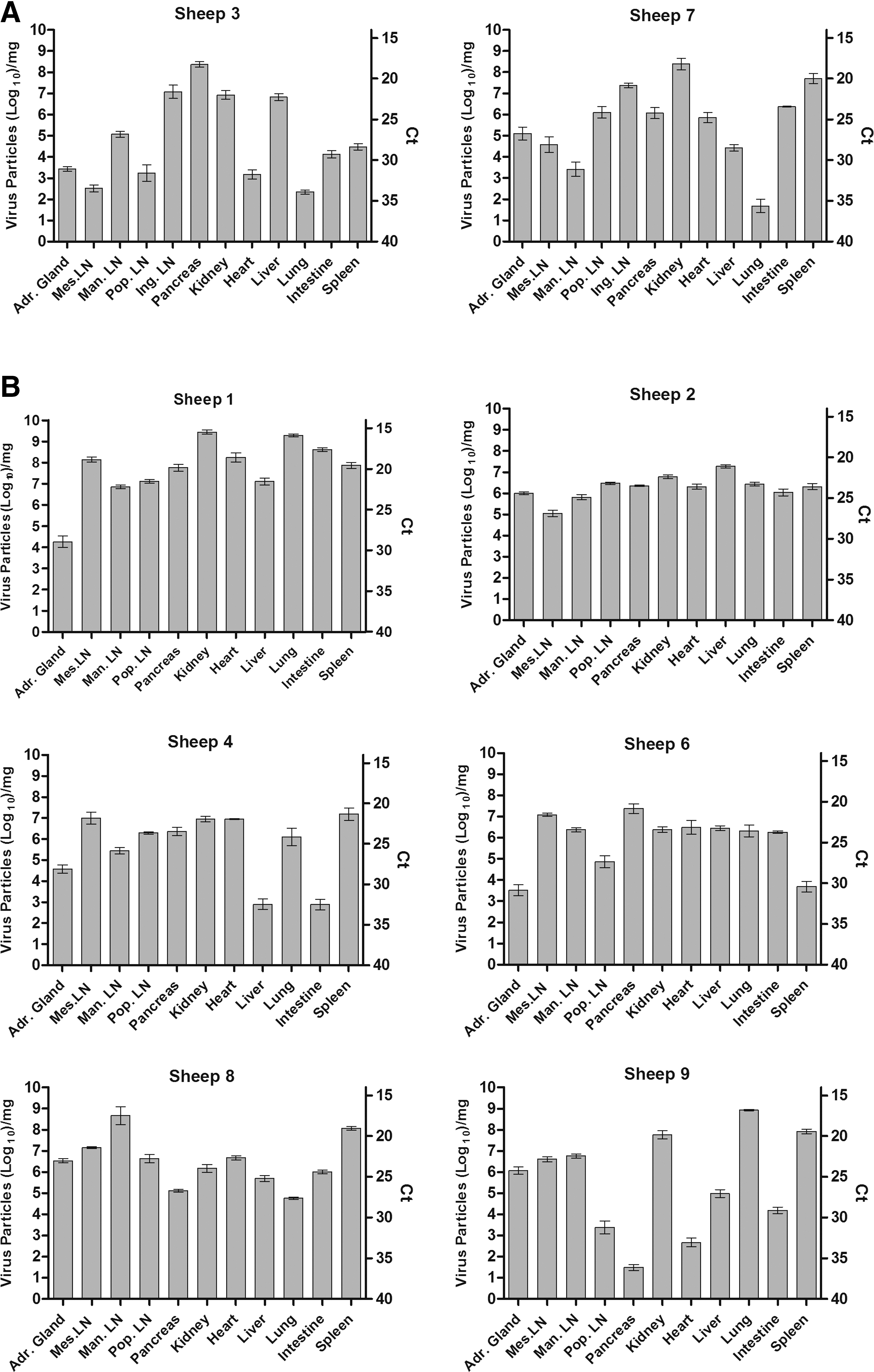
Virus distribution in tissues of American Polled Dorset sheep following BTV-8 inoculation.
Virus isolation
All BHK flasks inoculated with blood samples collected at 8 dpi were CPE positive after 3–5 days indicating the presence of infectious virus. Virus was confirmed as BTV-8 by real-time qRT-PCR of the VP2 gene (data not shown). Circulating virus in sheep was confirmed specifically as that of the original inoculum by sequencing the entire VP2 gene from the blood of sheep 3 on day 8. This gene confers serotype identity and has the most variable genomic region among the serotypes (Roy 2001). The 2841 nucleotide VP2 gene sequence showed 99.5% identity (100% similarity) to the original BTV-8 inoculum (data not shown). All BHK flasks inoculated with blood from the six remaining sheep on day 28 were CPE positive, confirming infectious viremia in all sheep at the end of the study. In addition, all of the PCR-positive tissues of one randomly chosen animal (sheep 1) were CPE positive, except the liver.
Serological analysis
Sera from three of eight sheep were positive for BTV antibody by cELISA at 9 dpi, and seven of eight were positive by 14 dpi (data not shown). All animals were seropositive for BTV by the end of the experiment. The two negative control animals, housed with inoculated animals, remained BTV antibody negative throughout the study.
Gross and microscopic pathology
In sheep euthanized at the anticipated peak of viremia and clinical disease (8 dpi; sheep 3 and 7), gross pathology was mild, but consistent with BTV infection and included hemorrhages in the spleen and mild accumulation of fluid in the pericardium. Although microscopic lesion characteristic of bluetongue disease was not identified at this early point in infection, other inconsequential background lesions were present, including lymphocytic portal hepatitis in the liver, mild medullary mineralization in the kidney, and mild eosinophilic bronchiolitis in the lung. In the six sheep euthanized at the end of the experiment (28 dpi), mild to moderate histopathological findings consistent with BTV infection (MacLachlan et al. 2009) were identified, including marked lymphoid hyperplasia with increased lymphocytic apoptosis in the spleen and tonsil, as well as multifocal cardiomyocyte degeneration and necrosis with mineralization, lymphocytic myocarditis, multifocal acute suppurative myocarditis, and mild lymphocytic arteritis with endothelial and perivascular proliferation in the heart. Similar to 8 dpi, other microscopic findings were also identified in these sheep, including mild medullary mineralization with tubular degeneration in the kidney, hepatic lipidosis, lymphocytic portal hepatitis, and mild multifocal suppurative hepatitis in the liver, as well as mild suppurative or eosinophilic bronchopneumonia in the lung.
Discussion
The presence of BTV-competent vector species and susceptible hosts throughout the United States results in a highly favorable environment for new serotypes to become established once introduced. Evidence of this has been seen in the repeated isolation of several incursive serotypes since the first confirmed detection (Ostlund 2015). Clearly, during any given vector season, the expected severity of a bluetongue outbreak must take into consideration not only endemic serotypes, with known outbreak histories and at least the potential for regional herd immunity, but also newly invasive serotypes of unknown virulence in herds with naive immune status.
The incursion of BTV-8 into Northern Europe and the UK has highlighted the risks that countries face with regard to this transboundary disease. The levels of morbidity and mortality in sheep, the overwintering, the transplacental transmission, and the rapid geographic spread were unprecedented (De Clercq et al. 2008; Elbers et al. 2008a, 2008b; Saegerman et al. 2011; van der Sluijs et al. 2011). Consequently, the U.S. sheep industry is greatly concerned about the potential for a similar introduction of BTV-8, resulting in an equally devastating outbreak, and speculation of whether vaccine stockpiling is warranted.
While the general susceptibility of American sheep to circulating endemic serotypes of BTV is well established, virulence can be highly variable between serotypes. Thus, it was not clear whether BTV-8 would result in the severe disease and mortality seen in Europe, or disease levels similar to our endemic serotypes. As sheep are the livestock species most often affected by reemerging endemic serotypes, they would be the sentinel livestock species and would play an important role as amplifying reservoirs for BTV-8 to become established in the new ecosystem. Understanding the degree to which our most susceptible livestock species is vulnerable to BTV-8 is a critical epidemiological component in determining complete and relevant disease risk analyses for the US livestock industry.
It is understood that clinical disease resulting from experimental infections can be affected by breed, age, dose, and inoculation route, making comparisons between studies difficult (MacLachlan et al. 2009). That said, clinical disease of sheep in this study was less severe, with significantly less pathology than was reported for English Dorset sheep (est. 1950, Australia) inoculated intradermally and subcutaneously with a 2006 Netherlands BTV-8 isolate (Darpel et al. 2007) or a 2007 German BTV-8 isolate (Worwa et al. 2010), as well as those reported in Merino sheep subcutaneously inoculated with a Belgian 2006 BTV-8 isolate (Sanchez-Cordon et al. 2013). In this study, viremia, seroconversion, viral tissue distribution, and clinical disease were generally similar to what is typically seen with experimental and natural infections of US endemic BTV serotypes (Ghalib et al. 1985; Foster et al. 1991; Verwoerd and Erasmus 2004).
Repeated exposure to multiple circulating serotypes in any given vector season can play a role in moderating the susceptibility to, and severity of, disease in certain regions when those serotypes reemerge. This was clearly evident in the 2007 BTV-17 Wyoming outbreak where flocks pastured in the mountains, where lower temperatures naturally limit vector populations, had the most severe clinical disease and highest mortality (35%) when brought to lower elevations near the end of the vector season (Miller et al. 2010). With multiple circulating serotypes, general sheep populations cannot be considered as naive to BTV as the Northern European flocks were at the beginning of 2006. However, they would certainly be classified as naive seronegative populations for BTV-8 specifically. With little to no cross-neutralizing protection between serotypes (Schwartz-Cornil et al. 2008), they would not be protected. Results of this study suggest that should BTV-8 emerge in the United States, this common sheep breed would be highly susceptible to infection, as expected, but clinical disease would likely be similar in form, duration, and severity to what is typically observed in moderate to severe outbreaks with our endemic serotypes, not the unprecedented devastation seen in Northern Europe. In addition, a majority of exposed sheep would be expected to survive, acting as significant BTV-8 amplifying reservoirs with high titer viremias for subsequent transmission by Culicoides spp. to other livestock and to wildlife populations, facilitating establishment.
Footnotes
Acknowledgments
Funding for this research was provided by the USDA, ARS, NP103 Animal Health National Program, project number 5430-32000-006. Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the USDA.
Author Disclosure Statement
No competing financial interests exist.