
Abstract
Shiga toxin-producing Escherichia coli (STEC) O157:H7 was isolated from 30 (4%) of 744 cattle hide swab samples collected at Estonian slaughterhouses within a 3-year monitoring program of zoonotic pathogens. The isolates were characterized by determining the presence of STEC main virulence factors, the antimicrobial resistance profiles, and the genetic relatedness by pulsed-field gel electrophoresis (PFGE). Thirteen strains carried the stx2 gene alone and 17 both the stx1 and stx2 genes. The most frequently detected stx subtype was stx2c, occurring alone (n = 12) or in combination with subtype stx1a (n = 13). All isolates harbored the intimin-coding eae gene and produced enterohemolysin. Twelve isolates (40%) showed resistance to at least one of the 14 antimicrobials and the isolates were predominantly resistant to streptomycin, sulfamethoxazole, and ampicillin. No extended-spectrum beta-lactamase-producing isolates were detected. PFGE characterization of the isolates showed an overall similarity higher than 75%, and four clusters based on 100% similarity were revealed.
Introduction
E
A considerable number of STEC serotypes have been recognized as causative agents of human disease, but severe infections and outbreaks have been mostly associated with the O157 serogroup (Scallan et al. 2011, EFSA Panel on Biological Hazards 2013). E. coli O157:H7 infections are often associated with consumption of raw or undercooked beef because ruminants, particularly cattle, have been deemed as the main reservoir of this pathogen (Ferens and Hovde 2011). Cattle and other ruminants can be linked also with STEC transmission through direct contact with animals and farm environment or consumption of water and fresh produce contaminated with cattle feces (Caprioli et al. 2005).
The incidence rate of STEC infection in humans in Estonia has ranged from 0.2 to 0.6 per 100,000 inhabitants in the period of 2010–2014 (Terviseamet 2015). In most cases, the serogroup of the infecting STEC strains was not identified.
At the slaughterhouse level, cattle hides have been considered major vehicles of carcass contamination by E. coli O157 and other STEC serogroups (Keen and Elder 2002, O'Brien et al. 2005). Microbial sampling of hides can give an indication of the STEC load at the slaughterhouse level. The results reflect the possibility of further contamination of carcasses and meat and represent a tool to evaluate the efficacy of good hygiene practices as a control measure of carcass contamination (EFSA 2009).
The aim of the present study was to evaluate the prevalence of STEC O157:H7 in cattle at slaughter in Estonia by investigating the contamination of cattle hides, which is an important source of these zoonotic pathogens in the first steps of beef processing. STEC O157:H7 isolates were analyzed for the presence of virulence genes, the subtypes of the Stx-coding genes, the antimicrobial susceptibility spectrum, and the pulsed-field gel electrophoresis (PFGE) patterns.
Materials and Methods
Sample collection
A total of 744 samples were collected and analyzed between January 2011 and December 2013 in the frame of national monitoring program. Hide swabs were taken from the 400-cm2 brisket area of the animals after exsanguination and before dehiding at slaughterhouse using a sterile abrasive sponge kit (3M™ hydrated sponges). The sponges were premoistened with 10 mL buffered peptone water. Immediately after sampling, the sponges were cooled to 5 ± 3°C and sent to the National Reference Laboratory.
The sample size was based on estimates of a predicted prevalence of 12%, with confidence intervals (CIs) of 95% and an accuracy of 4% in the total annual number of slaughtered cattle at 3–24 months of age. All slaughterhouses were ranked by the throughput of interested cattle population, and predetermined numbers of animals to be sampled were distributed according to the proportional throughput from the previous year. Starting with the slaughterhouses of largest throughput, those covering at least 80% of the national throughput were included. The number of slaughterhouses enrolled in the monitoring program varied slightly in three consecutive years, 2011–2013, including 27, 26, and 23, respectively. Animals to be sampled were chosen randomly throughout the year and only one animal per slaughterhouse was sampled per selected day. Furthermore, the request that the animals to be sampled had to originate from different herds was followed.
Isolation of Escherichia coli O157
All samples were analyzed for the presence of E. coli O157 using the procedure described in ISO 16654 (ISO 2001). Shortly, 90 mL of prewarmed (37 ± 1°C) modified Tryptone Soya Broth supplemented with novobiocin 20 mg/L (Oxoid) was added to the sponge sample and homogenized in Stomacher blender for 60 s. Following the selective enrichment (6 ± 0.5 h and 21 ± 3 h at 41.5 ± 0.5°C), immunomagnetic separation (IMS) was performed using Captivate™ anti-O157 beads (LabM Ltd.) according to the manufacturer's instructions. Fifty microliters of IMS suspension was streaked onto the sorbitol MacConkey agar containing cefixime–tellurite supplement (Oxoid) and CHROMagar™ STEC agar (CHROMagar) plates. Inoculated media were incubated at 37 ± 1°C for 21 ± 3 h. Non sorbitol-fermenting colonies on cefixime tellurite sorbitol MacConkey (CT-SMAC) agar and mauve-colored colonies on chromogenic medium were subcultured and tested for the production of β-glucuronidase and indole by conventional biochemical analyses.
Serological confirmation
Serotyping of the presumptive E. coli O157:H7 isolates was performed by conventional agglutination tests using O157 and H7 antisera (Statens Serum Institute) according to the manufacturer's instructions. To exclude false-positive O157 agglutination, the presence of O157 serogroup was confirmed by agglutination of the boiled culture of the strain (100°C, 1 h).
Detection of the enterohemolytic phenotype
The enterohemolytic phenotype was detected on blood agar plates containing 10 mM calcium chloride and 5% defibrinated washed sheep erythrocytes as described by Beutin et al. (1989). The plates were examined after 24 h of incubation at 37 ± 1°C for the presence of lysis areas around the bacterial colonies.
Antimicrobial susceptibility testing
The susceptibility to antimicrobials was determined by the minimal inhibitory concentration microdilution method using VetMIC GN panel (version 4) as described by the manufacturer (National Veterinary Institute). The antimicrobial agents tested were as follows: ampicillin, ciprofloxacin, nalidixic acid, gentamicin, streptomycin, tetracycline, florfenicol, colistin, sulfamethoxazole, trimethoprim, chloramphenicol, kanamycin, cefotaxime, and ceftazidime. Based on epidemiological cutoff values, the results were categorized as susceptible or resistant according to the guidelines of European Committee on Antimicrobial Susceptibility Testing and EU Reference Laboratory for Antimicrobial Resistance (EURL-AR 2013).
Detection of stx1, stx2, and eae genes by polymerase chain reaction
Total DNA was extracted from pure bacterial cultures using a commercial DNA extraction kit (RTP® Bacteria DNA Mini Kit; Stratec Molecular GmbH) as described by the manufacturer and used as template in real-time PCR assays.
The primers, probe systems, and real-time PCR assay conditions described in ISO technical specification ISO/TS 13136 (ISO 2012) were used for the identification of the virulence genes stx1, stx2, and eae. The primers and probes were obtained from Eurogentec S.A. Real-time PCR amplification for each gene was performed in a final volume of 25 μL containing 12.5 μL of AmpliTaq Gold 360 Master Mix (Applied Biosystems®; Thermo Fisher Scientific, Inc.), 5 μL of primer mix (final concentration of each 1 μM), 4 μL of sterile water, 3 μL of template DNA, and 0.5 μL of probe with final concentration 0.2 μM. Each sample was run in duplicate. DNA extracted from the reference Escherichia coli O157 strain, EDL 933, was used as positive control, and sterile water was used as negative control. Real-time PCRs were performed using the real-time PCR cycler. The thermal cycling was carried out with the pre-PCR activation at 50°C for 120 s and at 95°C for 600 s following 40 cycles of denaturation at 95°C for 15 s, annealing and extension at 60°C for 60 s, and final extension at 72°C for 120 s.
Detection of stx1 and stx2 gene subtypes by conventional PCR
Conventional PCR for the detection of sxt1 and stx2 gene subtypes was performed as described by Scheutz et al. (2012).
Typing by PFGE
The genetic relationship among the O157:H7 isolated strains was investigated by PFGE in accordance with the standard operating procedure of EFSA external scientific report (Caprioli et al. 2014). The restriction profiles obtained with XbaI were analyzed with the BioNumerics software, version 7.1 (Applied Maths). Each PFGE pattern was given a number within the serotype O157:H7, for example, EC01, EC02, and EC03. The patterns were compared using Dice's similarity coefficient with tolerance and optimization values at 1.5%. Strains were deemed to have the same PFGE patterns when all bands were 100% identical. The dendrogram was built using the unweighted pair-group method with arithmetic averages.
Statistical analysis
The prevalence and CI based on probabilities derived from binominal distribution for proportion were calculated using the software VassarStats (Lowry 2014). The statistical significance of the difference in prevalence between different groups was investigated using Fisher's exact test to the respective reference. A p-value <0.05 by Fisher's exact test was considered statistically significant.
Results
E. coli O157 was isolated from 30 (4.0%; CI95 2.8–5.7%) of 744 hide swabs of cattle collected within a 3-year monitoring program. No statistically significant difference was found in terms of E. coli O157 prevalence in the three consecutive years (Table 1). Two-third of the positive animals (n = 19) were slaughtered at three main slaughterhouses located in different geographical regions of the country. E. coli O157-positive cattle originated from 26 farms, located in 12 of the15 counties in Estonia.
E. coli O157 was isolated more often after the 6-h enrichment step compared with an enrichment time of 18–24 h (p < 0.0001). There was only one sample found to be negative in 6-h time point, but positive after 18–24 h. Twenty-nine samples were found to be positive after 6-h enrichment.
All isolates agglutinated with H7 antiserum showed enterohemolytic phenotype and possessed the intimin-coding eae gene. Statistically significant difference (p < 0.0001) was found between the occurrences of the stx gene families, stx1 and stx2, among the 30 isolates: 13 (43.3%) of them carried the stx2 gene only, whereas the remaining 17 (56.7%) harbored both the stx1 and stx2 genes. Stx2c was the most frequently detected stx subtype, occurring alone (n = 12, 40.0%) or in combination with subtype stx1a (n = 13, 43.3%). Subtype stx2a alone was detected in one isolate and in combination with stx1a in four isolates. None of the isolates harbored genes of the stx1c, stx1d, stx2b, stx2d, stx2e, stx2f, and stx2g subtypes.
Comparison of PFGE profiles showed that the isolates presented an overall similarity of 76.9%. In total, 20 different XbaI PFGE patterns were differentiated among the 30 isolates, of which 14 grouped into four clusters based on a genetic relatedness criterion of 100% (EC14, EC01, EC04, and EC07, Fig. 1). The number of isolates belonging to the same cluster ranged from two to five. Cluster EC14 grouped two isolates from year 2013, whereas the other three clusters included strains isolated in 2011 and 2012. Both strains showing the EC14 pattern were isolated from cattle originating from the same locality, but from different farms and slaughtered in different facilities. Sampling time was different as well. Clusters EC04 and EC07 showed more than 97% similarity to other PFGE patterns of isolates from 2012 originating from geographically dispersed localities.
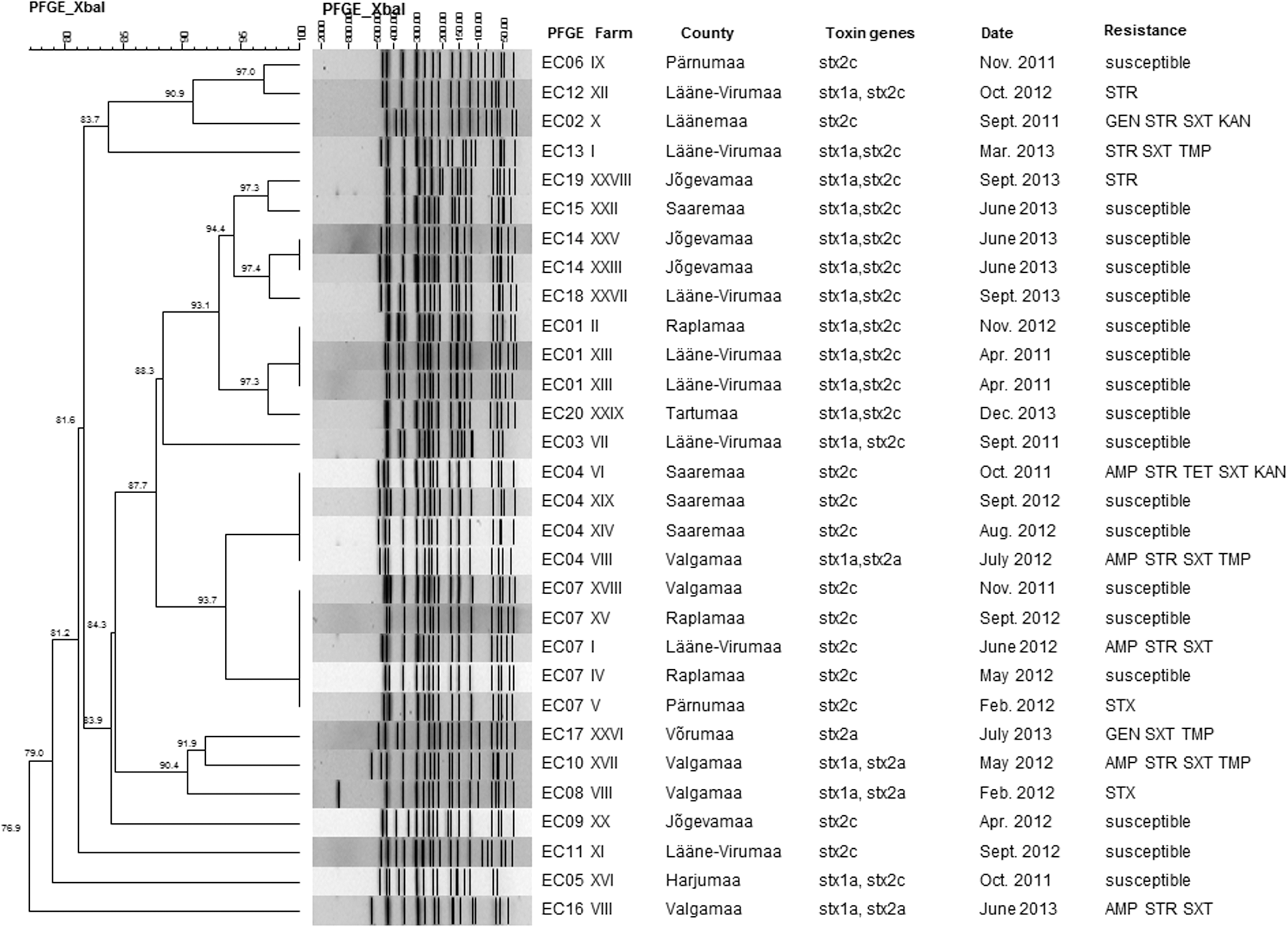
Dendrogram of the pulsed-field gel electrophoresis profiles obtained after XbaI digestion of DNA from Escherichia coli O157:H7 isolates from cattle hide samples during the period 2011–2013. The scales at the top indicate the similarity indices and molecular sizes in percentages and kilobases, respectively. STR, streptomycin; GEN, gentamicin; SXT, sulfamethoxazole; TMP, trimethoprim; AMP, ampicillin; TET, tetracycline; KAN, kanamycin.
Generally, the PFGE profiles reflected the stx gene subtypes, with the exception of profile EC04 (Fig. 1). Three isolates recovered from the same locality harbored the stx2c gene, whereas one isolate having the same PFGE pattern and originating from the nonadjoining county carried genes of the stx1a and stx2a subtype. The determination of toxin subtype genes for the latter isolate was repeated and results confirmed the presence of stx1a and stx2a genes. The high diversity of PFGE patterns among three E. coli O157:H7 strains isolated from cattle of the same farm and harboring both stx1a and stx2a genes was observed.
Table 2 shows the distributions of the minimal inhibitory concentration (MIC) values for the 30 E. coli O157:H7 isolates. All the isolates were susceptible to nalidixic acid, ciprofloxacin, florfenicol, colistin, chloramphenicol, cefotaxime, and ceftazidime. Eighteen isolates (60.0%) were sensitive to all the tested antimicrobials, while resistance to one or more agents was detected in 12 (40.0%) isolates.
The MIC values for the isolates were evaluated according to the instructions given by the manufacturer (National Veterinary Institute, Uppsala, Sweden).
Percentage of resistant isolates. Bold values indicate epidemiological cutoff values between sensitive and resistant isolates.
The number of isolates with MIC values less than or equal to VetMIC GN-panel minimum concentration range.
The number of isolates with MIC values more than VetMIC GN-panel maximum concentration range.
AA, antimicrobial agents; AMP, ampicillin; CIP, ciprofloxacin; NAL, nalidixic acid; GEN, gentamicin; STR, streptomycin; TET, tetracycline; FFC, florfenicol; CST, colistin; SXT, sulfamethoxazole; TMP, trimethoprim; CHL, chloramphenicol; KAN, kanamycin; CTX, cefotaxime; CAZ, ceftazidime; MIC, minimal inhibitory concentration.
Multiple drug resistance, which was defined as resistance to three or more unrelated antimicrobials, was detected in eight (26.7%) isolates. A high proportion of isolates were resistant to streptomycin (33.3%) and sulfamethoxazole (33.3%). Resistance against ampicillin, trimethoprim, kanamycin, tetracycline, and gentamicin was found for five (16.7%), four (13.3%), two (6.7%), one (3.3%), and one (3.3%) isolate(s), respectively. The highest MIC values (Table 2) were obtained for sulfamethoxazole, streptomycin, and ampicillin.
Coincidence between PFGE patterns and antimicrobial susceptibility was found among isolates in profiles EC01 and EC14. Conversely, isolates belonging to the PFGE profiles, EC04 and EC07 (Fig. 1), showed different resistance profiles.
Discussion
Contaminated bovine meat is considered to be a major source of foodborne STEC infections in humans, particularly those sustained by E. coli O157:H7 (Farrock et al. 2013). Sampling cattle hides at slaughterhouse for STEC monitoring is advisable since the highest prevalence is expected at this stage of production and hides represent a major source of carcass contamination by E. coli O157 (Keen and Elder 2002, O'Brien et al. 2005).
In the present study, the prevalence of STEC O157:H7 on cattle hides ranged from 3.3% to 5.3% in three consecutive years. The studies on the prevalence of E. coli O157 in cattle at slaughter performed in several countries are not fully comparable because of the differences in sampling site and time, sample type and size, study population, and analytical methods. In Italy, Bonardi et al. (2015) reported a 2.5% prevalence of STEC O157 in cattle hides. However, the samples were taken from 100-cm2 brisket area instead of 400 cm2. The studies performed by O'Brien et al. (2005) and Thomas et al. (2012) on Irish cattle hides in various years showed a prevalence of 7.3% and 13.4%, respectively. In a monitoring program carried out in Sweden, Boqvist et al. (2009) reported an E. coli O157 prevalence of 12%. However, in this study, swabs were taken from ears instead of the brisket area.
All the E. coli O157:H7 isolates obtained in the present study possessed the intimin-encoding eae gene. The stx2c gene, alone or in combination with stx1a, was the most frequently detected stx subtype, similar to what was reported in studies by Lefebre et al. (2009) and Bonardi et al. (2015). The stx1a, stx2a, stx2c, and stx2d subtypes were similarly reported in STEC strains from cattle and beef meat products (Martin and Beutin 2011), and the first three have also been detected in the present study. The same subtypes have been reported in STEC O157 strains from Swedish cattle, and potential links to human infections have been described by Aspan and Eriksson (2010). It is noteworthy that STEC strains possessing stx2a and stx2c subtypes have been associated with hemorrhagic colitis and HUS in humans (Persson et al. 2007).
The E. coli O157:H7 isolates obtained from the monitoring program were characterized by PFGE, a broadly used method for molecular typing of E. coli O157 (Caprioli et al. 2014) and, more generally, for investigating bacterial transmission throughout the food production chain and for identifying clusters of STEC strains linked to outbreaks (Louie et al. 1999, Thomas et al. 2012). The results of the present study showed that cattle originating from geographically dispersed farms can carry STEC O157:H7 strains with an indistinguishable PFGE pattern. This could be explained by the presence of STEC O157:H7 clones circulating in a small area of the country and finite cattle population. On other hand, we cannot exclude that cross-contamination may have occurred during subsequent stages, such as at slaughter level. Significant cross-contaminations between animals during transportation and housing in a lairage at slaughterhouse have been demonstrated by Mather et al. (2008).
Further characterization of the isolates was obtained by determining their antimicrobial susceptibility pattern. Resistance to streptomycin, sulfamethoxazole, and tetracycline has been frequently found among different STEC strains from animal and human origin (Morabito et al. 2002, Ziebell et al. 2011). The use of antibiotics for prophylactic purposes or in therapy can select resistant bacteria, and multiresistant strains might reflect the history of antimicrobial usage (Smith et al. 2007). This phenomenon might partly explain the high proportion of E. coli O157:H7 isolates (33.3%) that were resistant against streptomycin and sulfamethoxazole in the present study. High resistance was also found against ampicillin (16.7%) and trimethoprim (13.3%). On the contrary, no fluoroquinolone resistance and extended-spectrum beta-lactamase were detected. Multiple drug resistance was observed in 26.7% of our E. coli O157:H7 isolates, the most frequent resistance pattern being streptomycin, sulfamethoxazole, and ampicillin. These results seem to be consistent with those of previous studies reporting multiple drug resistance in STEC O157 isolates from cattle origin (Mora et al. 2005, Scott et al. 2009).
In conclusion, the current study provides a first description of the E. coli O157:H7 strains circulating among Estonian cattle, including their virulence gene profiles and stx gene subtypes. STEC O157:H7 isolates with indistinguishable PFGE pattern were found from cattle originating from different localities and in different years, suggesting that some persistent clones are present in Estonian beef production chain. The high proportion of strains showing multiple drug resistance may indicate problems caused by an inappropriate use of antimicrobials in the Estonian cattle herds. Up to now, STEC state monitoring in Estonia has been focused only on E. coli O157, but other STEC serogroups are also important in STEC epidemiology and should be included into the national monitoring program.
Footnotes
Acknowledgments
This study was supported by the applied research project T13057VLTH of the Ministry of Agriculture of Estonia. The authors would like to thank the Veterinary and Food Board for its cooperation and the colleagues from Veterinary and Food Laboratory for valuable laboratory assistance. Sincere thanks are extended to Dr. Alfredo Caprioli (ISS, Rome, Italy) and his colleagues for genotyping the E. coli isolates.
Author Disclosure Statement
No competing financial interests exist.