
Abstract
Wildlife remains an important source of zoonotic diseases for the most vulnerable groups of humans, primarily those living in rural areas or coexisting with forest. The Upper Paraná Atlantic forest of Misiones, Argentina is facing ongoing environmental and anthropogenic changes, which affect the local biodiversity, including the bush dog (Speothos venaticus), a small canid considered Near Threatened globally and Endangered locally. This project aimed to expand the knowledge of zoonotic parasites present in the bush dog and the potential implications for human health and conservation medicine. From May to August 2011, a detection dog located 34 scats that were genetically confirmed as bush dog and georeferenced to northern Misiones. Of these 34 scats, 27 had sufficient quantity that allowed processing for zoonotic parasites using morphological (sedimentation and flotation) and antigen (coproantigen technique) analyses. Within these 27 scats, we determined that the parasitic prevalence was 63.0% (n = 17) with 8 (47.1%) having mixed infections with 2–4 parasitic genera. No significant differences (p > 0.05) between sampling areas, sex, and parasite taxa were found. We were able to summarize the predominant nematodes (Ancylostoma caninum, Toxocara canis, and Lagochilascaris spp.), cestodes (Taenia spp. and Spirometra spp.), and apicomplexa (Cystoisospora caninum) found in these bush dogs. With the copro-ELISA technique, 14.8% (n = 4) of the samples were positive for Echinococcus spp. This study represents the first comprehensive study about parasitic fauna with zoonotic potential in the free-ranging bush dog. This information combined with the innovative set of techniques used to collect the samples constitute a valuable contribution that can be used in control programs, surveillance of zoonotic diseases, and wildlife conservation, both regionally and across the bush dog's broad distribution.
Introduction
W
Despite the bush dog's (Speothos venaticus) broad distribution, which ranges from Panama across northern South America south to Paraguay and northeastern Argentina (DeMatteo et al. 2011), it is not considered abundant in any location (DeMatteo 2008, DeMatteo and Loiselle 2008, Oliveira 2009). The lack of detailed information on its status results in the species listed as Near Threatened globally (DeMatteo et al. 2011). The seminomadic bush dog, which is noted to occur in groups and as individuals (DeMatteo et al. 2011), is crepuscular using dens/burrows for shelter (Lima et al. 2012), favoring those previously used by nine-banded armadillos (Dasypus novemcinctus) and paca (Cuniculus paca).
Bush dogs are considered highly carnivorous consuming a variety of small and medium prey [e.g., paca, nine-banded armadillo, agouti (Dasyprocta azarae), capybara (Hydrochoerus hydrochaeri), ground birds] plus large prey [e.g., tapir (Tayassu tajacu)] and fruits (Deutsch 1983, Peres 1991, Strahl et al. 1992, Aquino and Puertas 1997, Silveira et al. 1998, Wallace et al. 2002, Beisiegel and Zuercher 2005, Zuercher et al. 2005, DeMatteo 2008, DeMatteo et al. 2011). Considered a habitat generalist, the bush dog has been associated with a variety of habitat types, including fragmented and altered (Michalski and Peres 2005, DeMatteo and Loiselle 2008, Lima et al. 2012, Michalski 2010); however, these human-modified areas are associated with an increased home range size (Lima et al. 2014) and increased risk of disease exposure through contact with domestic dogs (DeMatteo et al. 2011).
Postmortems of captive/semicaptive bush dogs and opportunistic samples from free-ranging animals have provided insight into the disease-causing agents present in this species, including bacteria (Escherichia coli, Klebsiella spp., Proteus vulgaris, Shigella spp., Staphylococcus aereus, S. epidermis, Bartonella spp.), viruses (canine parvovirus, rabies), fungi (Candida spp., Mycoplasma spp.), mites (Sarcoptes scabiei), and parasites (Echinococcus vogeli, Lagochilascaris spp. Spirocerca lupi, Leishmania infantum, Giardia spp.) (Rausch and Berstein 1972, Volcán and Medrano 1991, Oostburg et al. 2000, Beisiegel and Zuercher 2005, Jorge et al. 2007a, b, Chebez 2008, Jorge et al. 2008, Felix Lima et al. 2009, Rinas et al. 2009, Fleischman et al. 2015).
Several of these infectious agents are zoonotic, including E. vogeli, which causes Neotropical polycystic echinococcosis in humans. As a definitive natural host of E. vogeli and predator of paca, the adult stage of the parasite stage lives in the intestines of the bush dogs, where parasite eggs are eliminated in feces resulting in ground contamination in the forest. The paca, the major intermediate host of E. vogeli, becomes a host when it feeds on wild fruits contaminated with viable parasite eggs. When the bush dog feeds on infected paca, the cycle is completed.
Humans are considered an accidental intermediate host. Domestic dogs become parasitized when humans feed the organs (e.g., liver) from paca. Domestic dogs can then transmit infection to humans indirectly through scat containing parasite eggs contaminating the ground around homes or directly through contact with viable eggs that remain attached to their fur (D'Alessandro et al. 1981, Otranto et al. 2015b). However, this indirect infection of humans can also result through exposure to E. vogeli eggs from free-ranging carnivores. Rural and indigenous populations are tightly associated with the forest and often make excursions to hunt meat and gather fruits; specifically, exposure to ground contaminated with infected scat of bush dogs or the consumption of unwashed, contaminated wild fruits.
Both Neotropical echinococcosis and human lagochilascariasis are emerging zoonoses with rural persons and persons coexisting with forest are considered the groups most vulnerable (Volcán and Medrano 1991, Barrera-Pérez et al. 2012, Vizcaychipi et al. 2013). Ongoing anthropogenic changes to the environment, like those in Misiones, Argentina, can favor the emergence/reemergence of diseases and new patterns of transmission in the synanthropic field (Daszak et al. 2000) making it essential that we understand the current status of zoonotic diseases so proper monitoring and management programs can be established.
While Misiones contains the largest remaining tract of Upper Paraná Atlantic forest, only ∼40% is located in protected areas (DeMatteo et al. 2014a, b) and the forest outside of protected areas is threatened with ongoing land conversion (Izquierdo et al. 2008). The current work aims to update knowledge of zoonotic parasites in free-ranging bush dogs in Misiones, with a special focus on its relation to human health and conservation medicine. We used a combination of direct morphological measurements and coproantigen detection to evaluate parasite loads and suggest possible implications from our findings.
Materials and Methods
Study area
Misiones, Argentina, which is bordered by Paraguay and Brazil, is located between 25°28′S–28°10′S and 53°38′W–56°03′W. The province is characterized by humid, subtropical climate with no distinct dry season (Crespo 1982). The average monthly rainfall is about 100 mm; however, in October and November the rainfall can be >200 mm. The hot season (late September to mid April) is characterized by warm days (30–35°C) and moderate nights (18–24°C) compared with the cool season that has moderate days (23–27°C) and cool nights (9–12°C).
Sample collection
Scats were collected from May to August 2011 (late fall to early spring) with the assistance of a detection dog that was trained to indicate on five target species: jaguar (Panthera onca), puma (Puma concolor), ocelot (Leopardus pardalis), oncilla (Leopardus tigrinus), and bush dog (DeMatteo et al. 2014a, b).
Precautions were taken with the detection dog to prevent environmental contamination with common domestic dog diseases, as well as transfer of wildlife diseases to other regions of the world (e.g., Argentina to USA). Before transport to Argentina, the detection dog received an international USDA health certicificate, typical vaccines (6 in 1 distemper and rabies), and medicines for internal/external parasites [i.e., ivermectin (tablet and paste), pyrantel, imidaclorprid, and deltamethrin). In Argentina, the detection dog continued to receive treatments for internal/external parasites at an interval recommended by a veterinarian. Before returning to the USA, a screening of parasites was conducted, per SENASA requirements. An additional health screening plus continued preventive treatment for internal/external parasites was done upon returning to the USA.
With each scat, its GPS location, composition, approximate time in the environment, and location relative to trails/roads were recorded (DeMatteo et al. 2014a, b). After locating a sample, the surface of the scat was swabbed with a cotton-tipped applicator soaked in phosphate buffered saline solution to collect cells sloughed from the animal's intestine (Ball et al. 2007, Rutledge et al. 2009). These swabs are used to genetically identify species, individual, and gender of each scat (DeMatteo et al. 2014a, b). In addition, each sample was collected and stored in a labeled, 18oz Whirl-Pak®bag for later use in this and other studies. At the end of the day, all swabs and scats were preserved at −20°C.
Since samples were from free-ranging animals, where type and load of pathogens were unknown, work was conducted in Biosafety Laboratory Level III (BSL3). A 5–10 g aliquot of each preserved scat was separated and stored in a labeled resealable plastic bag. Samples were inactivated for two weeks at −70°C before subdividing into two 15-ml tubes for subsequent analyses. For morphological analyses, the tube was filled with saline–formalin solution and agitated for 24–48 h to hydrate the sample. For antigen analysis, the sample was mixed with equal parts PBS-0.3% Tween centrifuged for 30 min at 3500 rpm then the resulting supernatant was removed and stored at −70°C until processing. All analyzed samples were coded for long-term storage at −70°C in Argentina's National Reference Center at the Instituto de Infectologia (INEI-ANLIS “Dr. Carlos G. Malbrán) in Bueños Aires.
Morphological analyses
Scat samples were processed using sedimentation (modified Teleman) and flotation (Sheather and Willis) techniques (WHO 1991). Morphometric identification and taxonomic assignment were completed using a Leica ICC50 HD microscope and LAS EZ software (Leica Application Suite). This combination captured digital images that allowed fine-scale measurements (microns) of parasitic structure.
For genus and species identification of protozoa and helminth eggs, previously described morphological (i.e., shape, cover, color, content) and metric (i.e., length, diameter) criteria were used (Anderson et al. 1974, Acha and Szyfres 1986, Soulsby 1987, Volcán and Medrano 1991, Anderson 2000, Bowman et al. 2004, Beltran-Saavedra et al. 2009, Scioscia et al. 2014). The mean ± standard deviation (SD) was determined when there were ≥3 eggs of a genus. Prevalence was determined by evaluating the proportion of samples with infection compared with the total number of samples tested. A chi-squared test was used to evaluate whether there were significant differences between sampling areas, sex, and parasite taxa.
Antigen analysis
Enzyme immunoassays (ELISA) are a semiquantitative technique to detect coproantigens, whose presence in scat can be used to diagnose various intestinal diseases (e.g., helminthes and protozoans) and infections due to bacteria and viruses. A genus-specific copro-ELISA, with the technique previously described by Guarnera et al. (2000), was used to detect the presence of Echinococcus spp. antigens in the current samples. Because this copro-ELISA test has a very high specificity (98%) and high sensivity (70%), crossreactivity with other genera (e.g., Taenia spp.) is unlikely. The next step will be to confirm species identification using copro-PCR (e.g., Abbasi et al. 2003, Hüttner et al. 2008) and optimization of extraction protocols is ongoing.
Results
Sample collection
Of the collected scat, 34 were confirmed as bush dog (DeMatteo et al. 2014b), from which 27 had sufficient quantity of scat for the current study and 22 of this subset could be identified to the individual and gender level (Table 1). Of the 20 unique individuals represented in these 22 scats, two individuals (one male and one female) were found two times each with a mean distance (± SD) between scat of 1.50 ± 3 km. Since data collection focused on maximizing area covered in the survey and species detection, the density of bush dogs could not be determined.
Of the 27 scat, approximately half (n = 12) were collected in six protected areas in northern–central Misiones (Table 1 and Fig. 1). The remainder (n = 15) were located outside of protected areas: around Parque Provincial (P.P.) Guardaparque H. Foerster, W/SW of P.P. Urugua-í, along Ruta 101, and east of P.P. Cruce Caballero/Valle del Arroyo Alegría (Table 1 and Fig. 1). Specific locations for the current scats are not provided due to local legal regulations aimed at protecting native wildlife.
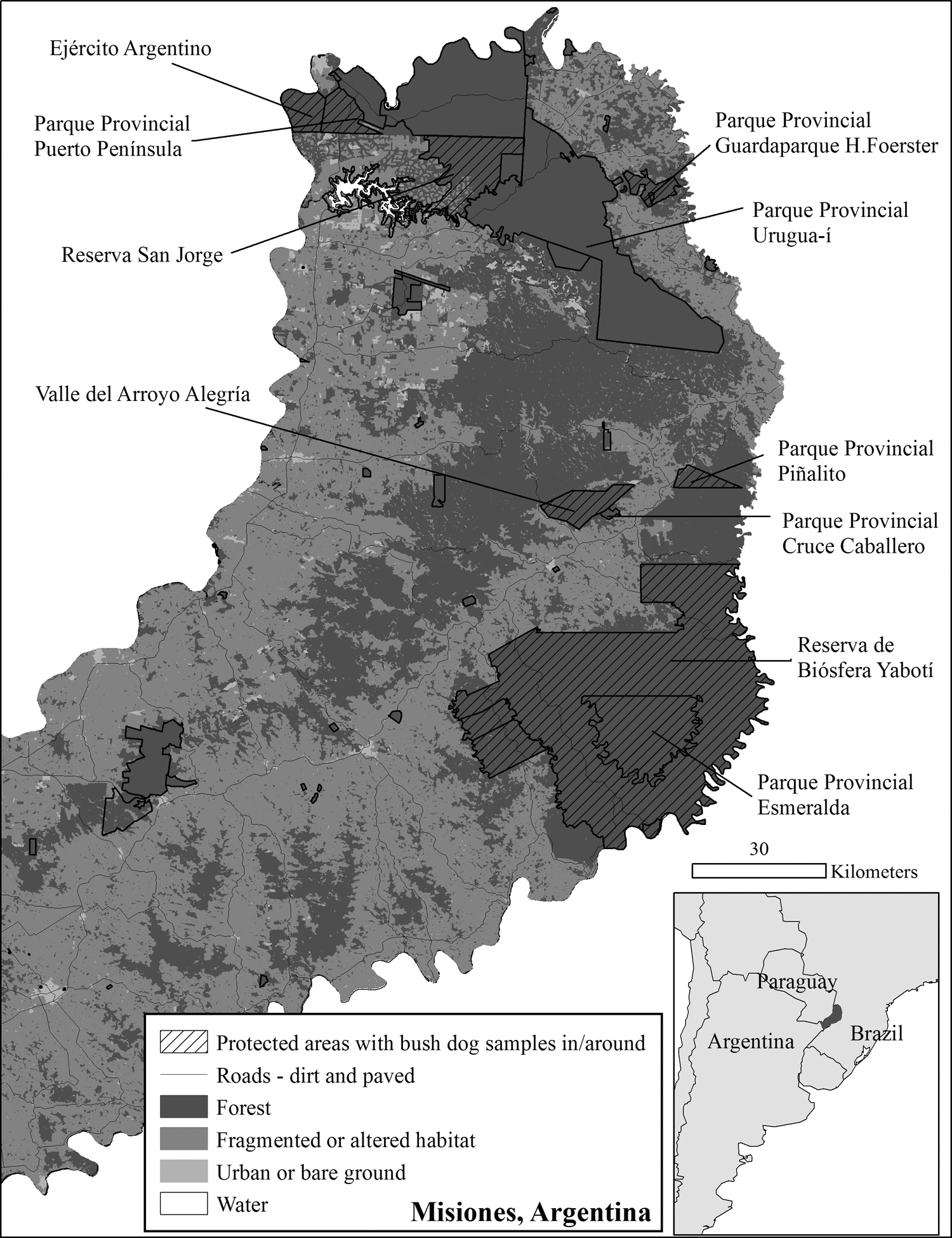
Location of Misiones, Argentina in South America (inset). Map of Misiones showing protected areas, where bush dog samples were located in or around. These areas are shown in relation to the roads and land-use pattern existing in Misiones in 2009: forest, fragmented or altered areas, urban or bare ground, and water [Izquierdo et al. (2008) land cover map updated by A. Izquierdo for 2009 personal communication].
Morphological analyses
In the parasitological analyses of these 27 samples, we found a 63% prevalence (n = 17) of parasitic taxon in 16 of the individual bush dogs, with a single individual represented twice. Of these 17 samples with parasitic taxon, 8 (47.1%) had mixed infections with 2–4 parasite genera. Included in these 8 samples was the single individual represented twice with the infection being identical in both samples.
The frequency of specific taxonomic infections in these samples can be broken down into 17 (63%) positive for genera in the class Secernentea and phylum Nemata, 6 (22.2%) positive for genera in the class Cestode and phylum Platyhelminthes, and 4 (14.8%) positive for genera in the class Apicomplexa and phylum Protozoa (Table 2). Among the genera in the phylum Nemata, Ancylostoma caninum and Toxocara canis had higher occurrences in the northern versus central zones (Table 2). Lagochilascaris spp. (class Secernentea), Taenia spp. (class Cestode), and Cystoisospora caninum (class Apicomplexa) had similar proportions in the northern and central zones (Table 2). Spirometra spp. in the cestode class was only in the northern zone (Table 2). No significant differences were found between sampling areas, sex, and parasite taxa [χ2 (1, N = 19) = 0.53, p = 0.46]. The mean length and diameter of all eggs and oocysts were similar to previously published results (Table 3).
In addition, a summary of how the parasitic taxon are split among the northern and central (N-C) zones.
When no SD or range is provided, all values were equal.
Antigen analysis
Among the 27 samples analyzed with the copro-ELISA technique, 14.8% (n = 4) were positive for the Echinococcus spp. antigen.
Discussion
We identified five zoonotic helminthes (T. canis, Lagochilascaris spp., Spirometra spp, Echinococcus spp., and A. caninum), which represent the first time that parasites have been systematically documented in free-ranging bush dogs. All of these infections are among the Neglected Tropical Diseases (NTDs) that negatively affect Global Public Health and represent a main cause of human morbidity and mortality in developing countries (WHO 2007). Each genus has a common life cycle involving carnivores as the definitive host, as their intestines allow the adult parasites to reach maximum physiological and reproductive levels (Beaver 1958, Volcán et al. 1993). This fact has a profound epidemiological significance on human health and ecology, since it corresponds to the period of environmental contamination and risk of parasitic transmission among wildlife, domestic animals, and humans (Umhang et al. 2014, Otranto et al. 2015b).
In the Upper Paraná Atlantic forest, the soil conditions, elevated precipitation, and temperature range offers suitable ecological conditions for dispersal, evolution, and survival of infective stages of helminth eggs and larvae, like those found in the present study. From an ecological perspective, this shows the diversity of specificity used by parasites, as well as the breadth of their niche resources, with values of prevalence, intensity, and abundance indicating ecological differences between hosts and habitat (Rohde 1994, Mehlhorn and Piekarski 1998, Tompkins and Clayton 1999). Parasitological studies contribute to the elucidation of the host's ecology, including diet, microhabitat characteristics, and species interactions (Brooks and Hoberg 2000, Poulin and Morand 2004, Navone et al. 2012), as well as emphasize the importance of zoonotic studies in conservation medicine.
The three techniques used in this study constitute a single tool to evaluate parasites in their natural environment through the collection of species-specific samples with detection dogs, determination of genetic identities for samples, and spatial analysis of samples with GIS. While the use of scat in parasite studies is not new (Elmore et al. 2013, Massara et al. 2015, van Kesteren et al. 2015), these studies have traditionally located samples opportunistically or through captured animals. The use of detection dogs to locate scats is a way to maximize collection success, minimize collection of nontargets, maximize area surveyed, and remove bias in finding samples (DeMatteo et al. 2009, 2014a, b).
While this combined approach has had limited application to date (Vynne and Kinsella 2009), the current results demonstrate how this set of techniques provides an integrated approach that can be used to inform control programs, monitor zoonoses, predict animal movement, and identify potential areas of risk for coexisting human–animal populations. Detection dogs and microsatellites provided samples from 20 unique individuals, whereas GPS and GIS technology allowed an evaluation of the spatial distribution/abundance of parasites present among and between individual bush dogs.
Most likely due to the relatively low number of samples, no statistical differences were found in the distribution of parasites (northern or central zones) or parasitic assemblies; however, these techniques did allow for the evaluation of two individuals 2 × , which increases the probability of detecting parasitic elements since each sample potentially represents a different stage of the parasite's biological cycle (Beaver et al. 1985, Botero 1994).
The fact that T. canis eggs were found in 22.2% of the bush dog scat (3 males and 3 females) suggests that this technique is effective at detecting infections in juveniles (<1 year) or puppies since the avenue T. canis larvae take in the definitive host is age dependent (Botero 1994). In older dogs, the T. canis larvae reach the blood through the lung, but are subsequently deposited in tissue where they fester, so it is not possible to detect eggs in the scat of older animals. However, in younger animals, the T. canis larvae traverse the pulmonary alveoli and ascend to the pharynx, where they are swallowed resulting in adult parasites in the small intestines, which allow both eggs and adults to be released in scat.
It is possible that some of the prey in the bush dog's varied diet (e.g., paca, agouti, capybara, nine-banded armadillo) may behave as partenic versus intermediate hosts, in which parasite development does not occur, but the species can serve as an ecological bridge to link a food gap in the cycle of the parasite (Glickman and Schantz 1981, Bush et al. 2001, Cardillo 2012). These paratenic hosts would essentially help in the temporal and spatial spread of the infective stage of T. canis, overcoming the temporary absence of a definitive host.
Group living in the bush dog and many of their prey would also favor the transmission of T. canis by creating a high density of animals contaminated with larval eggs. Given the risk to public health, corroborating the epidemiological implications of these partenic hosts and their link to transmission of zoonoses would benefit the development of preventive measures for coexisting human–animal populations.
The confirmation of Lagochilascaris spp. corresponds to a previous report in bush dogs from Venezuela (Volcán and Medrano 1991). Present in Neotropical areas, this parasite lacks a defined lifecycle, but is known as responsible for lagochilascariosis in humans. Human infection takes place after ingestion of raw or undercooked meat from rodents (e.g., capybara, agouti), which act as intermediate hosts harboring encysted larvae of the parasite in their muscles (Costa Araújo et al. 1986, Volcán and Medrano 1991, de Moura et al. 2012). Like Volcán and Medrano (1991), we consider the confirmation of Lagochilascaris spp. eggs in male and female bush dogs in the northern–central zones of Misiones to be important with the hope that it can contribute to the elucidation of the parasites natural life cycle and the epidemiology of the disease.
This confirmation of Spirometra spp. represents the first report of this cestode in bush dogs. The overlap between the morphological measurements of the eggs in this study with Scioscia et al. (2014) suggests that the specific genus found in Misiones may be Spirometra erenicei. This study's confirmation of Spirometra spp. in the northern zone of Misiones highlights a potential zoonotic scenario with high availability of transmissible parasitic forms, carnivorous hosts to complete the life cycle, and resident human populations with cultural habits that enable transmission (De Roodt et al. 1993). Specifically with this cestode, adult parasites are present in the intestines of felids and canids and subsequently contaminate freshwater when their eggs are excreted with the host's scat.
Continuation of the cycle requires two intermediate hosts: aquatic crustaceans or copepods of the genus Cyclops and paratenic tetrapods (i.e., amphibians, reptiles, humans, or other mammals) (De Roodt et al. 1993, Scioscia et al. 2014). In humans, the two main infestation routes are penetration of plerocercoid larvae through skin or mucus membranes and through eating of raw food or water that is contaminated with plerocercoid larvae or crustaceans that contain plerocercoids. This extended life cycle and the plerocercoid larvae are the cause of sparganosis in humans, an emerging zoonotic disease; however, it is the adult cestode larva that causes most human infections with Spirometra mansoni and Spirometra erinacei most common worldwide and Spirometra mansonoides associated with infections in South America.
With the coproantigen technique, used in programs to control hydatidosis/echinococcosis (Eckert et al. 2001), we found 14.8% of the bush dog scats positive for Echinococcus spp. antigens, indicating this genera is circulating between definitive hosts in the Upper Paraná Atlantic forest. It is possible that this is E. vogeli, as this was recently isolated in the forests of Misiones.
All results align with the epidemiology of the disease (D'Alessandro and Raush 2008, D'Alessandro 2010, Vizcaychipi et al. 2013, Carmena and Cardona 2014, Mayor et al. 2015, Navarro et al. 2015), considered an emerging zoonotic disease in Neotropical southern cone and pan-Amazonian countries. Since the parasites found in the scat allow us to indirectly observe the animal's diet, this information can be used to explore the interaction of these parasites and their free-ranging hosts (final and intermediate) and determine possible phylogenetic relationships.
Almost a quarter (26.2%) of the landscape in Misiones is composed of rural populations and 1.1% of this contains Mbya Guaraní communities or villages scattered in the Green Corridor, private land, and ecological reserves (INDEC 2010; IPEC 2012). The close association of these communities with the ecosystem reinforces the need to raise awareness, strengthen, and broaden the search for these zoonoses in wildlife areas. In addition, it places health, ecological–environmental, and tourist officials on alert to take preventive measures, monitoring, and control. The novel approach used in this study highlights the scientific value of this work, provides a new set of techniques to address zoonotic questions, and provides an avenue for preserving wildlife in a framework of public policies.
Footnotes
Acknowledgments
Funding for the collection of scats and related genetic analyses was provided by Conservation, Food, & Health Foundation, a National Geographic Society Conservation Trust Grant, the Palm Beach Zoo Conservation Fund, the Zoological Gardens of Chester–North of England Zoological Society Conservation and Research Fund, and the Jaguar Conservation Fund–Woodland Park Zoo. The Ministerio de Ecología y Recursos Naturales Renovables of Misiones (MEyRNR) and the Administración de Parques Nacionales of Argentina provided all collection/handling permits, help with field logistics, and field housing. Funding for the processing of scats for this study was provided by Instituto Nacional de Enfermedades Infecciosas (INEI)–ANLIS “Dr. Carlos G. Malbran,” Buenos Aires, Argentina. Sincere thanks to the numerous Argentinean students, provincial and national park guards, and private land owners who assisted with various aspects of this project, both in and out of the field. The authors also thank Graciela Santillan, Graciela Cespedes, Ariel Naidich, and Ariana Gutierrez for their invaluable assistance with various laboratory analyses, the parasitic immunology service for logistical support, and the enteroparasite division for use of a microscope for morphological measurements. A special thanks to Antonio D'Alessandro whose time, knowledge, and patience has been instrumental in establishing this comprehensive approach to studying neotropical parasites and zoonoses.
Author Disclosure Statement
No competing financial interests exist.