
Abstract
The spirochete Borrelia burgdorferi s.l. can enter into different eukaryotic cells. Intracellular localization of bacteria may cause many changes in different cell pathways like apoptosis-mediated caspase cascade. The present studies focused on gene expression associated with caspase cascade after normal human dermal fibroblasts (NHDF) infection with Borrelia garinii, Borrelia afzelii, and B. burgdorferi s.s. The use of oligonucleotide microarray technique enabled an expression level comparison of genes associated with caspase cascade in NHDF infected with spirochetes. The increased expression of genes associated with caspase cascade was observed in case of CASP5, CASP2, CARD10, CASP10, MALT1, and NLRP1. The decreased expression was observed in case of CASP4, CASP6, and CASP1. The mRNA expression for CASP3 was inhibited in cells infected with three genospecies of Borrelia. However, the intensity of fluorescence was not statistically significant. In addition, cell cultures were fixed and procedure of caspase-3 detection and the TUNEL assay were performed. The in situ caspase-3 detection procedure confirmed the results obtained from microarray analyses. Only several fluorescent signals were observed. Many apoptotic cells were detected in NHDF-infected cultures with all spirochete genospecies found using the TUNEL reaction.
Introduction
T
Caspases are grouped into two subfamilies: proapoptotic and proinflammatory. This classification is connected with major functions of caspases (Li and Yuan 2008). Caspases are family of enzymes important in the regulation of cell death and inflammation which are synthesized as inactive zymogens containing prodomain, a p20 large subunit and a p10 small subunit. Caspase zymogens are activated by proteolytic cleavage of the large and the small subunits and remove the prodomain. Native enzymes are endoproteases which hydrolyze peptide bonds (Li and Yuan 2008, McIIwain et al. 2015).
Proapoptotic caspases such as: caspase-2, -3, -6, -7, -8, -9, -10 are known to be mainly involved in the cell death signaling transduction, whereas proinflammatory caspases regulate inflammation process. Furthermore, proinflammatory caspases activate apoptosis process. Caspase-3 regulates apoptotic DNA fragmentation by proteolytic inactivation of DFF45 (DNA fragmentation factor-45)/ICAD (inhibitor of caspase-activated DNase). Then active DFF40/CAD (casapse-activated DNase) is released. DFF45/ICAD is a caspase-3 substrate that must be activated by cleaving before DNA fragmentation in apoptosis. The structure of caspase-3 is similar to currently known caspases. Caspase-3 is formed from zymogen that is cleaved into 17 and 12 kDa subunits. Its catalytic site contains the sulfhydryl group of Cys-285 and the imidazole ring of His-237. His-237 stabilizes the carbonyl group of the key aspartate residue, whereas Cys-285 attacks to ultimately cleave the peptide bond. Caspase-3 is responsible for chromatin condensation and DNA fragmentation. Moreover, caspase-3 may play a role in embryonic and hematopoietic stem cell differentiation (Wolf et al. 1999, Lechardeur et al. 2005, McIIwain et al. 2015).
Lyme disease is caused by spirochetes of Borrelia burgdorferi sensu lato complex. B. burgdorferi s.l. is divided into about 19 genospecies, of which Borrelia afzelii, Borrelia garinii, and B. burgdorferi sensu stricto are the most important human pathogens. This disease is transmitted by Ixodes ticks (Buczek et al. 2009, Stanek et al. 2011a, Biesiada et al. 2012, Jasik et al. 2015a, Pancewicz et al. 2015).
The spirochete B. burgdorferi s.l. can be extracellular and intracellular parasite, depending on their life cycle. They can enter into different eukaryotic cells, including fibroblasts, endothelial, and neuronal cells. B. burgdorferi has the ability to regulate a number of their genes that a spirochete adapts to diverse environmental conditions in ticks and mammalian tissue (Anguita et al. 2003, Jasik et al. 2015b). Intracellular localization of B. burgdorferi may cause many changes in different cell pathways like apoptosis mediated by caspase cascade.
The aim of this study was identification of gene expression associated with caspase cascade after infected normal human dermal fibroblasts (NHDF) with B. garinii, B. afzelii, and B. burgdorferi s.s. with microarray techniques supported with fluorescent microscopic methods. Furthermore, it was examined whether pathogens cause another way of apoptosis in NHDF.
Materials and Methods
In this study, CC-2511 line of human dermal fibroblast cells (NHDF; Lonza, Basel, Switzerland) infected with three genospecies of Borrelia: B. burgdorferi s.s.—B31 (ATCC 35210), B. garinii—20047 (ATCC 51383), and B. afzelii—VS 461 (ATCC 51567) were used.
Reference strains of Borrelia were cultured in BSK-H medium (Barbour–Stoenner–Kelly; Sigma-Aldrich, St. Louis, MO) in microaerophilic conditions at 35°C. Microscopic analysis of cultures and calculation of average number of bacteria in 1 mL of medium in Bürker chamber were performed after 7 days of growth.
Infected and uninfected cells of NHDF were cultured in FGM medium (Lonza) in culture flasks (Nunclon™ Sphera™ Flasks, culture area: 25 cm2) with bacteriological filters (Nunc, Wiesbaden, Germany). At the confluence stage, three genospecies of Borrelia were added to the tested NHDF cultures in the ratio of 10:1, bacteria to fibroblasts, to ensure a maximum effectiveness of infection. Infected cultures of NHDF cell line and the control one were incubated for 24 h.
Microarray method
The total RNA from treated and control NHDF cell line was extracted using TRIzol reagent (Invitrogen Life Technologies, California). Total RNA extracts were treated with DNase I (Fermentas International, Inc., Ontario, Canada) and purified by the used RNeasy Mini Kit (Qiagen Gmbh, Hilden, Germany). All steps of extraction were performed according to the manufacturer's instruction.
The quality of RNA was estimated by electrophoresis on 1% agarose gel stained with ethidium bromide (Sigma-Aldrich). The RNA concentration was determined using GeneQuant pro RNA/DNA Calculator (Pharmacia LKB Biochrom Ltd., Cambridge, United Kingdom).
Eight micrograms of total RNA was used to synthesize double-strand cDNA (SuperScript Choice system; Invitrogen Life Technologies). The synthesis of biotinylated cRNA was performed using the BioArray HighYield RNA Transcript Labeling Kit (Enzo Life Sciences, New York). Sample Cleanup Module was used for fragmentation of cRNA (Qiagen GmbH). Hybridization mixture was prepared using the GeneChip Expression 3′Amplification Reagents Hybridization Control Kit, and applied to hybridization with HG-U133A microarrays (Affymetrix, Inc., California). All steps were prepared according to the Gene Expression Analysis Technical Manual (Affymetrix, Inc.). Fluorescence intensity was measured using Affymetrix GeneArray Scanner 3000 7G (Affymetrix, Inc.).
Transcriptomes of 12 plates were analyzed: three for control cells of NHDF line, three for each group of NHDF infected with Borrelia. The oligonucleotide microarrays of Affymetrix HG-U133A enable analysis of 22 238 mRNA transcripts. For further study, 53 transcripts were selected in NetAffx Analysis Center database. These data from all arrays were analyzed using GeneSpring 12.6.1 platform (Agilent Technologies UK Limited, South Queensferry, United Kingdom) to identify differences between the control and treated NHDF cells. A significant differential gene expression (down or upregulation) was identified by a 1.1-fold change (FC) at p < 0.05 (one-way analysis of variance [ANOVA] and Tukey's post hoc test).
Fluorescent microscopic methods
Control NHDF cells and infected NHDF cells with three genospecies of Borrelia: B. garinii, B. afzelii, and B. burgdorferi s.s. were performed for this method. This material was fixed with paraformaldehyde. Culture flasks with infected and uninfected NHDF cells were cut to form microscope slides. Then the procedure of caspase-3 detection and the TUNEL assay were done. Additionally, all the slides with cell cultures were stained by DAPI method.
Caspase-3 detection
Infected and uninfected NHDF cells were washed in TBS (Sigma-Aldrich) for 5 min and in TBS containing 0.1% Triton X-100 (Sigma-Aldrich) for 5 min. Then the material was incubated with 5% BSA (Sigma-Aldrich) in TBS for 1.5 h at room temperature. Dry material (slides) was incubated with anti-caspase-3 antibody (Sigma-Aldrich) diluted in 5% BSA in TBS (1:200) for 12–24 h in a humidified chamber at 4°C. Slides were washed twice with TBS—each time for 5 min and incubated in goat anti-rabbit IgG biotin conjugated (Sigma-Aldrich) diluted 5% BSA in TBS (1:100) for 1.5 h in a humidified chamber at room temperature. After second wash, the slides were stained with streptavidin–fluorescein diluted in TBS (1:50) for 30 min in a humidified chamber, protected from light, at room temperature. After the third wash, material was stained with DAPI (1 μg/mL) (Sigma-Aldrich) for 10 min at room temperature in the dark. The washed slides were closed with cover slides using ProLong Gold Antifade Mountant (Molecular probes by Life Technologies, California) and examined under the fluorescent microscope (Olympus Corp., Tokyo, Japan).
TUNEL assay
Infected and uninfected NHDF cells were incubated in 1% sodium citrate for 2 min at 4°C. Slides were washed in TBS (Sigma-Aldrich) three times for 5 min and stained with a TUNEL (Tdt-mediated deoxyuridine triphosphate biotin/digoxigenin nick end labeling) reaction mixture for 1 h at 37°C in the dark. All steps were performed according to the manufacturer's protocol (the In situ Cell Death Detection Kit, TRM red; Roche, Mannheim, Germany). The material was analyzed using an Olympus BX60 fluorescent microscope (Olympus Corp.).
Results
In the first part of this study we performed microarray analysis of human dermal fibroblasts' transcriptomes, which were infected with three genospecies of Borrelia: B. garinii, B. afzelii, B. burgdorferi s.s (Fig. 1a, b). From among 22283 mRNA probes presented on the HG-U133A microarrays, 53 transcripts associated with caspase cascade were selected (Table 1). Transcripts were selected using Agilent GeneSpring GX Software.

Borrelia afzelii spirochetes (arrows) infecting NHDF cell
One-way ANOVA; FC >1.1.
ANOVA, analysis of variance; FC, fold change.
Twenty-two statistically significant differentially expressed transcripts were found by one-way ANOVA test (p < 0.05, FC >1.1). The samples were separated into groups according to Borrelia species. The first group was NHDF infected with B. afzelii, second group—B. garinii, third group—B. burgdorferi s.s. The fourth group was the control (NHDF cell line). Tukey's post hoc test was used to show similarities and differences between groups (Table 2).
Tukey's post hoc test; number of transcripts that exhibit differential expression are listed in bold (p < 0.05). Gray-shaded values-one-way ANOVA; p < 0.05.
[G], Borrelia garinii; [A], Borrelia afzelii; [SS], Borrelia burgdorferi s.s.
The increased expression of genes CASP10 and NLRP1, and inhibition of CASP4 were observed in the fibroblast infected with B. garinii, B. afzelii, and B. burgdorferi s.s. The increased expression of one gene MALT1 was shown in the fibroblasts infected with B. afzelii. In the group of fibroblasts infected with B. afzelii and B. garinii increased expression of CASP2 and CARD10 was observed. Moreover, increased expression of CASP5 was shown in the fibroblast infected with B. afzelii and B. burgdorferi s.s (Table 3).
↑, increased expression of mRNA; ↓, inhibited expression of mRNA.
The expression of mRNA for CASP3 was inhibited in the fibroblasts infected with all three genospecies of Borrelia. However, the intensity of fluorescence was not statistically significant.
In the second part of this study, the caspase-3 in situ detection procedure was done. Single positive signals were detected in the NHDF cells line infected with B. garinii, B. afzelii, and B. burgdorferi s.s. (Fig. 2a–c). There were no fluorescent signals in the control NHDF cells (Fig. 2d, e).
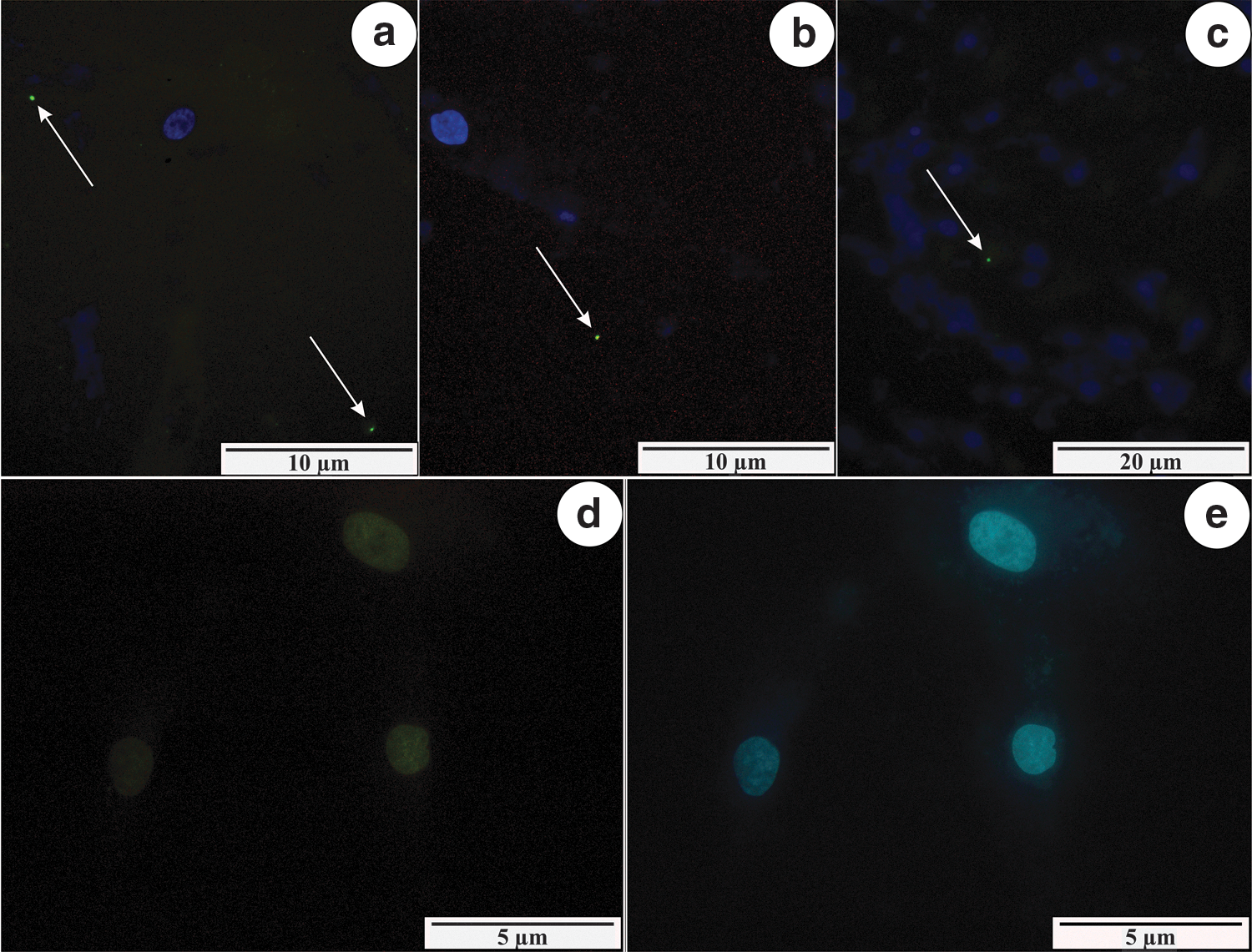
Single signals (arrows) showing the presence of caspase-3 in NHDF cells, infected with B. afzelii
The TUNEL reaction showed more fluorescent signals. DNA strand breaks were identified by labeling 3′-OH termini with modified nucleotides in an enzymatic reaction. Many positive signals were shown by all treated samples. The apoptotic cell death was detected in the treated materials infected with B. afzelii, B. garinii, and B. burgdorferi s.s. (Fig. 3a–c). The negative signals were detected in the control samples (Fig. 3d, e).

Signals showing the presence of apoptosis (red points) in NHDF cells, infected with B. afzelii
Discussion
Since many years, an increase in the prevalence of Lyme borreliosis is reported in Europe. There is the systematic increase of borreliosis cases reported in Poland during the last few years (Buczek et al. 2009, Cisak et al. 2012, Rozwadowska and Hudzik 2012, Lewandowska et al. 2013). According to data collected by the National Institute of Public Health - National Institute of Hygiene in Warsaw (Poland), in 2012, a total of 8794 infections were reported, whereas 12,763 in 2013 (Pancewicz et al. 2015).
B. burgdorferi spirochetes can enter to immune cells and other mammalian cells just like tick tissue. It allows bacteria to survive and infect host tissues. Spirochetes can hide from host immune system (Chmielewski and Tylewska-Wierzbanowska 2011). Our study showed the interaction between three methods proving the ability of B. burgdorferi to stimulate apoptosis. Microarray method confirmed the increase of many caspase pathway gene expressions in cells infected with three genospecies of Borrelia. Notwithstanding this fact, the expression of caspase-3 gene was inhibited. The same results were obtained using microscopic methods. Caspase-3 detection assay confirmed poor activation of apoptosis process in the human fibroblast cells. However, TUNEL procedure proved that apoptosis of infected NHDF cells was more intensive.
TUNEL method is based on DNA fragmentation. Short fragments (length of 180 base pairs) contain free 3′-OH ends that are responsible for the incorporation of labeled nucleotides. The reaction is catalyzed with the participation of polynucleotide TdT (Terminal deoxynucleotidyl transferase) and nucleotides labeled with digoxigenin (Gavrieli et al. 1992). Single signals in the caspase-3 detection method suggested that some cells might have died showing an apoptotic morphology, but without caspase involvement (Liang et al. 2001, Abraham and Shaham 2004).
Several models of caspase-independent pathways have been described. Alternative models of PCD independent of caspase activation were described, including autophagy, paraptosis, mitotic catastrophe, and apoptosis-like and necrosis-like PCD. In these death routes not only caspases, but also caplains, endonucleases, and cathepsins can activate cell death. These proteins can be distributed by cellular organelles, including mitochondria, lysosomes, and ER. They can act independently or together with each other. The advantage of these cell death pathways is protecting the organism against development of malignant diseases before cells become tumor cells. The knowledge of caspase-independent pathways is important for new cancer therapies (Bröker et al. 2005, Elmore 2007, Roomi et al. 2015).
Similar results were obtained by Chmielewski and Tylewska-Wierzbanowska (2011). They investigated the influence of B. afzelii, Coxiella burnetii, and Bartonella henselae on apoptosis measured as the level caspase-3 activity for 28 days. Only C. burnetii and B. henselae were able to inhibit such a process. The ability B. afzelii to inhibit apoptosis was limited because the caspase-3 activity in the early time after infection was low. Then the apoptosis process in the cells infected with B. afzelii was activated and the level of caspase-3 was higher. This process is connected with releasing bacteria to the extracellular space.
Diagnosis of Lyme disease is very difficult. It is connected with different immune responses, life cycle of the pathogen, and imperfect diagnostic method. In many cases a two-step serological diagnosis is not enough. It is also reported that some patient with chronic symptoms do not respond to antibiotic therapy (Aguero-Rosenfeld et al. 2005, Biesiada et al. 2012, Kubiak et al. 2012). Moreover, clinical manifestations of Lyme disease depend on the genospecies of Borrelia that caused the infection (Shapiro and Gerber 2000, Stanek et al. 2011b, Kubiak et al. 2012).
Problems with Lyme disease detection have many reasons. In the available literature most frequently described factors are antigenic variation, and morphological and immunological camouflage. Of importance is the phenomenon of chemotaxis of Borrelia spp. toward connective tissues and the ability to penetrate into different cells such as fibroblasts (Anguita et al. 2003, Chmielewski and Tylewska-Wierzbanowska 2011).
Apoptosis could have also played a significant role in the pathogenesis caused by B. burgdorferi s.l. strategy. Death of cells penetrated by B. burgdorferi s.l. does not activate the cause of inflammation.
Our apoptosis analysis of NHDF cells infected by spirochetes using different detection methods showed that Borrelia spp. are inducing mainly Caspase-independent cell death pathways of fibroblasts. Identifying the causes of this problem requires further biochemical research.
The knowledge on immunological mechanism enables introduction of advisable therapy for Lyme disease in the future.
Footnotes
Acknowledgment
This study was supported by the grant no. KNW-1-037/N/6/B from the Medical University of Silesia in Katowice, Poland.
Author Disclosure Statement
No competing financial interests exist.