
Abstract
Introduction:
Toscana virus (TOSV) is a sandfly-borne bunyavirus with a significant public health impact. Preliminary studies have revealed TOSV exposure in dogs and they were suggested as potential reservoirs. This study was performed to characterize canine TOSV infections in an endemic region. Sequencing of TOSV small (S) segment in several previously identified specimens was also undertaken to reveal viral genealogy.
Materials and Methods:
Canine and feline plasma were collected in several districts of Mersin province, Mediterranean Anatolia, Turkey, during May–September, 2015. Phlebovirus RNA was screened through two nested polymerase chain reaction (PCR) assays, targeting S and large (L) segments of the viral genome. A kinetoplast minicircle nested PCR was employed for Leishmania DNA detection and typing. Previously collected TOSV-positive specimens from humans, dogs, cats, and sandflies from various regions in Turkey and Cyprus were further evaluated through the S segment PCR. All amplicons were characterized through sequencing.
Results:
A total of 210 specimens that comprise canine (76.2%) and feline (23.8%) plasma were screened. In three (1.9%) and two (1.3%) canine specimens, TOSV and Leishmania nucleic acids were detected, respectively. The TOSV strains were characterized as genotype B, and Leishmania infantum was identified in positive specimens. Twenty-four partial S segment sequences were amplified, which demonstrated a maximum intramural diversity of 3.88% in the nucleotide level. Sequence comparisons revealed significant similarities to particular genotype B strains characterized in Spain and France, whereas a notable divergence was observed among several TOSV strains. Single or recurrent amino acid substitutions were noted in eight residues of the viral nucleocapsid.
Discussion:
Canine infections of TOSV genotype B, with temporal and spatial association with L. infantum, were detected. Divergent TOSV S segment sequences with amino acid substitutions, presumably associated with host adaptation, were observed.
Introduction
A
Phleboviruses are associated with the flagellate protozoans of the Leishmania species, the causative agents of cutaneous, mucocutaneous, and visceral leishmaniasis, due to the shared sandfly vectors (Maroli et al. 2013). Several studies have revealed a probable epidemiologic interaction among these agents, as well as coinfected sandflies (Bichaud et al. 2011, Ergunay et al. 2014, Es-Sette et al. 2014, Faucher et al. 2014). Crucial issues on the TOSV natural cycle and disease epidemiology have not yet been completely elucidated, and virus exposure in vertebrate species other than humans has only been superficially explored (Navarro-Mari et al. 2011). Recently, canine and feline TOSV infections as well as Leishmania and TOSV coinfected dogs in the Mediterranean region of Turkey have been reported (Dincer et al. 2015). Moreover, the findings suggested canine species to act as a potential reservoir or amplifying host for TOSV, a concept similar to that of leishmaniasis, where the dogs may serve as primary domestic reservoirs. This study was planned and executed as a follow-up and confirmatory effort for the initial report, to identify and characterize ongoing TOSV infections in canine and feline species in an area endemic for leishmaniasis and phlebovirus exposure (Ok et al. 2002, Tezcan et al. 2015). As a secondary objective, characterization of partial S segment of TOSV, for which very limited information is available in Turkey, was performed in several archival specimens.
Materials and Methods
Study area, sample collection, and processing
The study was undertaken during May–September, 2015, in Akdeniz, Mezitli, Pozcu, Tasucu, and Yenisehir districts of Mersin province (36°48′N, 34°38′E), Mediterranean Anatolia, Turkey. Human phlebovirus/TOSV exposure was previously identified in the region and having domestic or farm animals has been reported to be associated with an increased risk for virus exposure (Ergunay et al. 2012, Tezcan et al. 2015). Plasma from dogs and cats was obtained from local veterinary clinics and animal shelters. Specimen collection was conducted with full compliance with the national regulations on the operation and procedure of animal experiments ethics committees (Regulation No. 26220, date: July 09, 2006), and the protocols were approved by Ankara University Local Ethics Committee (No. AULEC/201-96-346). All specimens were aliquoted and stored at −80°C until processing.
Nucleic acid purification in the collected specimens was carried out using High Pure Viral Nucleic Acid Kit (Roche Diagnostics). Subsequent reverse transcription reactions were undertaken through randomized hexamers, employing RevertAid First Strand cDNA Synthesis Kit (Thermo Scientific).
Archival specimens
Purified nucleic acids or complementary DNAs from previously collected specimens, originating from human, dog, cat, or sandflies, with detectable TOSV RNA (Ergunay et al. 2011, 2014, Erdem et al. 2014, Ocal et al. 2014, Dincer et al. 2015), were evaluated for the partial characterization of viral nucleocapsid sequences, as described hereunder. These specimens had been processed identically as the screening samples.
Detection of Phlebovirus and Leishmania nucleic acids
Two previously described nested polymerase chain reactions (PCRs), targeting the L and S segments of the viral genome, were employed for phlebovirus RNA screening in the collected specimens (Sanchez-Seco et al. 2003, Charrel et al. 2007). The assays were optimized using Vero cell-grown TOSV ISS.Phl.3 isolate. The conserved region of the kinetoplast minicircle of the Leishmania species was targeted through a previously published nested PCR method (Noyes et al. 1998). The assay detects several clinically relevant Leishmania species, with species characterization based on the product size. All expected amplicons were observed under ultraviolet light after electrophoresis in 1.5–1.7% agarose gels.
Sequencing and phylogenetic analyses
The cleanup of the amplification products was undertaken through High Pure PCR Product Purification Kit (Roche Diagnostics). Subsequently, the sequencing reactions were performed using an ABI Prism 310 Genetic Analyzer (Applied Biosystems), with forward and reverse primers employed for the second step of each PCR. Obtained reads were aligned and analyzed using Bioedit v7.0.9.0 (Hall et al. 1999), CLC Main Workbench v7.6.4 (CLCBio), and MEGA software v6.0 (Tamura et al. 2013). Nucleotide and protein similarity searches were performed using the megablast, blastn, and blastp tools of the GenBank (
Results
Canine and feline specimens
A total of 210 specimens that comprise 160 (76.2%) canine and 50 (23.8%) feline plasma were collected throughout the 5-month sampling period. Monthly distribution of the specimens was recorded as 25 (11.9%) in May, 49 (23.3%) in June, 71 (33.8%) in July, 53 (25.2%) in August, and 12 (5.7%) in September (Table 1).
TOSV, Toscana virus.
Phlebovirus and Leishmania PCR in animal specimens
Three specimens (3/210, 1.42%) revealed detectable products for the phlebovirus L and S segment PCRs during the screening. They originated from separate canine specimens, collected in May, July, and September, 2015 (Table 1). Viral nucleic acids were not detected in any other canine or feline specimen (207/210, 98.6%). No prominent clinical symptom was present in any of the PCR-positive dogs.
Leishmania PCR was positive in two canine samples (2/210; 0.95%), collected during May and July, 2015 (Table 1). These were characterized as Leishmania infantum, through amplicon size and sequencing (data not given). The animals with detectable parasite DNA had skin lesions on various legs and back, suggesting cutaneous leishmaniasis. No codetection of Phlebovirus and Leishmania nucleic acids was noted.
Phlebovirus PCR in archival specimens
Amplification of the S segment in previously collected specimens revealed positive results in 21 out of 34 cDNAs (61.7%). The reactive samples comprise 14 canine and 1 feline specimens collected in 2013 as well as three human plasma or urine specimens collected in 2012 and 2015 from central Anatolia and eastern Thrace regions (Table 2).
L, large; S, small.
Analysis of Phlebovirus L segment sequences
Sequencing of the L segment PCR products (GenBank accession numbers: KU248879, KU248880, and KU248881) revealed all three canine specimens to group with TOSV genotype B strains (Fig. 1), with an intramural nucleotide divergence of 2.06–3.09%. Diversity rates of 1.04–2.07% were also noted with previously identified genotype B strains from the same region (strains N7 and N9, Fig. 1). Moreover, the sequences demonstrated 94.82–97.42% similarity to various genotype B strains from France and Spain, as well as 80.41–81.96% similarity to genotype A strains from Turkey, France, and Italy. Maximum likelihood analysis supported genotype distribution and phylogenetic grouping with high bootstrap values (Fig. 1).
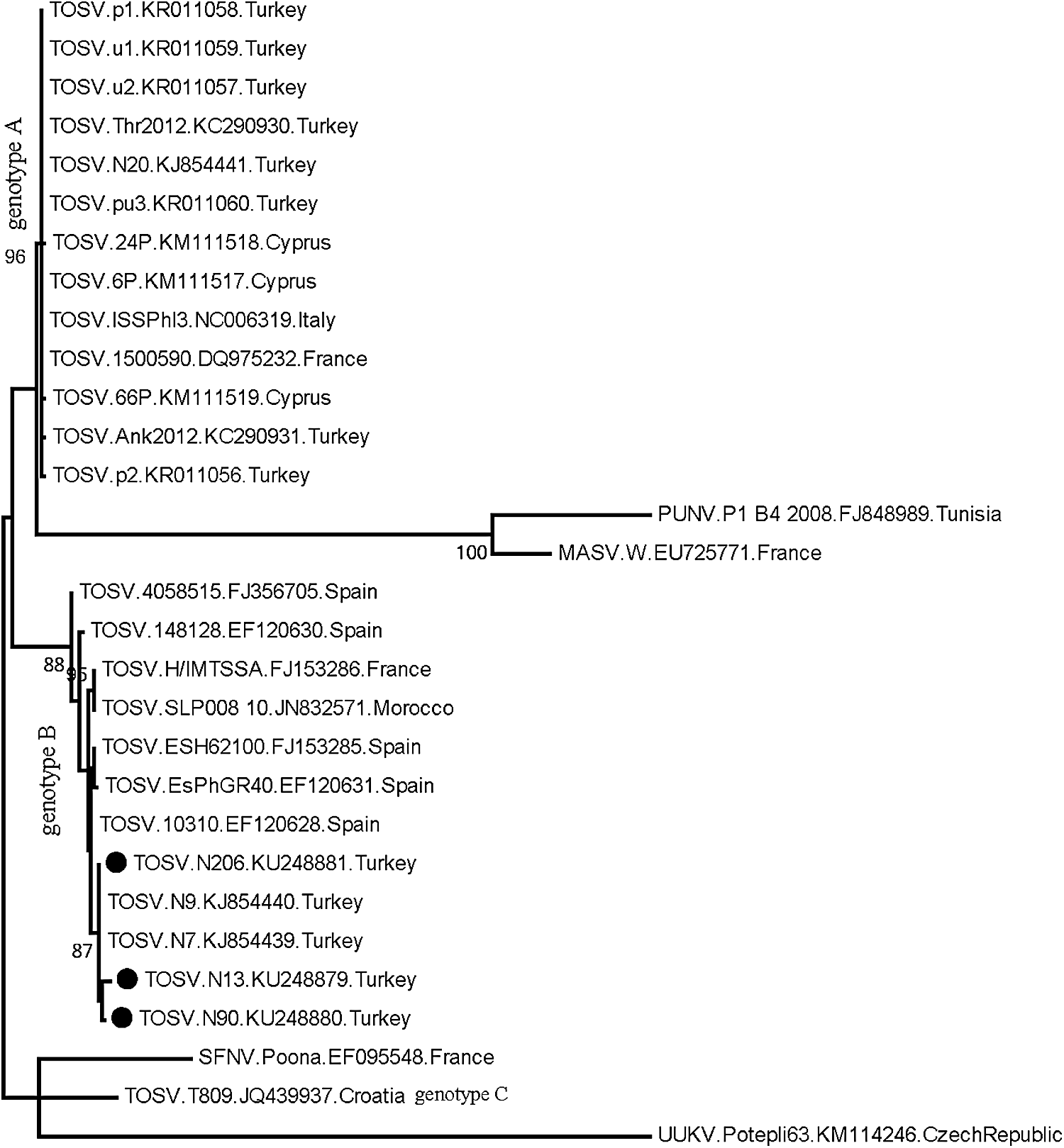
Maximum likelihood analysis of the phlebovirus partial L segment nucleotide sequences. The evolutionary distances were computed using the Kimura 2-parameter model. Bootstrap values lower than 60 are not shown. Viruses in the analysis are indicated with name, isolate identifier, GenBank accession number, and country of detection. Black circles indicate the sequences characterized in this study. Uukuniemi virus isolate Potepli63 is included as an outlier. L, large; MASV, Massilia virus; PUNV, Punique virus; SFNV, Sandfly Fever Naples virus; TOSV, Toscana virus; UUKV, Uukuniemi virus.
Analysis of Phlebovirus S segment sequences
In addition to the current sequences from dogs (n: 3), a total of 21 partial S segment sequences were characterized in specimens originating from various sources and regions (GenBank accession numbers: KU248882–KU248902) (Table 2). All animal sequences were significantly conserved, showing intramural similarity rates of 96.60–100%. No prominent association of sequence variation rate or divergent positions with virus genotype was observed (data not shown). Comparison with sequences from clinical specimens also revealed a maximum diversity of 3.88% (eight nucleotides). However, one sequence from central Anatolia, previously characterized in a patient in 2011 (KE4) (Ergunay et al. 2011), stood out from the group, with 86.44–92.37% identity matches.
Comparison of the sequences with several TOSV strains from different endemic regions revealed 34.11–39.35% similarity rates. However, certain genotype B viruses from France and Spain (strains AR, 10310, D320390, 159586, and 148128) were observed to be closely related to the Turkish strains, with significantly higher (96.12–99.03%) nucleotide identity matches. These observations were further reflected in the maximum likelihood analysis, where these strains cluster with sequences from Turkey, distinct from other TOSV sequences (Fig. 2). Various analysis models further supported these findings (data not given).
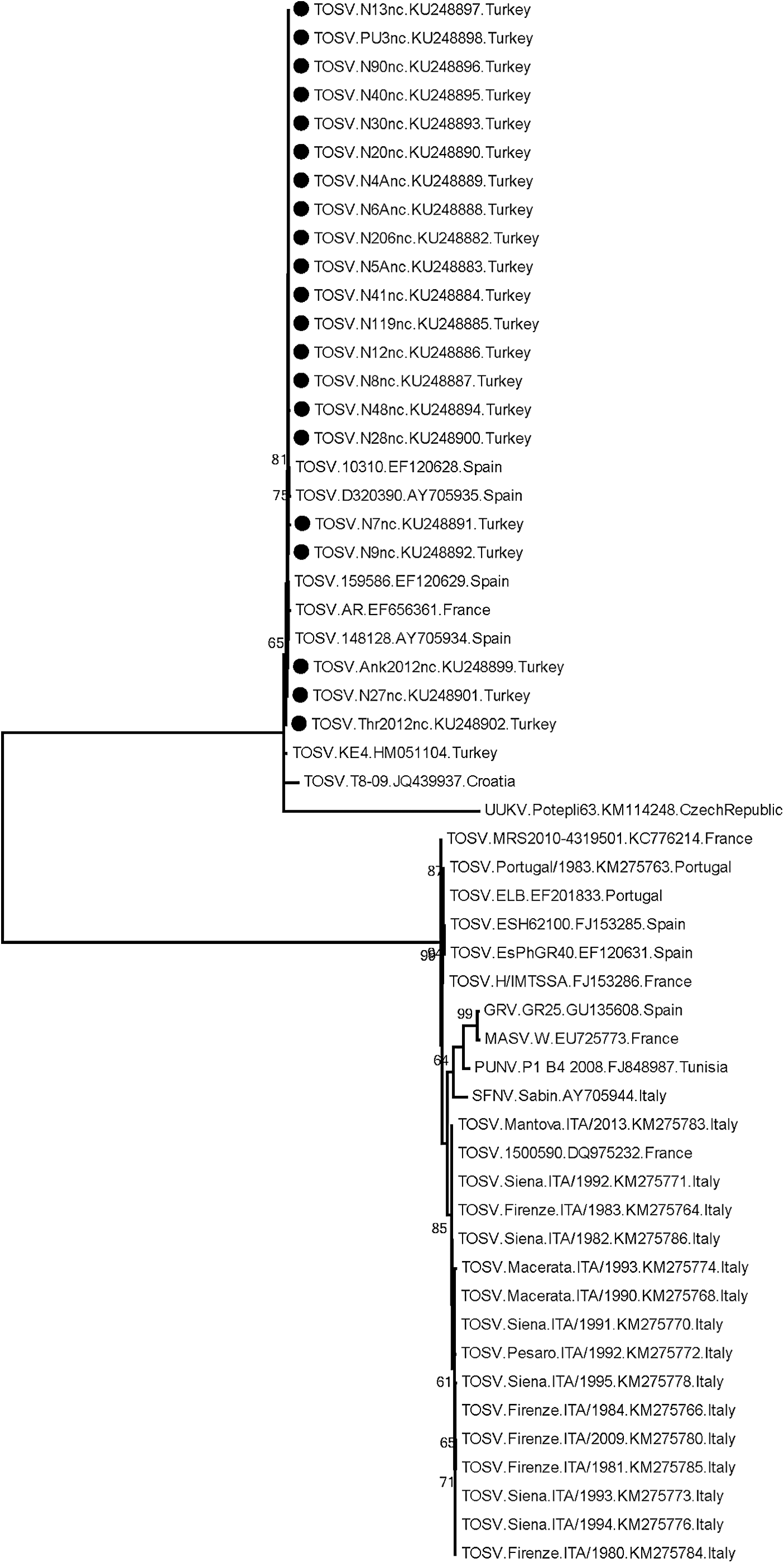
Maximum likelihood analysis of the phlebovirus partial S segment nucleotide sequences. The evolutionary distances were computed using the Kimura 2-parameter model. Bootstrap values lower than 60 are not shown. Viruses in the analysis are indicated with name, isolate identifier, GenBank accession number, and country of detection. Black circles indicate the sequences characterized in this study. Uukuniemi virus isolate Potepli63 is included as an outlier. GRV, Granada virus; S, small.
Partial amino acid sequences of nucleocapsid protein, corresponding to the amino acids 76 to 208 of the TOSV genotype A strain TOSV/Firenze.ITA/1983/2, were further analyzed (Table 3). A maximum of three amino acid variations were noted among strains identified in Turkey (between N206 and N12 strains in positions 81, 100, and 101), including the previously characterized KE4 isolate. Substitutions were observed in eight positions (81, 95, 100, 101, 113, 125, 133, and 146). The substitutions S81P, Y95S, S113P, and Y146 were noted more than one isolate and one divergent residue (in position 100) were identified among global strains (Table 3).
Viral sequences are designated by strain name and country of origin. Positions are given according to the alignment of the complete nucleocapsid protein of TOSV genotype A strain, TOSV/Firenze.ITA/1983/2 (GenBank accession AIQ84538). All other virus genotypes, strains with higher similarity on the nucleotide level, and the previously characterized sequence from Turkey (KE.4) were also included. Substitutions detected in the study are in italics and bold. Positions with identical residues in all strains are excluded. Amino acids not covered by the available sequences are shown as blank (dashes).
A, alanine; C, cysteine; D, aspartic acid; E, glutamic acid; F, phenylalanine; G, glycine; H, histidine; I, isoleucine; K, lysine; N, asparagine; P, proline; S, serine; T, threonine; V, valine; Y, tyrosine.
Discussion
TOSV infections in dogs and cats have recently been reported, with implications for about the potential reservoir capability of dogs, as observed in canine leishmaniasis (Bichaud et al. 2011, Navarro-Mari et al. 2011, Es-Sette et al. 2014, Dincer et al. 2015). One goal of this study was to investigate TOSV in canine and feline specimens to clarify their role as probable reservoirs in the region where the initial detection of canine TOSV infections was documented (Dincer et al. 2015). Nested PCR assays targeting different regions in the viral genomes were employed for viral RNA detection. Serological screening was not performed, for data on previous exposure could only be provided by this approach.
Among the 210 specimens collected during the sandfly-active season in 2015, phlebovirus RNA could be detected in a total of 3 canine specimens (1.42%). The infecting strains were characterized through sequencing as TOSV, confirming the occurrence of ongoing infection or viremia in dogs. The initial documentation from Mediterranean Anatolia comprised mainly dogs, where viral RNA could be detected in 15.5% of the specimens (Dincer et al. 2015). The current results demonstrate new infections in dogs, although with a lower incidence. Similar to the previous report, no clinical symptoms ascribable to TOSV were observed in animals with detectable viral nucleic acids. Previous investigation of the viremic animal specimens has suggested a continuous transmission from June to September, which also seemed to coincide with the emergence of human cases (Dincer et al. 2015). The findings in this study also confirm this observation and demonstrate an even earlier virus activity, becoming detectable in May (Table 1). Since TOSV has not yet been detected in sandflies from Turkey, a temporal and spatial analyses of the infected vectors with vertebrate infections remain to be performed. Previous reports on animal infections are scarce, with low-level viremia identified in a goat from Spain, and virus isolation from neuronal tissues of a Pipistrellus kuhli bat from Italy (Verani et al. 1988, Navarro-Mari et al. 2011). Nevertheless, TOSV as well as other phlebovirus exposure could be revealed through serology in several endemic countries, lately in Algeria (Tahir et al. 2015). Investigation for viremia through viral nucleic acid detection and isolation studies must be performed in other regions with TOSV circulation to elucidate the occurrence and dynamics of the infections in dogs.
Canine leishmaniasis was detected in two dogs with cutaneous lesions in this study. The infecting strains were characterized as L. infantum, which is known to be endemic in the region and was previously detected in coinfections (Dincer et al. 2015). Although no TOSV–Leishmania coinfections were detected, the epidemiological connection is evident, with the seasonal co-occurrence of the infections (Table 1).
Sequence divergence in L and M genomic segments of various TOSV strains has been revealed, and the circulation of two distinct virus lineages or genotypes has been characterized. The strains detected in Italy and Iberian Peninsula are denoted as genotypes A and B, respectively, whereas the occurrence of another lineage, tentatively called as genotype C, is identified in Croatia (Collao et al. 2009, Charrel et al. 2012, Punda-Polic et al. 2012). In some regions such as southeastern France, genotypes A and B are shown to cocirculate (Charrel et al. 2007). In this study, all TOSV strains identified in dogs are characterized as genotype B, according to the partial L segment sequences, with significant similarities to the sequences of the same genotype reported previously (Fig. 1). So far, TOSV genotype A has been identified in all human infections from Turkey (Ergunay et al. 2011, 2015, Erdem et al. 2014, Ocal et al. 2014). However, circulation of TOSV genotype B was reported for the first time in dogs in 2013 from the Mediterranean Anatolia, as well as presence of genotypes A and B within the same region (Dincer et al. 2015). The findings confirm the activity of genotype B strains, detected 2 years after the initial report in the region. It appears that the distribution of virus genotype within a particular area is dynamic, and introduction and elimination of virus genotypes are possible, which is presumably affected with the sandfly species active in the region.
To provide information on the nucleocapsid gene and the genetic variability of the circulating TOSV isolates in Anatolia, stored archival RNA/cDNAs were evaluated through the nested S segment PCR, coemployed for screening. Partial sequences were characterized in 21 specimens originating from serum, plasma, and urine of canine, feline, and human subjects, collected in various regions of Anatolia during 2012–2015 (Table 3). Regardless of the virus genotype, these sequences were observed to be significantly conserved. To date, only a partial stretch of 153 nucleotides of the TOSV S genomic segment, originating from serum of a patient presenting with meningoencephalitic symptoms in 2011, has previously been available from Turkey (Ergunay et al. 2011). The alignment of this sequence with the current sequences demonstrated a more emphasized divergence. Comparison of the partial S segment sequences from Anatolia with those from various regions with endemic TOSV activity revealed similarities comparable with the intramural diversity of currently explored Turkish sequences and with particular genotype B strains characterized in Spain and France (Fig. 2). Conversely, other genotype B strains originating mostly from the Iberian Peninsula as well as genotype A strains revealed notable divergence, with a maximum similarity of 82.04%. Phylogenetic analyses further supported these observations, where genotype B isolates closely related to the Anatolian sequences are clustered as separate clades, distinct from other TOSV strains and genotypes (Fig. 2).
Previous analyses of complete/partial S segment sequences from Italy and Spain, with the predominance of one particular virus genotype, revealed limited variability in nucleotide and amino acid sequences (Collao et al. 2009, Baggieri et al. 2015). The findings in this study reveal that the genetic variation is more pronounced in TOSV strains identified in Anatolia. This is further supported by the circulation of the two major virus genotypes, also confirmed for genotype B in this study, even within the same surveillance zone. Furthermore, nucleotide variations leading to substitutions were observed in 8 residues of the available 132 amino acid regions (Table 3). Certain substitutions were identified in more than one local strain and in other strains characterized in endemic regions. Interestingly, the Y146F substitution is present in new world phlebovirus isolates such as Buenaventura virus, CoAr 171616, CoAr 170255, and VP-334K, vectored by the Lutzomyia species of sandflies (data not shown). Thus, some of these substitutions can be hypothesized to reflect adaptive processes to host variation, probably to various sandfly species. It must be indicated that only partial sequences, provided through the amplicons of the screening assays, could be analyzed in this study, which constitutes the main limitation of the current effort. Virus isolation and complete genomic characterization are required to reveal the precise genealogy of the TOSV strains circulating in Anatolia.
Conclusions
Canine infections of TOSV as well as L. infantum have been identified, 2 years after the initial detection, in Mediterranean Anatolia, Turkey. Circulation of TOSV genotype B strains was demonstrated, confirming the presence of this lineage. Partial sequences of the S genomic segment revealed a relatively higher genetic diversity than other endemic regions, with several amino acid substitutions, presumably associated with host adaptation. Coinfections of TOSV and L. infantum in the same organism were not characterized, despite an observed temporal and spatial association of the two pathogens.
Footnotes
Acknowledgment
Preliminary findings of this study have been presented orally at the 1st National Virology Days, held during February 25th–28th, 2016, in Ankara, Turkey. The corresponding author is a recipient of the Georg Forster Research Fellowship (HERMES) for Experienced Researchers of the Alexander von Humboldt Foundation, 2015.
Author Disclosure Statement
No competing financial interests exist.