
Abstract
The purpose of this review is to provide practical information to help researchers intending to perform “from field to laboratory” studies on phleboviruses transmitted by sandflies. This guideline addresses the different steps to be considered starting from the field collection of sandflies to the laboratory techniques aiming at the detection, isolation, and characterization of sandfly-borne phleboviruses. In this guideline article, we address the impact of various types of data for an optimal organization of the field work intending to collect wildlife sandflies for subsequent virology studies. Analysis of different data sets should result in the geographic positioning of the trapping stations. The overall planning, the equipment and tools needed, the manpower to be deployed, and the logistics to be anticipated and set up should be organized according to the objectives of the field study for optimal efficiency.
How to Determine the Region for Trapping Sandflies to Search for Viruses
Using entomological data
S
Female individuals of sandflies require a blood source for egg maturation and both female and male individuals need a sugar source for energy. Sandflies' weak flight capability is affected by the wind and windy weathers make conditions difficult for sandflies to achieve the sugar and blood sources (Alexander 2000). After maturation of the eggs, they are laid in the soil that is rich in organic matter such as herbivorous animal feces that provide food for larvae (Feliciangeli 2004). Therefore, it is important to place traps in or near animal housing places due to these requirements. Sandflies are mainly dispersed in rural and periurban areas; thus, collaborating with local veterinarians might help with finding suitable places for setting traps and explain to the local people the aim of the trapping.
Using parasitology data
Besides phleboviruses, sandflies can also transmit the flagellate protozoan Leishmania that cause three forms of the disease called leishmaniasis: (1) visceral leishmaniasis, which affects 300,000 people with more than 6.6% lethality rate, (2) cutaneous leishmaniasis, with more than 1 million cases worldwide, (3) and mucocutaneous leishmaniasis with most cases occurring in South America (WHO 2014). Leishmaniasis is listed in the 10 most worrying neglected tropical diseases (
Using virology data
Seroprevalence studies performed using the sandfly-borne phlebovirus antigens are of utmost interest to help researchers at the design step of field studies aiming at the detection, isolation, and characterization of viruses transmitted by phlebotomine flies (Fig. 1 and Supplementary Table S1; Supplementary Data are available online at
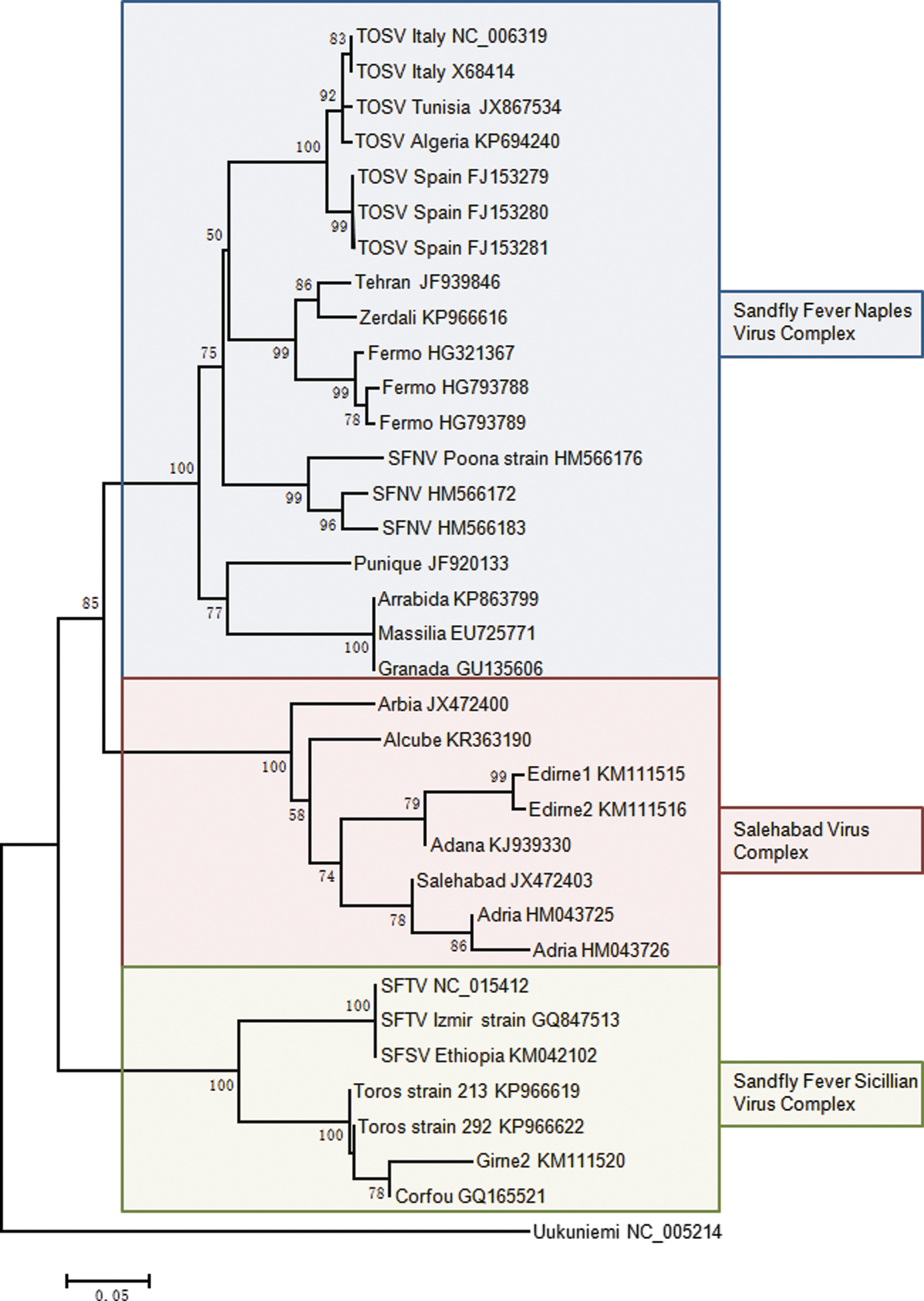
Phylogenetic analysis of Old World sandfly-borne phleboviruses using a 193-amino acid region in the polymerase protein. Sequences were aligned using the Clustal W program. Distances and groupings were determined by the p-distance method and neighbor-joining algorithm implemented with the pairwise deletion model in the MEGA 6.06 software program. Bootstrap values are indicated and correspond to 500 replications.
Using medical data
Phleboviral infections demonstrate a seasonal incidence peaking between April and October, depending on the geographical location (Tesh et al. 1976), correlated with the regional sandfly activity (Figs. 2 –5). Medical reports on outbreaks in autochthonous or imported populations as well as case reports are indicative of the presence of infected sandflies in specific geographic areas (Supplementary Table S1). The main problem of the clinical diagnosis is the symptoms being nonspecific; thus suspected cases must be confirmed by virological methods to demonstrate either the presence of the virus in blood or cerebrospinal fluid or the seroconversion in two successive serum samples. Since standardized and commercialized assays for the RT-PCR detection of these viruses are lacking and a limited number of commercially available serological tests are available, definitive confirmation is rarely obtained and the majority of probable cases remain unconfirmed.
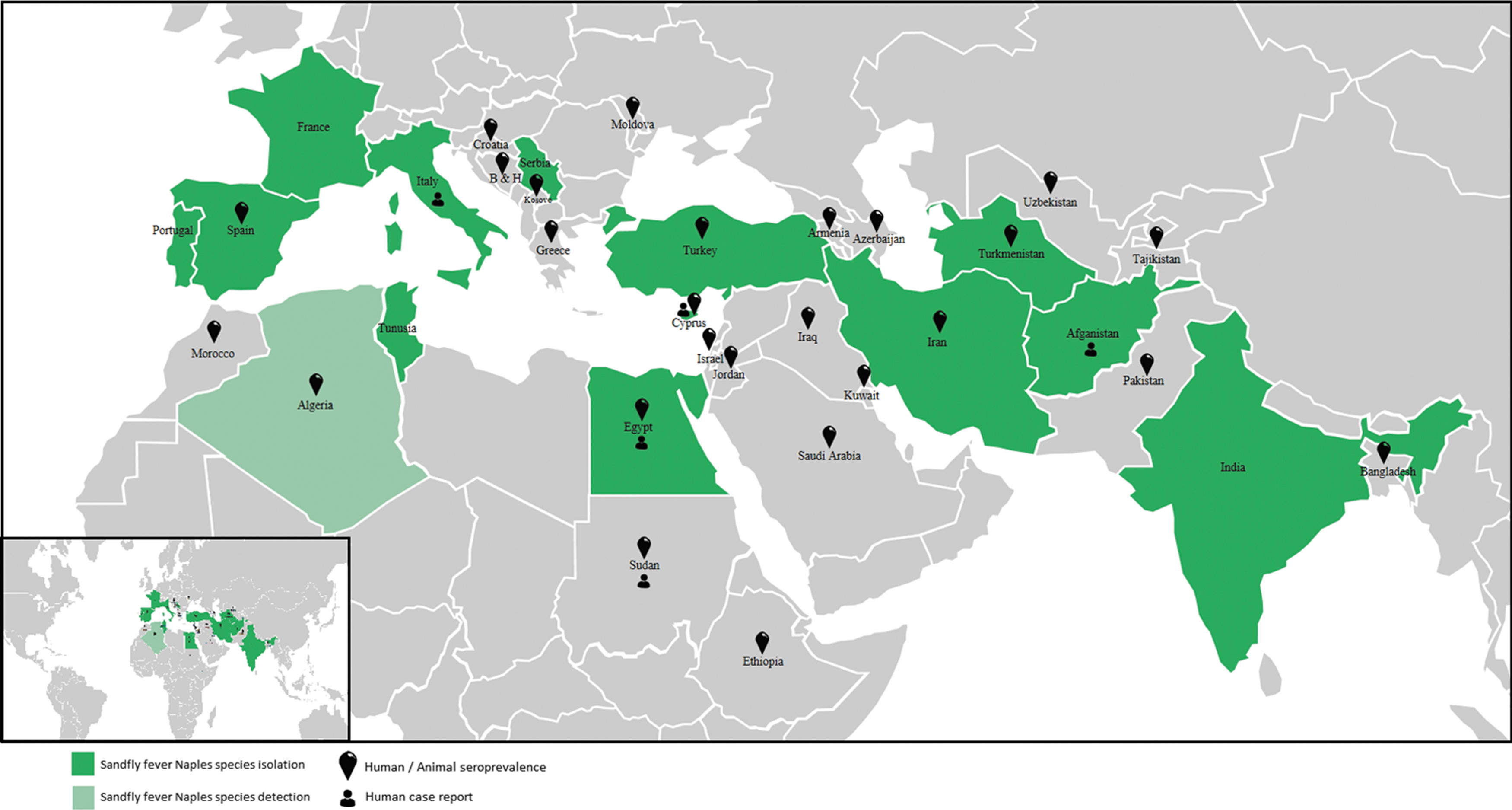
Countries where data are available for seroprevalence, PCR detection, and virus isolation for viruses belonging to the Sandfly fever Naples species.
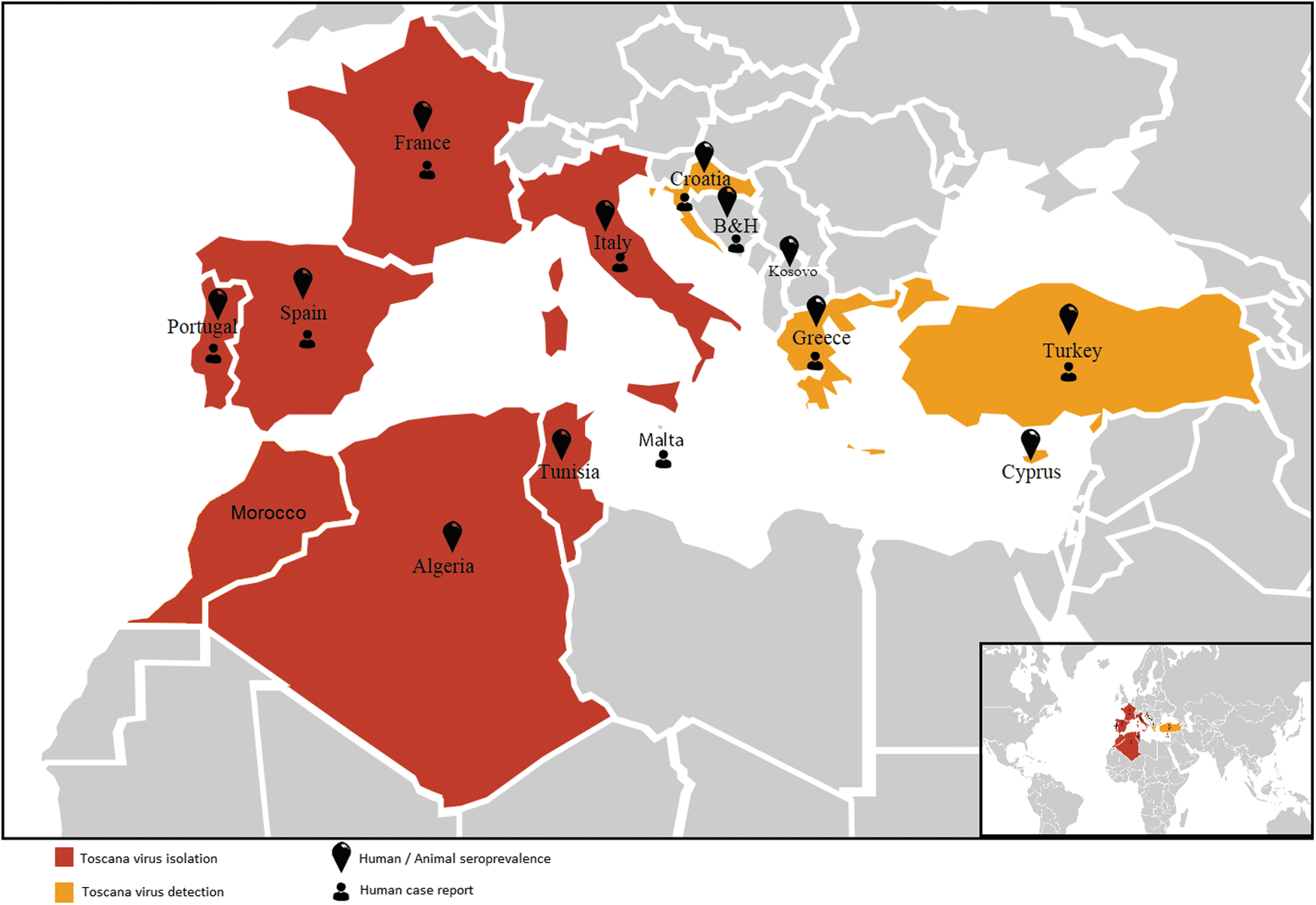
Countries where data are available for seroprevalence, PCR detection, and virus isolation for Toscana virus.
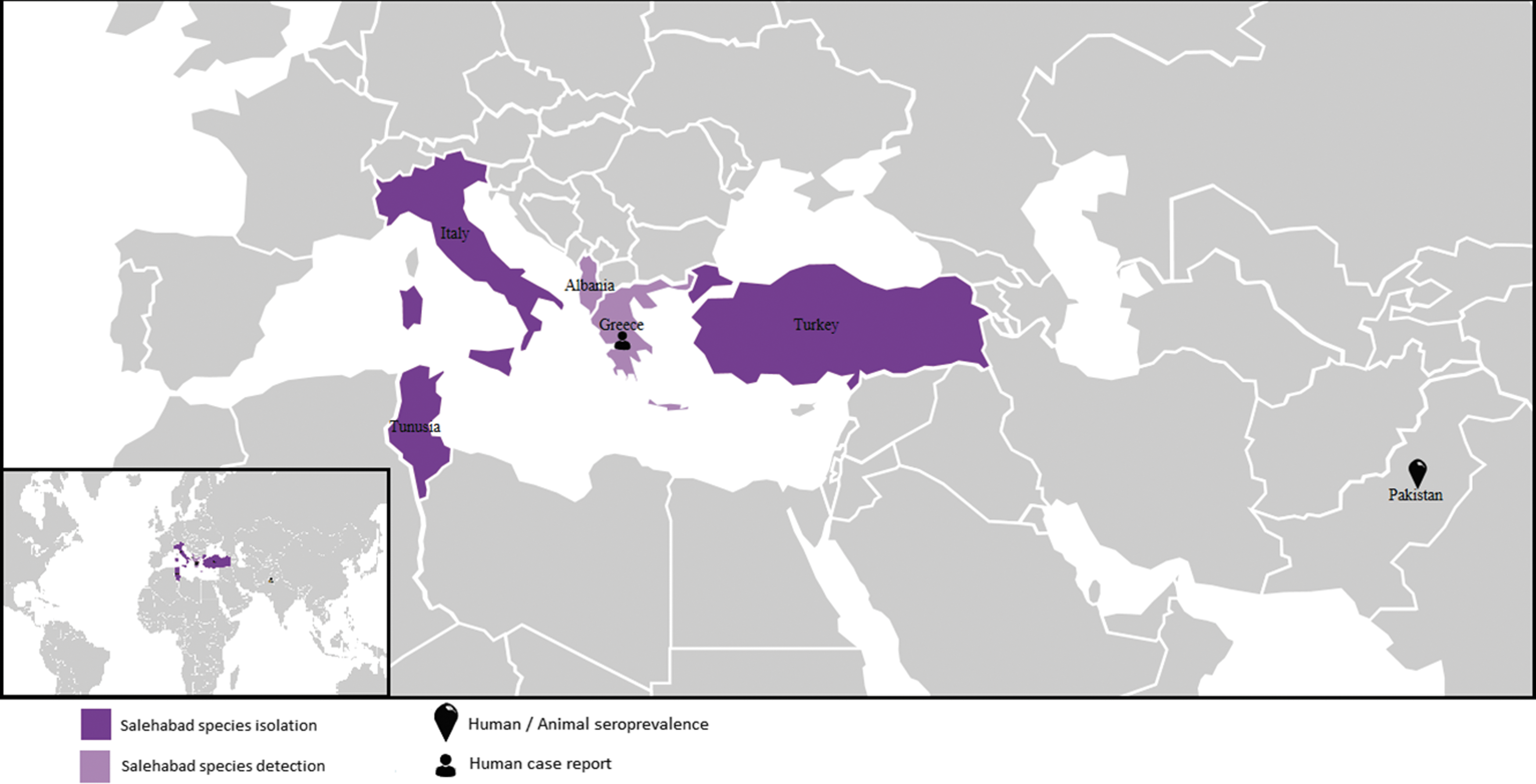
Countries where data are available for seroprevalence, PCR detection, and virus isolation for viruses belonging to the Salehabad species.

Countries where data are available for seroprevalence, PCR detection, and virus isolation for viruses belonging to the sandfly fever Sicilian serocomplex.
Naples and Sicilian viruses have identical clinical syndromes, which are fever, headache, malaise, photophobia, myalgia, and retro-orbital pain. Because the fever lasts for 2–3 days, the disease was named as “3-day fever.” In contrast, TOSV can cause aseptic meningitis, or meningoencephalitis presenting with headache, fever, nausea, and vomiting in infected individuals (Dionisio et al. 2003, Charrel et al. 2005, 2012, Depaquit et al. 2010). During World War II, a large number of soldiers was affected by sandfly fever (Sabin 1951). Recently, TOSV human case records came from Italy (Serata et al. 2011, Calzolari et al. 2014), France (Dupouey et al. 2014, Marlinge et al. 2014), Portugal (Santos et al. 2007, Amaro et al. 2011), Croatia (Punda-Polić et al. 2012), Turkey (Ocal et al. 2014, Ergunay et al. 2015), Greece (Papa et al. 2014), and Tunisia (Fezaa et al. 2014) (Fig. 3). A large sandfly fever Sicilian virus outbreak recently occurred in Ethiopia (Woyessa et al. 2014). However, due to lack of specific manifestations and reliable differential clinical diagnosis, medical records need to be complemented by virological and microbiological tests for the definitive etiological identification.
Using veterinary data
Although the capacity of sandfly-borne phleboviruses to cause diseases in animals is currently unknown, accumulating data indicate that mammals can be infected with at least some of these viruses (Navarro-Marí et al. 2011, Alkan et al. 2013, 2015b, Sakhria et al. 2014, Dincer et al. 2015, Bichaud et al. 2016, Tahir et al. 2016); accordingly they can serve as sentinels for the presence of the corresponding viruses. There is no undisputable evidence that birds can be infected by sandfly-borne phleboviruses, but few studies have addressed this point.
Using ecological and environmental data
Since the dynamics of sandfly populations is intimately linked to environmental parameters, ecological data are of great importance for an optimal yield of field studies. The organization of field collections requires a deep survey analysis in the study region. The suitable habitats for Phlebotominae sandflies need to be determined using climatic and geographic data. Sandflies are small (1.5–3 mm), delicate, nocturnal insects with short distance flight capability. Factors such as yearly, monthly, and daily temperatures can have a major impact on sandfly population size and activity, and therefore can affect the sampling success (Tesh et al. 1976, Alexander 2000). The altitudinal distribution and climatic needs are varying between sandfly species from sea level to 3500 m (Killick-Kendrick 1999, Aransay et al. 2004, Guernaoui et al. 2006a, 2006b, Belen and Alten 2011, Alten et al. 2015). In Spain, Phlebotomus ariasi was collected at higher altitudes (600–900 m) from coolest and most humid Mediterranean bioclimatic zone (supra-Mediterranean), whereas Phlebotomus perniciosus predominated in the lower altitudes, warmer and drier bioclimatic zones (Aransay et al. 2004). Biogeographic parameters have a huge impact on the species distribution and density (Zhioua et al. 2010, Fares et al. 2015). Rainfall is another factor with a huge impact on sandfly activity; heavy rains could decrease the flight range of the sandflies. In Panama, rainfall amount and distribution were found to correlate with seasonal sandfly density (Chaniotis 1974). The adult individuals resting sites are animal barns, houses, poultries, caves, tree holes, animal burrows, spaces between rocks, and holes of walls. Heavy rains could flood these resting sites and reduce suitable places for sandflies (Alexander 2000). Old traditional animal husbandry barns with stone construction can shelter bigger sandfly populations than modern new farms, due to providing more resting sites. However, sandfly species differ in their preference for resting sites. For instance, although Sergentomyia minuta tend to rest between small rocks, Phlebotomus mascitii has special habitat preference, which mainly includes caves (Grimm et al. 1993, Alten et al. 2015).
In addition, insecticides have huge effects on sandflies. In Greece, for instance, due to high-level DDT spraying in nation-wide malaria control program, the number of sandflies dramatically decreased in the year 1946 (Hadjinicolaou 1958, Tesh and Papaevangelou 1977). It would be useful to ask the local people in the trapping region if they use insecticides.
How to Organize for Field Collection
The objectives of the study determine the global organization of the field collection, the equipment and tools needed, the manpower to be deployed, the logistics to be anticipated, and the setup. Depending on the aim of the study, the field area can be chosen for specific sandfly species. Until now, Sicilian virus was isolated from Phlebotomus papatasi in 1943 by Albert Sabin (Sabin 1951) and following studies show the presence of Sicilian-like viruses in P. ariasi in Algeria (Izri et al. 2008, Moureau et al. 2010) and in Phlebotomus longicuspis, P. perniciosus, and S. minuta in Tunisia (Zhioua et al. 2010). Sandfly fever Turkey virus, a variant of the sandfly fever Cyprus virus, which are considered as Sicilian-like phleboviruses, was detected in Phlebotomus major complex (Ergunay et al. 2012). Naples virus was isolated from P. perniciosus in Italy (Vesenjak-Hirjan et al. 1980) and from Phlebotomus perfiliewi in Serbia (Gligic et al. 1982). The first isolation of TOSV was in central Italy in 1971 from P. perniciosus and P. perfiliewi (Vesenjak-Hirjan et al. 1980). Consecutive studies show the presence of TOSV in S. minuta (Charrel et al. 2006). Massilia and Granada viruses were isolated from P. perniciosus (Charrel et al. 2009, Collao et al. 2010) and Punique virus was isolated from P. longicuspis and P. perniciosus (Zhioua et al. 2010). NW Phlebovirus species such as Buenaventura virus, Punta Toro virus, and Leticia virus were isolated from Lutzomyia sp. sandflies (Tesh et al. 1974). Despite extensive studies have been done around the Mediterranean area, vector–virus association remains poorly understood. Trapping in the regions that are known as endemic for the target virus could enhance the chances of success and increase the detection rate.
A clear definition of the objectives is of great importance to organize the field campaign in a manner that is suited to fulfilling these objectives. Different strategies depending on the purposes to be served are detailed in the sister review article entitled “Practical guidelines for studies on sandfly-borne phleboviruses: part II: important points to consider for field work and subsequent virological screening.”
Detection of new viruses is very likely in regions where sandflies are present at high density. In our experience, the larger the number of sandflies the higher the chance to find a new virus. Recent studies have demonstrated that several sandfly-borne phleboviruses that may belong to distinct genetic complexes frequently cocirculate in a given locality (Amaro et al. 2015, Fares et al. 2015, Charrel unpublished data). Cocirculation of several viruses has been showed to be more frequent than initially considered. The outcome of the field campaigns is related to the number of sandflies trapped and tested. Even though there were previous studies in the same region, detection or isolation of novel phleboviruses can still be achieved. Recently, new phleboviruses isolation/detection was achieved from Turkey (Alkan et al. 2015b, Ergunay et al. 2014), Portugal (Amaro et al. 2015), Italy (Remoli et al. 2014), France (Charrel et al. 2009, Peyrefitte et al. 2013), Albania (Papa et al. 2011), and Tunisia (Zhioua et al. 2010), which shows the huge diversity of phleboviruses transmitted by sandflies. Moreover, the differences in the number of naturally infected sandflies depend on the region. The prevalence of the phlebovirus RNA in sandflies (phlebovirus positive pool/total number of tested sandflies) are reported as 1/460 (Charrel et al. 2007, France), 7/798 (Charrel et al. 2009, France), 5/427 (Peyrefitte et al. 2013, France), 4/896 (Amaro et al. 2015, Portugal), 5/1910 (Ergunay et al. 2014, Turkey), 7/900 (Remoli et al. 2014, Italy), 10/1489 (Zhioua et al. 2010, Tunisia) in various efforts. It is assumed that these values more or less reflect the level of virus circulation in a region. Surely, the high number of collection would increase the chance to detect or isolate the virus.
Actually, the majority of studies aiming at virus discovery in field-collected sandflies has resulted in the identification of new viruses when using open-detection techniques (generic PCR assays and cell culture), in contrast with specific techniques (Charrel et al. 2009, Moureau et al. 2010, Zhioua et al. 2010, Alkan et al. 2015b, Bichaud et al. 2016). Such nonspecific techniques have also shown to be capable of isolation and characterization of viruses belonging to the Flavivirus genus, not only the Phlebovirus genus (Alkan et al. 2015c).
Conclusions
It is unfortunate to address the virus discovery efforts in nature, just as additions to the virology stamp album. It must be recalled that the evidence for TOSV pathogenicity in humans (which is currently the most widespread arthropod-borne virus in Europe with at least 250 million people living in at risk area) was assessed 12 years after the virus was discovered in the field. Besides, the Rockefeller foundation has supported the most eminent arbovirologists to conduct studies of these viruses for more than 30 years. Although there is no doubt that Next Generation Sequencing will reveal many new discoveries about these viruses, the need to isolate and characterize the strains initially identified at their natural habitat, as well as investigating their pathogenic impact, has recognized globally among virologists. Without well-characterized infectious virus strains, serosurveillance or serodiagnosis studies to identify the specific etiological agent responsible for outbreaks or epidemics in susceptible populations cannot be performed. When carried out properly, the neutralization assay is the recognized gold standard for all virological seroepidemiological investigations. The virological “stamp album” is and has been for more than 60 years the essential tool with which to conduct these investigations and thence to inform health agencies charged with the responsibility of enabling implementation of the necessary disease control strategies.
Footnotes
Acknowledgments
This work was done under the frame of EurNegVec COST Action TD1303. This work was supported, in part, by (1) the European Virus Archive goes Global (EVAg) project that has received funding from the European Union's Horizon 2020 research and innovation program under grant agreement no 653316 and by (2) the EDENext FP7- n°261504 EU project; this article is cataloged by the EDENext Steering Committee as EDENext453 (
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.