
Abstract
Introduction:
Leptospirosis is caused by Leptospira spp. and is considered the most widespread zoonotic disease worldwide. It mimics nephropathia epidemica in humans, a disease mainly caused by Puumala hantavirus (PUUV). Small mammals are reservoirs for Leptospira spp. and PUUV. Seewis virus (SWSV) is a shrew-borne hantavirus with unknown pathogenicity. The objective of this study was to estimate the prevalence for Leptospira spp. and the frequency of Leptospira-hantavirus co-infections in small mammals collected at locations with high and low incidences in humans.
Materials and Methods:
In 2012 and 2013, 736 small mammals belonging to seven species (Apodemus flavicollis, Microtus agrestis, Microtus arvalis, Myodes glareolus, Sorex araneus, S. coronatus, and S. minutus) were collected at four high incidence sites (H1-H4) and four low (L1-L4) incidence sites for PUUV infection in humans. Kidney-derived DNA samples were tested for Leptospira spp. by real-time PCR targeting the lipl 32 gene and further analyzed by duplex PCR targeting the flaB and the secY genes. For the detection of Seewis virus, lung-derived DNA was tested via RT-PCR targeting the nucleocapsid gene.
Results:
Altogether, 42 of the 736 small mammals including 27 of 660 bank voles and 11 of 66 shrews, were positive for Leptospira spp., while Sorex spp. (14.7%) showed significantly higher prevalences compared to bank voles (4.1%). Detected Leptospira spp. were pathogenic species other than L. kirschneri. Significantly more Leptospira-positive bank voles were found at H sites than at L sites. Altogether 22.2% of positive bank voles were infected with PUUV. Double infection of PUUV and Leptospira spp. occurrence in bank voles is 1.86 times (OR = 1.86; 95% CI: 0.72–4.73) more likely than infections with each pathogen separately.
Discussion:
Leptospira- positive bank voles are focally positively associated with PUUV infection in bank voles and with human hantavirus cases. It should be considered that shrews may serve as Leptospira spp. reservoirs.
Introduction
L
The objectives of this study: (i) Detection and comparison of prevalences of pathogenic Leptospira spp. in small mammal species from sites with high and low hantavirus incidences in humans, (ii) Quantification of hantavirus (PUUV and SWSV)-Leptospira spp. co-infections in small mammals.
Materials and Methods
Small mammal samples
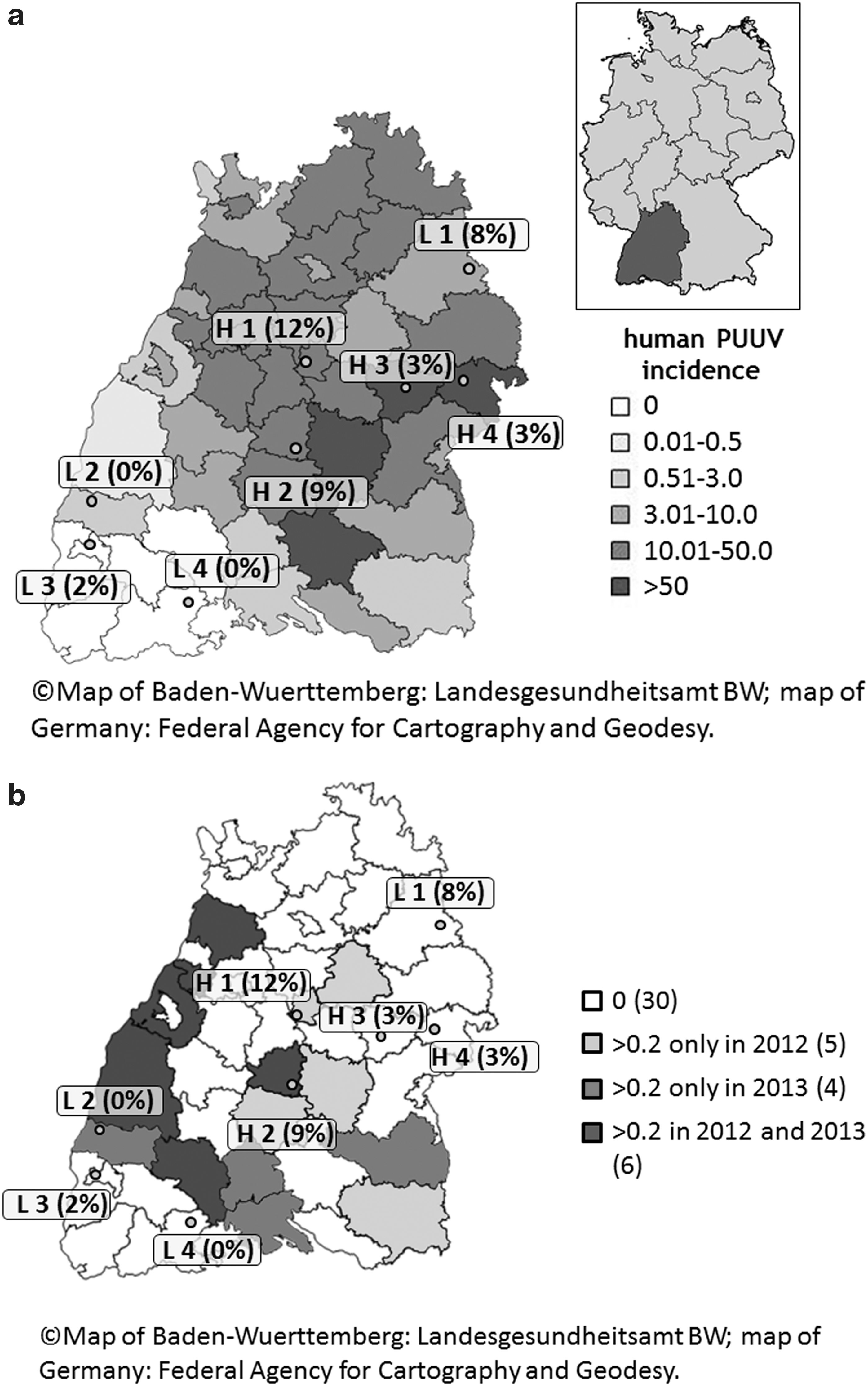
In 2012 (between June and November) and 2013 (between March and November), small mammals were collected at four sites with high incidence of notified human hantavirus cases (sites H1-H4) and four sites with low notified incidence (sites L1-L4) in Baden-Wuerttemberg, Germany (Drewes et al. in press; Fig. 1a and Table 1). In total 736 small mammals of seven species were available for the present study (Myodes glareolus (n = 660), Sorex coronatus (n = 44), Sorex araneus (n = 22), Apodemus flavicollis (n = 4), Microtus agrestis (n = 3), Sorex minutus (n = 2), and Microtus arvalis (n = 1)). In 2012, 526 small mammals (499 Myodes glareolus, 13 S. coronatus, 7 S. araneus, 3 Microtus agrestis, 3 A. flavicollis, and 1 Microtus arvalis) were trapped with a range of 38–122 animals per site. In 2013, 210 small mammals (161 Myodes glareolus, 31 S. coronatus, 15 S. araneus, 2 S. minutus, and 1 A. flavicollis) were captured. Further information on trapping methods were published before (Drewes et al. in press).
Microtus arvalis (n = 1) and Microtus agrestis (n = 3).
Microtus arvalis.
DNA and RNA extraction
For DNA extraction, 600–800 μL phosphate-buffered saline (PBS, pH: 7.2) was added to 0.01–0.02 g of each small mammals' kidney tissue, depending on the initial kidney sample weight. Subsequently, 0.6 g of sterile ceramic beads (PeqLab Biotechnologie GmbH, Erlangen, Germany) (diameter sized 1.4 mm) were added to each sample. Samples were homogenized for 3 × 15 s at 100 Hz by the Precellys® 24 Tissue Homogenizer (Bertin Technologies, Montigny-le-Bretonneux, France). Afterward, DNA was extracted with the QIAamp DNA Mini Kit (Qiagen, Hilden, Germany) as recommended by the manufacturer. Quantity and quality of the extracted DNA samples were determined with a spectrophotometer (Peqlab Biotechnologie GmbH, NanoDrop 2000c).
For RT-PCR detection of SWSV in shrews, RNA was isolated from lung tissue by QIAzol® Lysis Reagent (Qiagen) treatment as described by Schmidt et al. (2016).
Samples were tested for PUUV by PCR and ELISA in a previous study (Drewes et al. in press).
Altogether, 90of 660 M. glareolus (13.64%; 95% CI: 11.2–16.5) were previously detected positive for PUUV. In 2012, 85 of 499 M. glareolus (17.0%; 95% CI: 14.0–20.6) were positive for PUUV and in 2013, five out of 161 M. glareolus (3.1%; 95% CI: 1.1–7.3) were tested positive (Drewes et al. in press).
PCR methods for the detection of Leptospira spp.
A real-time PCR targeting the partial lipl 32 gene for pathogenic Leptospira spp. was performed with the primers LipL32-45F (5′-AAG CAT TACCGC TTG TGG TG-3′) and LipL32-286R (5′-GAA CTC CCA TTT CAG CGA TT-3′) following a modified protocol using the Mx3000P QPCR System (Agilent Technologies, Santa Clara, CA) (Stoddard et al. 2009, Woll et al. 2012). We modified the PCR as previously described by Woll et al. (2012) by using a different probe (Univ 18 s-p: 5′ YAK-CGCGGTTCTATTTTGTTGGT BBQ-3′). This PCR method detects following pathogenic Leptospira spp.: L. interrogans, L. kirschneri, L. weilii, L. noguchii, L. borgpetersenii, L. santarosai, L. alstoni, and L. alexanderi. Its limit of detection was 20 genomic equivalents/reaction with a 95% cutoff value. To distinguish L. kirschneri from other pathogenic Leptospira spp., samples were further investigated by duplex PCR using primers G1 (5′-CTG AAT CGC TGT ATA AAA GT-3′), G2 (5′-GGA AAA CAA ATG GTC GGA AG-3′), B64 I (5′-ACT AAC TGA GAA ACT TCT AC-3′), and B64 II (5′-TCC TTA AGT CGA ACC TAT GA-3′) (Gravekamp et al. 1993). The duplex PCR is targeting the partial flagellin-encoding flaB gene (563 base pairs, bp), which exclusively amplifies in L. kirschneri and the preprotein translocase-encoding secY gene (285 bp), which amplifies in several different pathogenic Leptospira spp. such as L. interrogans, L. weilii, L. noguchii, L. borgpetersenii, L. santarosai, and L. meyeri (Gravekamp et al. 1993). We used this PCR method to distinguish L. kirschneri from other pathogenic Leptospira species. Due to low amounts of Leptospira DNA detected by real-time PCR, only 18 of 42 samples yielded a result by duplex PCR.
RT-PCR method for the detection of SWSV
Serological and RT-PCR analyses for PUUV and TULV in bank voles and in other rodents have been described previously (Drewes et al. in press). RT-PCR detection of SWSV in shrews and sequencing of amplification products followed a previously described standard protocol with slight modifications (Schlegel et al. 2012) using SuperScript®III One-Step RT-PCR kit with Platinum®Taq (Invitrogen, Carlsbad, CA) and the BigDye® Terminator version 1.1 Cycle Sequencing-Kit (Applied Biosystems, Darmstadt, Germany). For RT-PCR novel S segment-specific primers SWSV_S_400_fw 5′-ATC TTG CTY AAR GGA CTC TAC ATG CT and SWSV_S_ 890 rev 5′-TCA GGY ACW GAC TGG ACA GCA GG were used and the annealing temperature was adjusted to 56°C. The obtained SWSV sequence was deposited in GenBank under Acc. No. KX447717.
Statistical analysis
Confidence intervals (95% CI) for prevalences of Leptospira and hantavirus in small mammals were determined by the Clopper and Pearson method using the Graph Pad Software (Graph Pad Software, Inc., San Diego, CA). Fisher's exact test was used with a type I error α of 0.05 to test the independence of compared prevalences for Leptospira spp. (Graph Pad Software, Inc.). The odds ratio (OR) with 95% CI were computed to measure the association between Leptospira spp. infection and PUUV infection in bank voles.
Results
PCR analysis for pathogenic Leptospira spp.
In total 42 of 736 small mammal DNA samples were tested positive for Leptospira by real-time PCR (5.7%; 95% CI: 4.2–7.6; Table 1). Leptospira spp. was found in all small mammal species, except S. minutus and Microtus agrestis. The average prevalences in 2012 (5.7%; 95% CI: 4.0–8.0) and 2013 (5.7%; 95% CI: 3.2–9.8) were identical (p = 0.13). Out of 660 bank voles, 27 were positive for Leptospira spp. (4.1%; 95% CI: 2.8–5.9). The prevalence in shrews (14.7% 95% CI: 9.4–27.6) was significantly higher than in voles (Myodes glareolus and Microtus spp.) (p = 0.001). The prevalence in the 44 crowned shrews was (18.2%; 95% CI: 9.3–32.2) and in the 22 common shrews it was 13.6% (95% CI: 3.9–34.2). The two pygmy shrews were Leptospira negative (0%; 95% CI: 0–71.0). Moreover, two out of four yellow-necked mice and likewise two out of four individuals belonging to the genus Microtus spp. were positive for Leptospira spp.(50%; 95% CI: 15–85). Altogether 18 of 42 samples were further tested by duplex PCR and identified to be pathogenic Leptospira spp. other than L. kirschneri.
Positive small mammal DNA samples were found at six (sites H1-4 and sites L1, L3) of the examined eight study sites (Fig. 1a, b). The average prevalence was significantly lower at the L sites (2.7%; 95% CI: 1.3–5.4); than at the H sites (6.7%; 95% CI: 4.3–10.3) (p = 0.03). The prevalences of H and L sites did not differ significantly between the years 2012 and 2013 (p = 0.13). Prevalences per site and small mammal species are shown in Table 1.
Analysis of Leptospira–hantavirus co-infections
Double infections of PUUV and Leptospira spp. (22.2%; 95% CI: 10.3–41.1) occurred in six out of 27 Leptospira positive bank voles. Double infection of PUUV and Leptospira spp. occurrence is 1.86 times (OR = 1.86; 95% CI: 0.73–4.7) more likely than separate infections either with Leptospira spp. or PUUV in bank voles. All co-infected bank voles originated from H sites in 2012. Altogether, 50% of Leptospira-positive bank voles from site H1 were co-infected with PUUV (95% CI: 23.7–76.3). Leptospira co-infections were detected in PUUV-ELISA-positive and in PUUV-RT-PCR-positive bank vole samples. Hantavirus-specific maternal antibodies did not impact on results as all co-infected bank voles were adults according to their body weight (Drewes et al. in press).
In the present study, one out of 22 S. araneus (4.6%; 95% CI: 0–23.5) was detected to be SWSV-RT-PCR positive, whereas the two pygmy shrews (S. minutus) (95% CI: 0–71.0) and the 44 crowned shrews (S. coronatus) (95% CI: 0–9.6) were negative. The SWSV-infected common shrew was not co-infected with Leptospira spp.
Discussion and Conclusions
This study demonstrated an average Leptospira prevalence of 5.7% for 736 small mammals, with positive bank voles (4.1%), shrews (14.7%), Microtus spp. (50%), and yellow-necked mice (50%). The average prevalence observed here (0–50%) is in a range of prevalences previously obtained from small mammals in Germany (Mayer-Scholl et al. 2014, Heuser et al. 2016, Obiegala et al. 2016). Leptospira spp. were previously also detected in a similar wide prevalence range (2.9–71.4%) in small mammal species including Apodemus spp., Microtus arvalis, Myodes glareolus, Mus musculus, Rattus norvegicus, and Sorex spp. from other European countries such as Switzerland, The Netherlands, Croatia, Denmark, and Austria (Hartskeerl et al. 1996, Adler et al. 2002, Turk et al. 2003, Schmidt et al. 2014, Heuser et al. 2016). Varying prevalences were also detected in Rattus norvegicus, which were collected in The Netherlands, Austria, Hungary, and Germany where human cases were reported (Roczek et al. 2008, Goris et al. 2013, Heuser et al. 2016). In a previous study from Germany, moderate Leptospira spp. prevalences of 12–14% were detected in Microtus arvalis and Apodemus agrarius (Mayer-Scholl et al. 2014). While a previous study from Germany reported the absence of Leptospira spp. in shrews belonging to the family Soricidae (Crocidura russula n = 4; S. araneus n = 7; S. coronatus n = 1) (Obiegala et al. 2016), the prevalence in Sorex spp. obtained in the present study was 14.71%. Other studies from Germany and Switzerland reported similar prevalences (11–20%) in shrews of the family Soricidae (Adler et al. 2002, Mayer-Scholl et al. 2014). Though voles (family Cricetidae), mice and rats (family Muridae) are known to be main reservoirs for pathogenic Leptospira spp., it should be taken into account that shrews (family Soricidae) may likewise serve as reservoirs.
A former study showed that beech mast promotes population growth of bank voles (Myodes glareolus) in the subsequent year and that the resulting high bank vole abundance is positively associated to the number of reported human PUUV cases (Reil et al. 2015). The in-transformed proportional cover of beech and oak forest per district and the categorical variable of beech mast in the previous year (yes/no) were determined by our study group in a former study on hantavirus infection in the same small mammal samples (Drewes et al. in press). This former study demonstrated that the PUUV incidences in humans and the PUUV prevalence in bank voles, which were also investigated in the present study, is positively correlated with a combination of beech mast from the previous year and the proportion of forest cover in the respective district in Baden-Wuerttemberg, Germany (Drewes et al. 2016).
Beech mast in 2011 was the likely cause for high bank vole population densities and associated high PUUV prevalences in 2012 in contrast to the year 2013, when bank vole populations collapsed (no beech mast in 2012) (Drewes et al. 2016).
The prevalences of Leptospira spp. detected in all small mammals were nearly the same in 2012 and 2013, which suggests that neither bank vole abundance nor beech mast have influence on the spreading and maintenance of Leptospira spp. Beech mast is promoting population growth not only in bank vole populations but also in populations of yellow-necked mice, wood mice, and field voles (Imholt et al. 2016). However, it should be that Leptospira spp. are pathogens with a variety of reservoir host species that are not affected by beech mast whereas PUUV is exclusively harbored and transmitted by bank voles. This can explain the multi-annual dynamics of PUUV outbreaks in contrast to the more constant Leptospira prevalence in small mammals in forest environments.
In a previous study the PUUV prevalence in bank voles was significantly higher at sites with high hantavirus incidences in humans (Drewes et al. in press). In the present study the prevalences for Leptospira spp. in bank voles were also significantly higher at H sites with high human hantavirus incidences compared to L sites with low human hantavirus incidences. The notified incidences for leptospirosis in humans differed between 0.5 and 19 among districts and years (Fig. 1b and Table 2).The incidences for leptospirosis in 2012/2013 were slightly, but not significantly higher in the H districts compared to the L districts (Robert Koch Institute: SurvStat@RKI 2.0,
According to: Robert Koch Institute: SurvStat@RKI 2.0,
In earlier studies, the prevalences ranged between 7% and 54.4% for SWSV in Sorex spp. from Germany, Slovakia, Hungary, Finland, and the Czech Republic (Kang et al. 2009, Schlegel et al. 2012). The prevalence obtained from Sorex spp. in the present study is much lower (1.5%). The SWSV nucleotide sequence from the common shrew showed the highest similarity of 84% to a SWSV sequence originating from a common shrew in Ceske Budejovice, Czech Republic (Schlegel et al. 2012). Moreover, the prevalence was not positively linked to areas with high hantavirus incidences in humans, in line with the assumption that SWSV is not pathogenic to humans.
In conclusion, this study confirms bank voles, yellow-necked mice, and field voles as reservoirs of Leptospira spp. in south-western Germany. Moreover, this study points out that shrews may likewise serve as Leptospira spp. reservoir hosts. Results from this study suggest that the prevalence of pathogenic Leptospira spp. is positively linked to PUUV-infection in bank voles and to related high incidence areas of PUUV-infection in humans. As the prevalences were high and restricted to co-infections with PUUV at the H sites and as leptospirosis is causing a variety of nonspecific clinical symptoms, patients with an appropriate case history, such as previous exposure to contaminated water, should be regarded infected with either Leptospira spp. and/or PUUV.
Footnotes
Acknowledgments
Special thanks to Dana Rüster, Rayan Ababneh, Dörte Kaufmann, and Stefan Fischer for their excellent technical assistance and support. The provision of novel SWSV primer sequences by Mathias Schlegel, Hanan Sheikh Ali, and Julia Schneider is kindly acknowledged. Maysaa Dafalla would like to thank the German Academic Exchange Service (DAAD) for their support. The work of AO and MP was done under the frame of EurNegVec COST Action TD1303.
Author Disclosure Statement
No competing financial interests exist.