
Abstract
Bacteria in the genus Anaplasma are responsible for diseases in animals and humans. Studies carried out in Brazil have demonstrated that Brazilian deer are able to act as hosts of agents in the family Anaplasmataceae and are possibly potential reservoirs of these pathogens. Molecular and phylogenetic studies have been carried out on samples of two gray brocket specimens (Mazama gouazoubira) from the city of Guarapuava, state of Paraná, Brazil, for the detection of Anaplasma sp. in these animals. Partial nucleotide sequences of the genes 16S rRNA and groESL were used for phylogenetic analyses and compared with other 13 and 17 partial sequences of the respective genes obtained in GenBank. These assessments showed topological incongruence among the trees generated in the phylogenetic analyses. Phylogenetic analysis based on the gene 16S rRNA of the genotypes amplified in the samples of this study was similar to those of A. bovis detected in dogs and wild deer in Japan, whereas studies carried out on gene groESL indicated proximity with sequences of Anaplasma sp. that were also isolated in deer in Japan and allocated in the same clade of partial sequences of A. phagocytophilum. As the 16S rRNA gene is highly conserved, with few polymorphic positions, it may show low reliability for studies on phylogenetic positioning. The present study detected an Anaplasma sp. genotype in two specimens of M. gouazoubira in southern Brazil, which may mean that this agent possibly circulates in deer populations, and demonstrated the need for studies related to the possible role of deer in enzootic cycles of Anaplasmataceae in Brazil.
Introduction
T
Important information on disease epidemiology may be obtained from studies on the identification of infectious agents in wild animals, especially deer, once these animals are growingly closer to populations of domestic animals (Silveira et al. 2011).
The gray brocket (Mazama gouazoubira) is a small species of deer that is found all over Brazil, and may be seen from Mexico to the north of Argentina. Besides being affected by hunting and habitat destruction, the fact that they may live in areas that are close to populations of domestic animals may be a threat to these animals due to the risk of disease transmission (Duarte et al. 2012).
Studies carried out in Brazil have demonstrated that Brazilian deer may act as hosts of agents in the family Anaplasmataceae, and may be potential reservoirs of these pathogens (Machado et al. 2006, Picoloto et al. 2010, Sacchi et al. 2012, Silveira et al. 2012, 2013, 2014).
Environmental changes caused by natural phenomena or human actions have changed the context and ecological balance in the reproduction, development, and disease transmission dynamics of parasites and vectors. The constant changes in the ecosystems have affected the distribution and behavior of parasites, as well as that of vectors and hosts (Patz et al. 2000).
The objective of the present study was the molecular characterization of the occurrence of Anaplasma spp. in two gray brocket specimens (M. gouazoubira) seen in the Attention Service to Wild Animals (SAAS) of the city of Guarapuava, state of Paraná, Brazil.
Material and Methods
Specimens sampled
From July 2013 to November 2014, two adult free-living specimens of Mazama gouazoubira, called Mazama gouazoubira one (MG1) e Mazama gouazoubira two (MG2), were seen at the Attention Service to Wild Animals (SAAS) at Universidade Estadual do Centro-Oeste (UNICENTRO), located in the city of Guarapuava, in the center-west region of Paraná, Brazil. The animals were attended in different periods, but both because of trampling on highways. The region of Guarapuava (25°23′24″S; 51°27′43″W) is located at the Midwest area of Paraná State, with a representative vegetation of Araucaria moist forest and rural activity mainly with extensive livestock production.
Clinical examination showed the presence of ticks, which were collected and stored in ethanol 70% (Merck®) to be identified later on using the dichotomous keys of Aragão (1936) and Cooley (1946). In addition, blood samples were collected in tubes with EDTA 5% and stored at −20°C. These samples were sent to the Laboratório de Imunoparasitologia at Universidade Estadual de Paulista “Júlio de Mesquita Filho,” in the city of Jaboticabal, state of São Paulo, Brazil, for DNA extraction for PCR and sequencing.
DNA extraction and cPCR for Anaplasma spp. based on genes 16S rRNA, groESL, MSP1-β, MSP4, and MSP5
DNA extraction was carried out in 200-μL aliquots of blood from the animals using the DNeasy® Blood & Tissue Kit (Qiagen®), according to the recommendations of the manufacturer. DNA aliquots were placed in microtubes and stored at −20°C for PCR. DNA concentration was measured in a NanoDrop 2000c spectrophotometer (Thermo Scientific, San Jose, CA).
PCR reactions for Anaplasmataceae agents based on gene 16S rRNA were carried out in 5 μL of DNA from the samples. Starter oligonucleotides and thermal sequences used in the PCR, as well as thermal sequences used in each agent, are shown in Table 1.
The amplified products were analyzed by horizontal electrophoresis in 1.0% agarose gel stained with Ethidium Bromide (0.5 μL/mL) in TEB buffer pH 8.0 (44.58 M Tris-base; 0.44 M boric acid; and 12.49 mM EDTA). Electrophoresis was carried out at 90 V/150 mA for 60 min. A molecular weight marker with 100-bp was used in the analysis of the amplified products (Life Technologies®), and the results were read and analyzed in an UV transilluminator (ChemiDoc™ MP Imaging System; Bio-Rad®).
Purification and sequencing of the amplimeres and phylogenetic analysis
PCR products were purified using the “Silica Bead DNA Gel Extraction Kit” (Fermentas, São Paulo, Brazil), according to the recommendations of the manufacturer. Quantification of the total genome material of the purified product was carried out in a NanoDrop instrument (Thermo Scientific®) by measuring the absorbency of each sample.
Amplified and purified DNA fragments obtained in positive samples were subjected to sequence confirmation in an automatic sequencer (ABI PRISM 310 Genetic Analyser-Aplicada Biosystems/Perkin Elmer). Consensus sequences were obtained by sense and antisense analysis using the CAP3 software (
Phylogenetic analyses were carried out based on Bayesian inference (BI), for partial sequences of the gene groESL, and on maximum-likelihood estimation for both amplified genes. BI was carried out with MrBayes 3.1.2 (Ronquist and Huelsenbeck 2003) on CIPRES science gateway (Miller et al. 2010), using the best models selected in the MEGA-4 software 4.0 (Kumar et al. 2004), based on the corrected Akaike Information Criterion (AICc). Markov chain Monte Carlo (MCMC) estimations were carried out for 109 generations, with sampling frequency at every 100th generation and burn-in of 25%. Maximum likelihood trees were generated using the RAxML Black Box cluster (Stamatakis et al. 2008) in the CIPRES Science Gateway platform (Miller et al. 2010). The proportion of invariant sites was estimated in the evolutionary model GTRGAMMA+I. Edition of phylogenetic trees, as well as rooting (through out-group), was carried out in TreeGraph 2.0.56-381 beta software (Stöver and Muller 2010).
Results
Diagnosis of Anaplasma spp. in blood samples of wild deer
Nested-PCR reactions based on the genes 16S rRNA and groESL showed positive results for Anaplasma spp. in the samples collected from the two deer analyzed in the present study (MG1 and MG2). It was observed that only one of the samples positive in nested-PCR based on 16S rRNA was also positive in the nested-PCR assay based on gene groESL.
Both animals did not show any clinical signs. During the inspection of the animals, deer MG1 was infested by ticks that were identified as adult specimens of Rhipicephalus (Boophilus) microplus, as they showed hexagonal basis capituli, palps with edges shorter than the hipostome, and no festoons (Aragão 1936, Cooley 1946). Hemaphysalis juxtakochi nymphs and adults were also identified by morphology similar to the description by Martins (2007), with rectangular basis capituli, second segment of the palp with a salient projection, absence of eyes, and presence of spurs in the trochanters of legs number I to IV. A female Amblyomma parvum, identified by the small size, long rostrum, presence of eyes, and short scutum with no spots (Aragão 1936), was also identified. Deer MG2 was not infested by ectoparasites at the moment of sample collection.
Phylogenetic analyses based on gene 16s rRNA
BLAST nucleotide sequence analysis (932-bp) based on a sample of MG1 blood showed 98% identity for Anaplasma bovis genotypes detected in Brazil, China, and South Korea (GenBank accession numbers: JF952893.1, KJ639885.1, and GU556627.1, respectively). For the sequence obtained with the MG2 sample, BLAST showed 100% identity with DNA fragments of the gene 16S rRNA of A. bovis detected in Brazil, Japan, and China (accession numbers: JF922893.1, AB983376.1, and KJ659043.1).
Phylogenetic inference led to a cladogram that placed the genotypes detected in the deer specimens (from the Guarapuava region, Brazil, MG1** and MG2**) close to the A. bovis ones found in dogs (LC012812.1) and in wild deer in Japan (AB211163.1). A partial sequence of the gene 16S rRNA of Neorickettsia sennetsu (M73219.1) was used as out-group (Fig. 1).
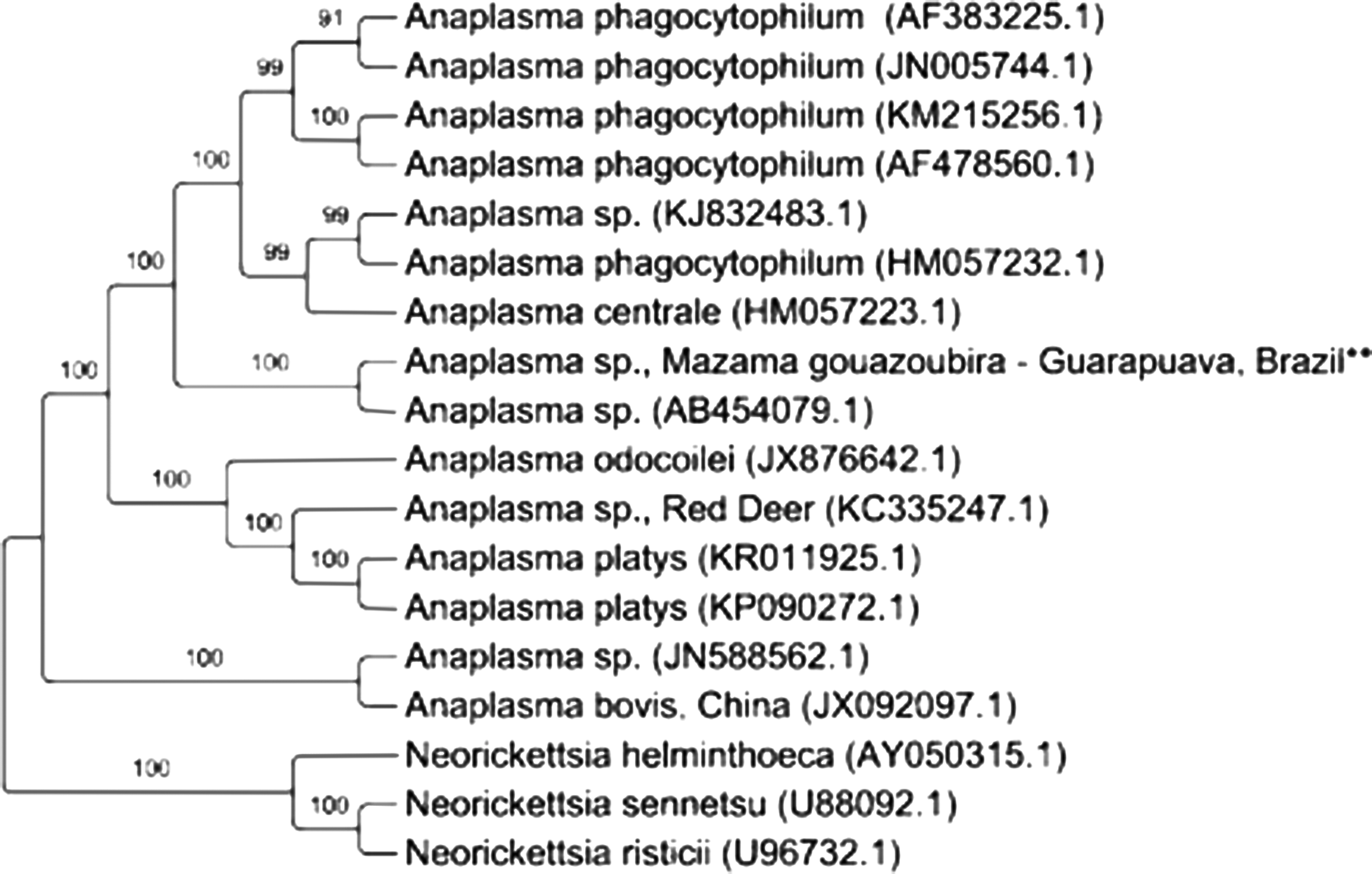
Phylogenetic positioning of Anaplasma sp. 16S rRNA gene sequences. Tree drawn using Bayesian inference in the GTR + I + G model and corrected Akaike Information Criterion (AICc). Markov chain Monte Carlo (MCMC) estimations were carried out for 109 generations, with sampling frequency at every 100th generation and burn-in of 25%. The numbers in the tree indicate the probability values for clade support above 50 for the branches. **Sequences of the present study.
Phylogenetic analyses based on gene groESL
BLAST analysis of a fragment of the gene groESL from Anaplasma sp. with 1297-bp that was amplified from the blood sample of deer MG1 showed 80% identity with Anaplasma sp. detected in wild deer in Japan (AB454079.1). Phylogenetic analyses based on BI estimation (Fig. 1) positioned the two sequences close to each other. It was observed that the sequence detected in the present study was not positioned close to A. bovis used in the drawing of the cladogram (JX092097.1). Partial sequences of the gene groESL of Neorickettsia helminthoeca (AY050315.1), N. sennetsu (U88092.1), and Neorickettsia risticii (U96732.1) were used as out-groups.
Discussion
The occurrence of an Anaplasma sp. genotype in two free-living M. gouazoubira deer was reported in the present study. The same species was reported as a host of other Anaplasmataceae agents, such as those phylogenetically related to A. platys and A. phagocytophilum (Silveira et al. 2012). Recently, Silveira et al. (2014) reported the detection of Anaplasma marginale and A. phagocytophilum in M. gouazoubira deer in the Brazilian states of Mato Grosso do Sul and Minas Gerais, evidencing the role of these wild ruminants as hosts of these agents.
Natural infection with Anaplasmataceae agents in dromedary camels has been reported by Bastos et al. (2015), using a combined 16S rRNA-groESL analysis, which demonstrates the occurrence of a novel Anaplasma lineage in these animals. In the present study, the found Anaplasma 16S rRNA and groESL partial sequences were used in phylogenetic analyses to compare with other 13 and 17 partial sequences of the respective genes retrieved from the GenBank database. These assessments showed topological incongruence among the trees generated in the phylogenetic analyses performed with partial sequences of the 16S rRNA and groESL genes.
In the present study, Anaplasma spp. genotypes showed phylogenetic proximity with A. bovis (LC012812 and AB212263), based on maximum likelihood phylogenetic inference (Fig. 1).
Cladograms drawn based on a groESL 297-bp gene fragment obtained from MG1-deer blood samples were closely related to a partial sequence of Anaplasma sp. (AB454079.1). These two sequences were allocated in a clade in which partial sequences of the A. phagocytophilum genotypes (AF383225, JN005744, KM215256, AF478560, and HM057232) were grouped. Phylogenetic trees drawn from these two analyses presented separation clades that were better supported by the maximum likelihood estimation based on gene16S rRNA and topology similar to that of the terminal nodes (Fig. 1).
According to Doan et al. (2013), the 16S rRNA gene is a molecular tool that is sensitive for the detection of circulating Anaplasma spp. genotypes. However, Paulauskasa et al. (2012) stated that the use of msp4 gene shows a higher reliability in terms of phylogenetic positioning, as gene 16S rRNA is highly conserved, with few polymorphic positions, a finding that is corroborated by the results of the present study. In a study performed to analyze the genetic variability of A. phagocytophilum genotypes detected in Ixodes persulcatus ticks and small mammals, differentiation of sequences obtained with the amplification of the gene groESL was higher than that observed with the amplification of gene 16S rRNA, when the method by Neighbor Joining was used, which demonstrated a distinction between A. phagocytophilum genotypes circulating in Russia and other regions (Rar et al. 2011).
It is also known that Anaplasma spp. genotypes phylogenetically related to A. phagocytophilum have already been detected in wild carnivores (André et al. 2012), wild birds (Machado et al. 2012), domestic cats (André et al. 2014), and gray brockets (M. gouazoubira) in Brazil (Silveira et al. 2012). These findings indicate a possible circulation of genotypes similar to A. phagocytophilum in wild animals in Brazil, whose zoonotic potential is still unknown. Genotypes associated with A. phagocytophilum have been detected in other regions of the world, such as in specimens of wild deer in oriental Slovakia (Víchová et al. 2014) and small wild mammals in Taiwan (Masuzawa et al. 2014).
According to Zobba et al. (2014), ruminants play an important role in the epidemiology of bacteria in the genus Anaplasma. The diversity of genotypes circulating in these groups of mammals is more complex than it was considered before. For example, Anaplasma spp. genotypes phylogenetically associated with A. phagocytophilum, A. marginale, A. ovis, A. bovis, and A. platys were detected in domestic ruminants in the Mediterranean region. In addition, Anaplasma spp. genotypes associated with A. phagocytophilum, Anaplasma sp., A. platys, and Ehrlichia ruminantium have been described in sheep in Senegal (Djiba et al. 2013). Recently, a high diversity of Anaplasma genotypes has been described in buffaloes (Syncerus caffer) in Mozambique (Machado et al. 2016).
However, the extinction risk for M. gouazoubira is considered least concern (LC), since this animal species is widely distributed in the state of Paraná, supporting its role as a disperser of vector and pathogens (Duarte et al. 2012).
The occurrence of the tick H. juxtakochi was described in other regions of the state of Paraná in animal species in the genera Tapirus and Mazama (Barros and Baggio 1992), as well as in birds in the Curitiba metropolitan area (Arzua et al. 2005). In the center-west region of the state of Paraná, this tick was also found in the city of Prudentópolis (Arzua et al. 2005), 50 km away from Guarapuava, also as a parasite in M. gouazoubira, which may indicate that this species of tick is common in the region.
In a study carried out by Barré et al. (2001), it was shown that there are no significant differences in the infection patterns of R. microplus in cattle or deer herds, suggesting that the presence of a primary host does not affect the infestation of alternative hosts by this tick in the same environment. Reports on R. microplus in different species of Brazilian deer have been described in the literature (Serra-Freire et al. 1996, Figueiredo et al. 1999, Bechara et al. 2000, Labruna et al. 2002, Szabó et al. 2003, 2007, Silveira et al. 2011), including in Paraná (Arzua et al. 2005).
Since Guarapuava surrounded regions are characterized by rural areas with mainly extensive livestock production, the occurrence of cross-contamination between wild and domestic animals is a potential problem for economic and conservative goals. Studies suggest that although deer do not act as preferential hosts of R. microplus, the proximity of these animals with cattle herds make infestation of wild deer easier, and may represent a potential threat that demands more studies to support correct deer management in the future (Szabó et al. 2003).
Herein, conventional PCR assays for MSP5 (Torioni de Echaide et al. 1998, Singh et al. 2012) and MSP4 (De La Fuente et al. 2005, Bown et al. 2007) and qPCR for MSP1β (Carelli et al. 2007) were run in an attempt to perform a molecular characterization of positive samples using other genes. Unfortunately, deer samples were negative to these additional target genes. This pattern of positivity to Anaplasmataceae 16S rRNA-based PCR assays and negativity to other target genes has already been described by previous works that aimed at detecting Anaplasmataceae agents in domestic and wild animals in Brazil (André et al. 2012, 2014, 2015, Sacchi et al. 2012) and Africa (Machado et al. 2016). We believe that new genotypes of Ehrlichia and Anaplasma circulate in wild and domestic animals in Brazil. The preestablished PCR protocols targeting different genes other than 16S rRNA have been showing low accuracy to catch Anaplasmataceae new genotypes in Brazil.
Conclusion
Epidemiological studies are still lacking to provide knowledge on the role of wild animals in the dissemination of pathogens infectious to men and domestic animals in Paraná state, Brazil. Therefore, the present study demonstrated the detection of an Anaplasma sp. genotype in two Mazama gouazoubira specimens in southern Brazil based on molecular techniques and phylogenetic analyses. It may be concluded, based on these data, on the possible circulation of this agent among populations of wild deer, a fact that demonstrates the need to carry out studies on the possible role of deer in enzootic cycles of Anaplasmataceae agents in Brazil to elucidate the real risks for deer's conservation medicine and transmission of pathogens to livestock.
Footnotes
Author Disclosure Statement
No competing financial interests exist.