
Abstract
Yersinia enterocolitica and Yersinia pseudotuberculosis are important zoonotic bacteria causing human enteric yersiniosis commonly reported in Europe. All Y. pseudotuberculosis strains are considered pathogenic, while Y. enterocolitica include both pathogenic and nonpathogenic strains which can be divided into six biotypes (1A, 1B, and 2–5) and about 30 serotypes. The most common types causing yersiniosis in Europe are Y. enterocolitica bioserotypes 4/O:3 and 2/O:9. Strains belonging to biotype 1A are considered as nonpathogenic because they are missing important virulence genes like the attachment-invasion-locus (ail) gene in the chromosome and the virulence plasmid. The role of wild small mammals as a reservoir of enteropathogenic Yersinia spp. is still obscure. In this study, the presence of Yersinia spp. was examined from 1840 wild small mammals, including voles, mice, and shrews, trapped in Finland during a 7-year period. We isolated seven Yersinia species. Y. enterocolitica was the most common species, isolated from 8% of the animals; while most of these isolates represented nonpathogenic biotype 1A, human pathogenic bioserotype 2/O:9 was also isolated from a field vole. Y. pseudotuberculosis of bioserotype 1/O:2 was isolated from two shrews. The ail gene, which is typically only found in the isolates of biotypes 1B and 2–5 associated with yersiniosis, was frequently (23%) detected in the nonpathogenic isolates of biotype 1A and sporadically (6%) in Yersinia kristensenii isolates. Our results suggest that wild small mammals, especially voles, may serve as carriers for ail-positive Y. enterocolitica 1A and Y. kristensenii. We also demonstrate that voles and shrews sporadically excrete pYV-positive Y. enterocolitica 2/O:9 and Y. pseudotuberculosis 1/O:2, respectively, in their feces and, thus, can serve as a contamination source for vegetables by contaminating the soil.
Introduction
W
Enteropathogenic Yersinia spp., including Y. enterocolitica and Y. pseudotuberculosis, are important foodborne zoonotic bacteria causing enteric yersiniosis, which is commonly reported among humans in Europe (EFSA 2015). The mechanisms of pathogenicity in enteropathogenic Yersinia spp. are complex, including several chromosomally and plasmid-located genes (Revell and Miller 2001). One of the most important chromosomal virulence genes is the attachment-invasion-locus (ail) gene coding for the Ail outer-membrane protein responsible for the colonization and invasion of Y. enterocolitica to epithelial cells in the ileum during the initial stages of infection (Revell and Miller 2001, Mikula et al. 2013). Y. enterocolitica strains can be biochemically divided into six biotypes (1A, 1B, and 2–5) whose pathogenicity varies widely: biotype 1A strains are considered nonpathogenic due to the lack of most common chromosomal virulence genes and the approximately 70-kb virulence plasmid (pYV), which are found in both the weakly pathogenic strains of biotypes 2–5 and in the highly pathogenic biotype 1B strains (Bhagat and Virdi 2011). All Y. pseudotuberculosis strains are considered pathogenic. Most environmental Yersinia spp. strains are nonpathogenic. The ail gene is typically found only in pathogenic Y. enterocolitica strains and it is the most commonly used target for PCR detection of pathogenic Y. enterocolitica (Nakajima et al. 1992, Fredriksson-Ahomaa and Korkeala 2003, Lambertz et al. 2008). All Y. enterocolitica and Y. pseudotuberculosis strains causing yersiniosis in humans and animals carry the plasmid pYV, which is essential for these bacteria to survive and multiply in the lymphatic tissue (Cornelis et al. 1998).
The role of wild small mammals in the epidemiology of enteropathogenic Yersinia spp. is still obscure, even though several findings suggest the importance of small mammals. For instance, grated carrots and lettuce, possibly contaminated by wild small mammals or other wildlife, have been vehicles for Y. pseudotuberculosis epidemics in Finland (Nuorti et al. 2004, Jalava et al. 2006, Kangas et al. 2008, Rimhanen-Finne et al. 2009). In addition, Yersinia spp., including human pathogenic biotypes of Y. enterocolitica and Y. pseudotuberculosis, have been isolated from wild rodents in Japan, where rodents also seem to play an important role in the epidemiology of Y. enterocolitica bioserotype 1B/O:8 (Fukushima et al. 1990, Iinuma et al. 1992, Hayashidani 1995). Moreover, similar pathogenic Y. enterocolitica 4/O:3 strains were isolated from pigs and rodents on Swedish pig farms, but not from rodents outside the farms (Backhans et al. 2011). Laukkanen et al. (2008) suggested that the higher prevalence of Y. pseudotuberculosis on organic pig farms may be due to the higher number of pest and pet contacts with organic pigs than with pigs in in-house conventional farming. To predict human epidemics, the role of wild small mammals needs to be quantified.
The aim of our study was to clarify the role of small mammalian wildlife hosts as a reservoir for Yersinia spp.
Materials and Methods
Study area
The study was conducted in an area of Finland (60°–64°, 22°–29°, Fig. 1). Wild small mammals were trapped from 37 trapping locations (Tables 1 and 2). Most of trapping locations (30) were selected without earlier knowledge about the presence of Yersinia spp. and made two transects across Southern and Central Finland. Additional five locations (locations 31–35 in the Table 2 and Fig. 1) were source farms of Y. pseudotuberculosis epidemics a year before trapping started. The remaining two locations (locations 36–37) were specifically chosen for Borna virus research. All trapping locations are shown in Figure 1. Habitats in trapping areas covered typical boreal (taiga) forest (Norway spruce and birch and Scots pine) that covers most of the landscapes, wild meadows (abandoned old fields), and field edges.

Trapping sites in Finland. The number refers to trapping locations in the Table 2 and the size of the circle to the number of trapped animals.
Bold italic numbers include samples of ail-positive animals. Bold numbers include Yersinia-positive animals. Localities 31–35 were selected due to known earlier Yersinia spp. occurrence.
ail, attachment-invasion-locus gene.
Rodents and shrews
Small mammals were trapped after the reproductive season from September to November in 2001–2007. Voles undergo strong population cycles in Finland; generally, 2001 was an increase year, 2002 a peak year, 2003 was a crash, 2004 an increase, and 2006 a peak. The 30 trapping sites in the two transects across Finland remained the same through the study period with the exception that in the crash year of the rodent cycle in 2003 only every second locality was trapped. At each trapping locality, altogether 150 traps were set at five different sites, 30 traps in each, within 1 km2. Traps were set in the afternoon of day 1, checked and animals collected on day 2, and animals collected and traps removed on day 3. Most of the animals were trapped in the first night. At source farms of Y. pseudotuberculosis epidemics 200–300 snap traps were used for 1–2 days. Locations for Borna virus research were trapped once with 200 traps. Snap traps, which killed the animals immediately, were used. During the trapping months, nighttime temperatures were already low, from −2°C to +10°C and animals were frozen in dry ice immediately after trap checking. Oral cavity samples, including the tongue and larynx, and/or the intestine, including the end of the colon and rectum, were later dissected in the Rodent Laboratory of Finnish Forest Research Institute (now Natural Resources Institute Finland) and were subsequently investigated at the Department of Food Hygiene and Environmental Health, University of Helsinki. The animals were stored at −25°C before dissection. Intestinal and tongue samples were removed from partially thawed animals and immediately refrozen at −20°C before culturing.
Ethical statement
In Finland, no ethical permit is needed for snap trapping because the Finnish Act on the Use of Animals for Experimental Purposes (62/2006) and the Finnish Animal Experiment Board (16th May, 2007) do not classify snap trapping as an animal experiment. All trappings were done with the permission of landowners. A permit (23/5317/2001) for capturing protected species (mainly shrews) was granted by the Finnish Ministry of Environment. Other species captured are not protected in Finland, and none of the captured species are included in the Red List of Finnish Species.
Culture methods
The intestinal samples (including feces) and tongue samples were cut into small pieces with scissors and suspended in 4.5 mL of peptone-mannitol broth (PMB). Direct culturing after 4-h incubation at room temperature and cold enrichment for 7, 14, and 21 days at 4°C were carried out according to Laukkanen et al. (2008). After each incubation step, 100 μL of the PMB was spread on two selective agar plates: cefsulodin-irgasan-novobiocin (CIN; Oxoid) and MacConkey (Scharlau) plates. After cold enrichment the PMB was alkaline treated with 0.25% potassium hydroxide (KOH) solution for 20 s before spreading on selective agar plates. Small colonies (1 mm in diameter) with a deep red center surrounded by a colorless zone on CIN and small colorless colonies on MacConkey were collected after 20–24 h incubation at 30°C and after further incubation at room temperature for 24 h. The typical colonies were inoculated on Tryptic Soy Agar (TSA; Difco) plates that were incubated at 30°C for 24 h to create pure cultures for further identification. Up to four colonies on the TSA plates were tested for oxidase reaction (Pro-Lab Diagnostics), and oxidase-negative isolates were further inoculated on urea agar slants. Urea-positive isolates were identified using the API 20E test (BioMérieux) incubated at 25°C for 18–20 h.
Characterization
All suspected Yersinia spp. isolates identified by API 20E were biotyped using the following tests according to Wauters et al. (1987): esculin hydrolysis, tween-esterase activity and pyrazinamidase production, indole production, and salicin, xylose, and trehalose fermentation. Suspected Y. pseudotuberculosis isolates were biotyped with the raffinose and melibiose fermentation and citrate utilization tests (Tsubokura and Aleksic 1995). Furthermore, the isolates were characterized using a multiplex PCR assay targeting the 16S rRNA and ail genes of Y. enterocolitica according to Wannet et al. (2001). All suspected Y. pseudotuberculosis isolates were confirmed by PCR targeting a specific region of the inv of Y. pseudotuberculosis (Nakajima et al. 1992). All Y. enterocolitica isolates confirmed by PCR targeting the 16S rRNA region and inv-positive Y. pseudotuberculosis isolates were serotyped by slide agglutination using commercial antisera O:1, O:2, O:3, O;5, O:8, and O:9 for Y. enterocolitica and O:1-O:6 for Y. pseudotuberculosis (Denka Seiken). The presence of the virulence plasmid was studied by PCR targeting the virF gene on the virulence plasmid (pYV) (Nakajima et al. 1992, Joutsen et al. 2013). The virulence gene ail from six Y. enterocolitica biotype 1A isolates (four sucrose-positive and two sucrose-negative ones) and one Yersinia kristensenii isolate was confirmed by sequencing in both directions. Sequencing data were analyzed with BioNumerics (Applied Maths).
Statistical analysis
The statistical analysis was conducted with SPSS software version 24 (IBM). The differences in the yearly proportion of Yersinia spp. positive voles, mice, and shrews separately and in the isolation from oral or intestinal samples were analyzed with Pearson's chi-squared test.
Results
We investigated samples from 1840 wild small mammals for the presence of Yersinia spp., including voles (1171), mice (376), and shrews (293), trapped in 2001–2007 from 37 trapping locations in Finland (Table 1). The sample size in each trapping location in each year is presented in Table 2. Most of the material comes from 30 trapping locations along two transect lines crossing Southern Finland from the western coast to the eastern border (Fig. 1). The isolates were characterized, and the presence of virulence gene ail studied to assess the distribution of different Yersinia strains among wild small mammals captured at several locations over several years. From 1840 small mammals screened, a total of 641 Yersinia spp. isolates were obtained from 227 (12%) wild small mammals (Table 3). Yersinia-positive animals were trapped from 78% (29/37) of the trapping locations. Yersinia spp. were isolated more commonly (p < 0.001) from intestinal samples (60%, 407 isolates from 678 samples) than from oral samples (15%, 234 isolates from 1603 samples).
Y. enterocolitica was the most common species found in 142 (8%) animals (Table 3). Almost all (358/359) Y. enterocolitica isolates belonged to the nonpathogenic biotype 1A, and only one isolate from intestinal sample of a field vole belonged to bioserotype 2/O:9, which is associated with human and animal yersiniosis (Table 4). Some (14%; 50/359) Y. enterocolitica isolates were sucrose negative and, thus, first identified as Y. kristensenii using API 20E; however, they were confirmed to be Y. enterocolitica 1A after biotyping and PCR targeting the 16S rRNA region specific for Y. enterocolitica. Biotype 1A isolates were of serotypes O:2, O:2,3, O:3, O:3,9, O:5, O:8, or O:9. Using API 20E, 13 isolates from four shrews were identified as Y. pseudotuberculosis (Table 3). However, only eight of these isolates, obtained from intestinal samples of two common shrews were confirmed as Y. pseudotuberculosis after PCR targeting the species-specific region of the inv gene (Table 4). All Y. pseudotuberculosis isolates belonged to biotype 1 and serotype O:2.
16S rRNA sequence studied only in Y. enterocolitica.
ail sequence typically found only in Y. enterocolitica strains of biotypes 1B, 2–5.
inv sequence studied only in Y. pseudotuberculosis.
ND, not determined.
In total, 82 (23%) out of 358 Y. enterocolitica 1A isolates and 12 (6%) out of 213 Y. kristensenii isolates carried the chromosomal virulence gene ail. The ail-positive Y. enterocolitica 1A and Y. kristensenii isolates were isolated from 41 and four small mammals, respectively. Y. enterocolitica 1A carrying the ail gene was isolated in most of the species studied (Table 5). The house mouse and harvest mouse were the only species without any ail-positive Yersinia isolates, but their sample sizes were also the smallest. The ail-positive isolates were most frequently found in field voles (9%) and sibling voles (10%), especially in 2005, which was a cyclic peak year of voles. The proportion of Yersinia-positive voles was significantly higher (p < 0.001) during the years 2005 and 2007 compared to other years, and also to other peak year of 2002. There were no significant differences in the proportion of Yersinia spp. carrying animals between other years when voles were trapped. Y. kristensenii carrying the ail gene was isolated from three bank voles and one common shrew, which were all trapped in the same area in 2005. Small mammals carrying the ail-positive Yersinia were trapped in 12 different locations (32%) (Table 5). Field voles excreting ail-positive Yersinia were found in seven (28%) trapping locations (Table 5). The prevalence of ail-positive Yersinia in the animals varied from 1% to 14% at the 12 positive trapping locations (Fig. 2). Most of the ail-positive isolates (84%) were found in the intestines.
Only species with ail-positive Yersinia spp. isolations.
Percentage of ail-positive animals in a species.
Percentage of trapping locations with ail-positive animal species out of locations where species was captured.
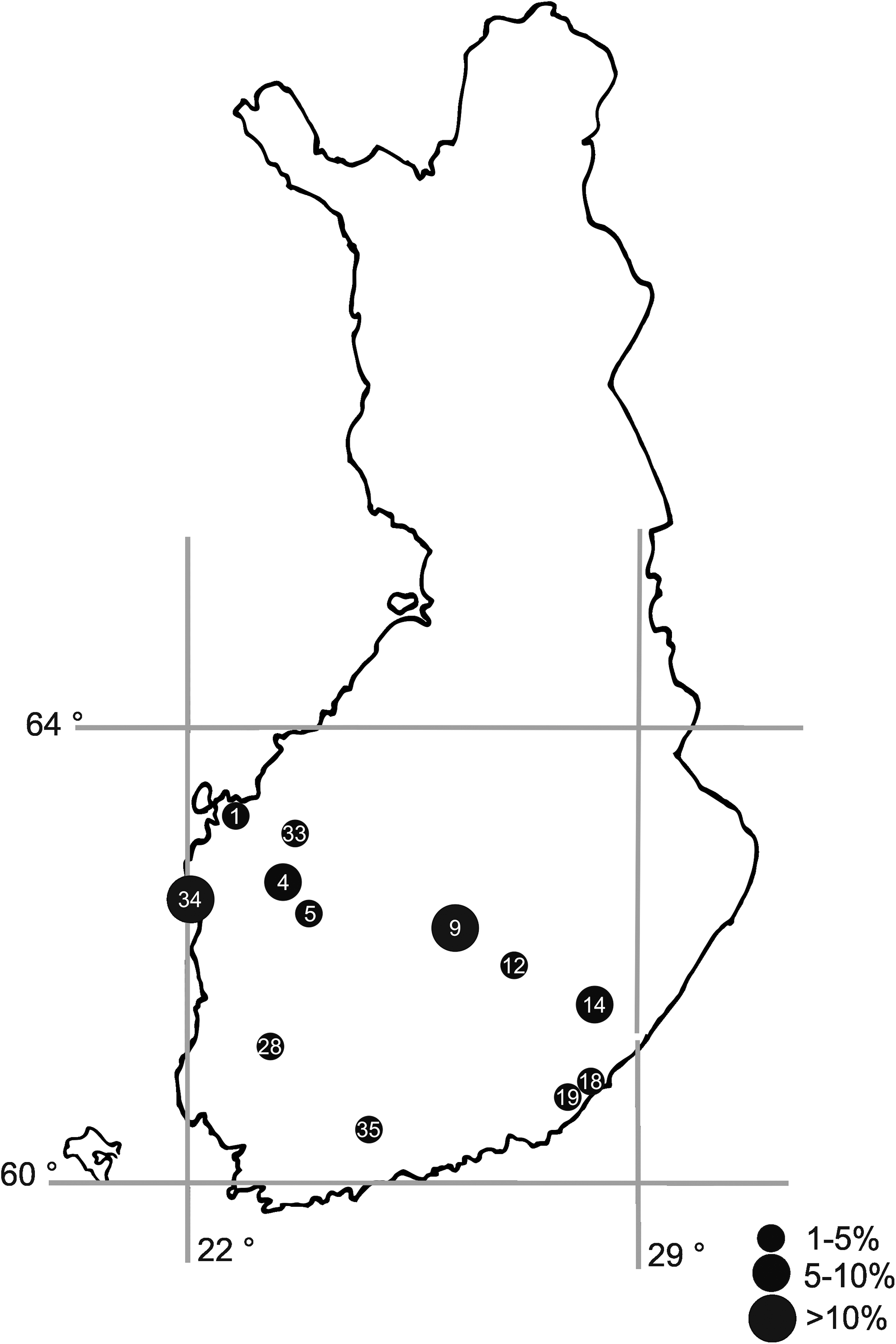
Trapping sites with ail-positive Yersinia spp. carrying animals in Finland. The number refers to trapping locations in the Table 2 and the size of the circle to the percentage of ail-positive Yersinia spp. carrying animals.
The partially sequenced ail-gene fragments (111-bp) from six Y. enterocolitica 1A isolates and one Y. kristensenii isolate were identical to each other and had 98.8% similarity with previously isolated ail gene from Y. enterocolitica biotype 1A (GenBank acc. no. FN812732.1).
Discussion
There are only few studies on the prevalence of Yersinia spp. in wild animals in Europe, including wild boars and deer but not wild small mammals (Fredriksson-Ahomaa et al. 2011, Joutsen et al. 2013, Bancerz-Kisiel et al. 2015). In our study, the prevalence of Yersinia spp. in Finnish wild small mammal populations from several locations across southern Finland was studied during a 7-year period. While most of the Yersinia isolates belonged to nonpathogenic species, especially Y. kristensenii or to nonpathogenic Y. enterocolitica of biotype 1A, pathogenic Y. pseudotuberculosis of bioserotype 1/O:2 and Y. enterocolitica of bioserotype 2/O:9, both of which carried the virulence plasmid (pYV), were also isolated. Y. pseudotuberculosis was only isolated from common shrews, suggesting that shrews may be carriers for this enteropathogenic species causing yersiniosis in both humans and animals. Multiple Y. pseudotuberculosis epidemics related to vegetables have occurred in Finland (Nuorti et al. 2004, Jalava et al. 2006, Kangas et al. 2008, Rimhanen-Finne et al. 2009). Wild small mammals with access to vegetables during harvest or storage were suggested to be the potential source of contamination in one of these Y. pseudotuberculosis-inflicted carrot epidemics (Jalava et al. 2006). Y. pseudotuberculosis was isolated from a pooled sample of common shrew intestines from one of these epidemic-related farms (Kangas et al. 2008). Long-term storage of carrots in cold temperatures favors the growth of enteropathogenic Yersinia spp. if the vegetables are contaminated by small mammals during the harvest or storage. The common shrew was the only species in our study known to excrete Y. pseudotuberculosis in its feces. Common shrew is, unlike rodent species in this study, an insectivore and eats a lot of worms (Churchfield 1990). Y. pseudotuberculosis thrives in cool soils and most probably worms continuously filtering soil accumulated bacteria, further contaminating shrews.
The only pathogenic Y. enterocolitica isolate carrying the virulence plasmid belonged to bioserotype 2/O:9 and was found in a field vole. This bioserotype is the second most common cause of human yersiniosis, but the reservoir remains unknown (EFSA 2007, Moriki et al. 2010). All these enteropathogenic Yersinia carrying the virulence plasmid were isolated from intestinal samples showing that wild small mammals sporadically excrete these pathogens in their feces. This may suggest that wild small mammals play a previously unknown role in the epidemiology of human yersiniosis by shedding pathogenic strains into the environment. In a previous study, it was shown that laboratory mice can develop a persistent but asymptomatic Y. pseudotuberculosis infection in the colon when infected with low doses of pathogen and shed the pathogen (Fahlgren et al. 2014). Wild small mammals carrying and possibly amplifying and excreting pathogenic Yersinia spp. may produce a risk if coming into contact, e.g., with noncarrier domestic animals, food storages, or irrigation water sources. It is very typical in the strongly seasonal Finnish climate that wild small rodents, and even shrews, invade human settlements in late autumn–early winter when the first frosts appear. For example, the epidemic peak of hantaviral disease caused by Puumala hantavirus takes place in October–January in Finland (Brummer-Korvenkontio et al. 1999, Kallio et al. 2009). However, further studies are needed to clarify the persistence of Yersinia infection and shedding of enteropathogenic Yersinia spp. in small mammals.
Yersinia was an occasional finding in all wild small mammals, and Y. enterocolitica biotype 1A and Y. kristensenii strains carrying the ail gene were isolated especially from voles. Bank and field voles, as well as common shrews, are the most common small mammal species in Finland and they were the most common species trapped during our study. Voles exhibit strong multiannual density fluctuations, often called cycles, in Finland (Korpela et al. 2013). Therefore, the annual numbers of trapped animals varied quite widely. Consequently the numbers of positive animals varied multiannually, particularly because the sample sizes were low in cyclic decline years (2003, 2006). The proportions of animals carrying ail-positive Yersinia spp. were higher during the peak years, while during years of lowest densities no ail-positive isolates were detected. However, with the prevalence of peak year ail positives, it was not very probable to find positives in low-density years. Furthermore, the prevalence of animals excreting ail-positive Yersinia isolates was high in some trapping areas (locations 4, 9, 24, and 34) suggesting that horizontal transmission of the ail gene between the pathogenic and nonpathogenic Y. enterocolitica and between Y. enterocolitica and Y. kristensenii isolates may have occurred in wildlife in these areas. This will be studied further. The ail-positive small mammals were found in the study localities across Finland, suggesting that there is no geographic clustering of them.
Pathogenic Y. enterocolitica strains attach and invade the host tissue cells with the help of ail (Pierson and Falkow 1993). The ail-positive Y. enterocolitica 1A and Y. kristensenii isolates are possibly able to invade the intestinal cells of small mammals and persist there despite a lack of the virulence plasmid needed for full virulence and infection. However, further studies are needed to evaluate the full virulence of these ail-positive isolates, including the functionality of the ail gene.
The ail gene is typically associated only with pathogenic Y. enterocolitica strains (Miller et al. 1989). The ail-positive Y. enterocolitica has sporadically been found in wild animals, food, and humans with diarrhea (Sihvonen et al. 2009, Cheyne et al. 2010, Kraushaar et al. 2011, Liang et al. 2014, Bancerz-Kisiel et al. 2015). PCR detection of ail is commonly used in the detection of pathogenic Y. enterocolitica (Fredriksson-Ahomaa and Korkeala 2003, Lambertz et al. 2008), but the presence of ail also in nonpathogenic Y. enterocolitica and other Yersinia spp. questions the usefulness of this gene alone in PCR detection.
The identification of Yersinia species was challenging during our study. Differentiating between sucrose-negative Y. enterocolitica and Y. kristensenii isolates was impossible using only API 20E, and PCR targeting the 16S rRNA was needed for correct identification. Identification of Y. pseudotuberculosis was also impossible with API 20E, and thus, confirmation was performed using a PCR targeting the species-specific region of the inv gene in Y. pseudotuberculosis. The identification of environmental presumptive Y. pseudotuberculosis strains has been shown to be potentially incorrect when using biochemical tests (Niskanen et al. 2009). Typically, correctly identified Y. pseudotuberculosis isolates can be serotyped and they carry the virulence plasmid. All Y. pseudotuberculosis isolates in our study were of bioserotype 1/O:2, which is one of the bioserotypes found in wild birds and boars in Europe (Niskanen et al. 2003, Fredriksson-Ahomaa et al. 2011). The Y. pseudotuberculosis isolates also carried the virulence plasmid indicating full pathogenicity of these isolates.
Conclusions
Y. pseudotuberculosis of bioserotype 1/O:2 was isolated from two common species of shrew and Y. enterocolitica 2/O:9 from one species of field vole, indicating that wild small mammals could carry enteropathogenic Yersinia spp. and thus could also act as possible contamination sources of vegetables. Nonpathogenic Y. enterocolitica of biotype 1A frequently carried the ail gene typically found only in pathogenic Y. enterocolitica of biotypes 1B and 2–5, hampering the identification of pathogenic Y. enterocolitica isolates. Surprisingly, the ail gene was also detected sporadically in nonpathogenic Yersinia species Y. kristensenii.
Footnotes
Acknowledgments
This article was written in memory of the late DVM Susanna Kangas, who isolated the Yersinia isolates studied. The study was funded by the Finnish Ministry of Agriculture and Forestry (4637/501/2004). The study was also partially funded by EU 6th Framework Programme (GOCE-2003-010284 EDEN,
Author Disclosure Statement
No competing financial interests exist.