
Abstract
Roads fragment vertebrate populations and affect the dynamics and dispersal patterns of vertebrate parasites. We evaluated how vertebrate ticks distribute near roads and road underpasses in human-caused road-fragmented landscapes in Seville, SW Spain. We sampled 49 stations with 93 individualized sampling points and assessed tick abundance. We explored the relationship between tick presence and abundance and distance to the nearest road and underpass (drainage culverts and other passages used by vertebrates), and landscape features through categorical regression and nonparametric statistics. The presence of the tick-borne pathogens Borrelia sp. and Bartonella sp. was also analyzed by PCR. We found preliminary evidence of high relative tick abundance next to roads and in the vicinity of road underpasses. Plant cover type was related to tick presence in this road context. Implications of road permeability and edge effect in patterns of vertebrate–tick relationships in road fragmentation contexts are discussed. Both Borrelia sp. and Bartonella sp. were detected in the ticks analyzed. This is the first report of these bacteria in ticks from Seville. The results confirm the potential risk of acquiring Lyme disease and bartonellosis in this area.
Introduction
R
Ticks have been earlier suggested to be attracted by roads, that is, through chemotactic gradients (McEnroe 1971). Global environmental change, ecosystem fragmentation, and especially roads could synergistically favor ticks and associated pathogens by attracting them to the road influence zone, increasing parasitizing rates near road edges (Estrada-Peña and de la Fuente 2014). Wildlife vector distribution and ability to populate distant places, and infect other animals could be also promoted by road underpasses. Besides, the vicinities of road underpasses are likely places where host species crossings would concentrate, especially if terrain structure or fences guide animals to the passage mouths (“funneling” effect) (Shoemaker and Servheen 2004).
Little work has been done about road edge effect on vertebrate ticks (McEnroe 1971) or the prevalence of parasites in road-killed fauna (Lorusso et al. 2011), but more effort has been devoted to tick distribution across landscapes and seasonality effects (Dobson et al. 2011, Renwick and Lambin 2013).
Some research has also been done on tick movement in relationship with host migration between contrasting habitats (Hoch et al. 2010, Ruiz-Fons and Gilbert 2010). Vegetation type and structure have been related with tick presence and risk for humans of picking up ticks in recreational areas, where ticks were abundant also near car parks in short grass (Dobson et al. 2011). Fragmentation of ecosystems and biodiversity loss is related to increased prevalence of ticks and tick-borne disease (Van Buskirk and Ostfeld 1998). However, we are not aware of researches on the role of road proximity or underpasses in tick distribution. If there is higher tick abundance in these sites than in places far from roads, there would be higher chances for contact with passing potential host, and hence in increased tick and pathogen dissemination across road-fragmented ecosystems.
In this article, we assessed if ticks were present more often near road edges and in the vicinity of road underpasses than far from them in road-fragmented landscapes in a sample of contrasting landscapes. We also assessed whether relevant landscape and habitat features relate with tick distribution around roads.
Borrelia and Bartonella are well-known tick-borne pathogens. Borrelia burgdorferi sensu lato complex is a diverse group of spirochetes that cause Lyme disease, the most diagnosed zoonosis in Europe, North America, and Asia (Radzijevskaja et al. 2011). Early symptoms are nonspecific, but in late stages of the disease, very variable symptoms may occur depending on the parasite species and the region (Stanek et al., 2011). Bartonella species cause a great variety of diseases depending on the species involved, from lymphadenopathy (reported in cat scratch disease) or bacillary angiomatosis to severe systemic disease in immunocompromised patients (Anderson and Neuman 1997). Therefore, we complement the study with the analyses of the presence of Borrelia and Bartonella, by molecular tools, in the ticks collected in a temperate endemic area of tick-borne diseases (Seville, Spain).
Materials and Methods
Field methods
The sampling was performed in the Seville province (Fig. 1), W Andalusia, Spain, from March to April 2012 and March to June 2013, and in June 2014. We combined the data taken from all collecting dates for analysis. We surveyed main landscape types between 10 and 375 meters altitude a.s.l., including built and periurban areas, agricultural areas, and forestry and natural areas. We differentiated two well-defined subregions: Quercus oak “dehesas” in the Sierra Morena mountain range (35 collecting sampling points) and the Guadalquivir river valley, with built and periurban areas and agricultural matrix (58 points). Overall, 93 sampling points were collected from 49 spatially separated sampling stations. We distributed sampling points haphazardly in the Seville province to encompass main ecological landscape units described above.

Map of the study area showing tick samplings.
Ticks were collected by the same person (J.D.D.) at all sites, by three different methods at each sampling point: (1) flagging; we used a 1-m2 piece of white flannel cloth attached to a pole to survey the ground and lower vegetation layers; we performed 10-min timed searches with the flag, starting at the UTM collecting point, proceeding outwards flagging the surrounding grounds and vegetation up to 5 meters from the point slowly covering a surface of ∼100 m2 at every bout, and stopping to recover nymphs and adults every 5–10 meter drag extensions. We collected all ticks found on the cloth in situ and stored them in absolute ethanol; (2) timed visual searches; we performed searches by 10-min units to detect ticks on branches, trunks, leaves, and the ground; and (3) passive sampling; all ticks found on the clothes of the recording/collecting person were collected. When several timed searches were consecutively done, we accumulated the expended sampling time in tick habitat to calculate ticks per unit time for “passive” sampling. We expressed tick abundance as tick number, and ticks recorded per 10 min as a relative abundance index.
At each sampling point we recorded the following: (1) altitude (in meters a.s.l.); (2) distance to the nearest road edge (in meters), (3) distance to the nearest underpass (in meters); both distances were measured whenever the conditions made it possible with a 1-meter precision laser telemeter and with a graduated tape for distances under 50 meters; since we could not ascertain exact distance to all underpasses, we classified them into eight distance intervals (0, 0–5, 5–10, 10–20, 20–50, 50–100, 100–500, 500–1000, and >1000); most underpasses presented signs of animal use, including humans (average pass width ±1 standard deviation [SD]) = 3.59 ± 2.42 meters); (4) grass cover (% area) and height (cm) within a radius of 5 meters around the sampling point; (5) dominant cover type: open terrain with a grassy/herb layer with no tree canopy above (Open), mostly ground under or near tree canopy (Tree), and mixed cover, mixture or mosaic of open and partially canopied patches (Mixed).
Statistical analysis of field data
We performed Kruskal–Wallis (K–W) tests to test the tick numeric abundance among distances to road edge and underpasses. We also used K–W tests to test for differences in grass cover and height on gradients of distance to roads and underpasses.
We performed a categorical regression (optimal scaling method, CatReg vs. 3.0, ran in SPSS 20) to analyze relationships between habitat descriptors as independent variables and tick presence/absence data per sampling site as dependent variable (De Leeuw 2005). The following factors were used as independent variables potentially affecting tick presence: (1) altitude; since it can serve as a proxy for humidity and saturation deficit gradients, and since they affect tick presence (Gilbert 2010); (2) dominant cover type; (3) grass layer height; (4) grass layer cover; (5) distance to road edge; and (6) distance to road underpasses. All variables were discretized by rank assignment before analysis.
Molecular analyses
Genomic DNA from a subsample of 53 ticks was extracted according to López et al. (2015). The presence of Borrelia sp. was examined by PCR. Standard PCR targeting a 276-bp fragment of the fla gene of B. burgdorferi s.l. was carried out following Picken (1992). Furthermore, a multiplex PCR that amplifies fragments with different size of the OspA gene, for Borrelia afzelii, Borrelia garini, and B. burgdorferi s.s. genospecies, was performed following Paulauskas et al. (2008). The ticks were also examined for Bartonella by a PCR targeting the rpoB gene as previously described, since this amplicon encompasses a specific zone, ∼800-bp, that is, useful for distinguishing Bartonella species (Renesto et al. 2001). PCR products were resolved by 1.5% agarose gel electrophoresis with RealSafe and visualized under ultraviolet (UV) light.
Results
Abundance and distribution pattern of ticks
We collected a total of 74 ticks (of which, 18 nymphs) at different distances from road edges and road underpasses in two different landscape contexts. Flagging, passive, and visual sampling yielded 33 (47.8% of ticks), 13, and 23 ticks, respectively (Fig. 2). Both mean number of ticks and relative abundance (ticks/10 min) were higher for “Sierra” (35 sampling points) than for “Valley” (58 points) environments (Table 1).

Numbers of ticks collected depending on dominant vegetation cover type and by different methods (blanket dragging—flagging, visual searches, and “passive” collecting). Passive collection refers to ticks adhered to clothes of the observer.
SD, standard deviation.
Relatively higher tick frequency and relative abundance were recorded closer to road edges than farther from them (Table 1 and Fig. 3). However, we found no significant differences in tick abundance among all distances to road edge (eight intervals) (χ2 7 = 12.154, p = 0.096). We found that tick abundance was significantly higher in sites near road underpasses (seven distance intervals) (χ2 6 = 18.604, p = 0.005) (Table 2). Sites with higher tick abundance and sites with tick presence were mostly located at distances ranging 0–50 meters from road edges. The higher tick relative abundance was recorded at distances up to 20 meters from road underpasses (Fig. 4). In three sites (Sierra Morena and oak woodland), adult ticks were visually detected questing along and across the asphalt right of way.

Box and whisker plots of number of ticks collected with distance to road edges. Asterisks are outliers.

Box and whisker plots of number of ticks collected with distance to the mouth of road passages. Circles are extreme values.
See Materials and Methods section for details. Significant outputs of the F test (analysis of variance) in boldface (α = 0.05).
df, degrees of freedom; SE, standard error.
The CatReg procedure identified major cover type, distance to road edge, and distance to nearest road underpass as significant predictors of tick presence (Table 2). Tick presence was thus significantly predicted by proximity to neighboring roads and drainage culverts, and other underpass types, although the gradients in grass cover and height did not apparently affect tick pattern. We found no relevant effects of altitude and grass cover and height at the sampling sites.
Grass cover varied significantly with distance to road edge (cover: χ2 7 = 16.377, p = 0.022) and underpasses (χ2 6 = 15.467, p = 0.017), although patterns were not clearly increasing or decreasing (Fig. 5). Grass height increased significantly with distance to road edge (χ2 7 = 37.044, p = 0.000) and underpasses (χ2 6 = 15.279, p = 0.018). We found a highly significant, but weak negative correlation, between tick numbers and grass height (Spearman's Rho = −0.337, p = 0.001), and no significant correlation of tick numbers with grass cover (Spearman's Rho = −0.002, p = 0.982).
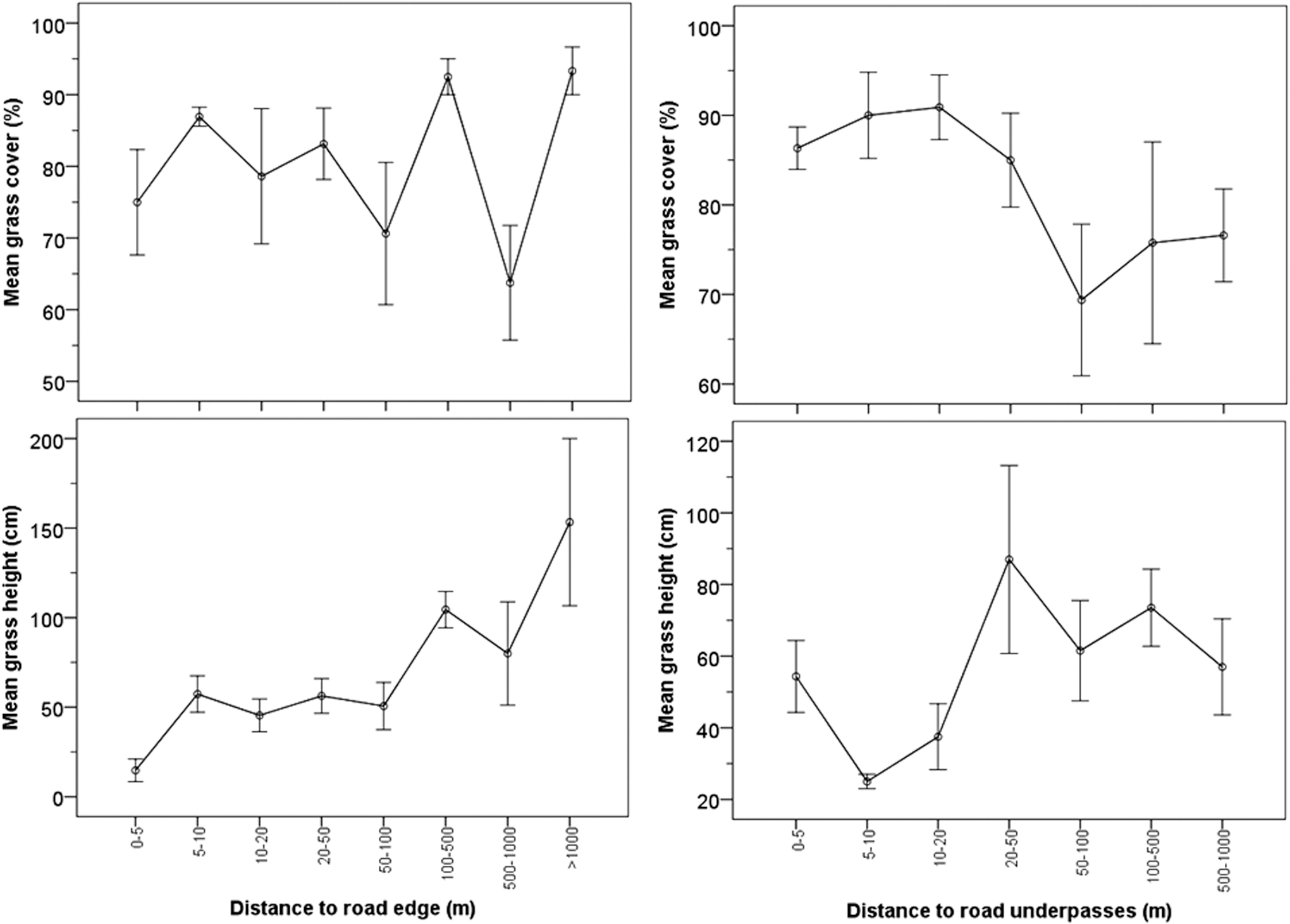
Variation in grass height and cover percentage per sampling site in relationship with distance to road edge and distance to nearest road underpass. Shown are mean ± 1 SE. SE, standard error.
Although we obtained a limited tick sample size, which could hamper a wide generalization of our results, tick presence and relative abundance were relatively higher near asphalt road edges. Distances to nearest underpass and road edge were selected as significant factors in categorical regression analysis. Other potentially influential variables, such as site elevation and grass height and cover, were very variable and site dependent, and did not contribute significantly to explain tick patterns in context of road or underpass proximity (Fig. 5). Grass height, for example, was generally higher far from than next to roads. Cover type was significantly related to tick presence, and we found higher tick prevalence in spots under tree canopies.
Tick-borne pathogens
Three out of the 53 (5.7%) ticks analyzed were positive for B. burgdorferi s.l., and they were obtained in valley and Sierra locations. However, only one tick (1.9%) harbored B. burgdorferi s.s., which was collected in the Sierra (Table 3). Regarding Bartonella, one out of 53 (1.9%) ticks amplified for the rpoB gene, concretely a sample from the valley (Table 3).
+, no. of ticks positive; n, no. of ticks analyzed; %, prevalence of ticks with Borrelia/Bartonella.
Discussion
Abundance and distribution pattern of ticks
First, we should not obviate our low sample size regarding number of ticks collected (compared to other works where ticks have been collected by hundreds). Reasons for limited catch size may reside, potentially, in the relatively early sampling season, adequacy for ticks of the surveyed sample of habitats, climatic determinants, or a combination of these factors. Climate influence on tick performance is determinant. Overall lower mean monthly temperatures, number of days with rain, and total monthly precipitation recorded in the immediately previous months, as well as in the previous year, 2011 (Diputación de Sevilla 2013), compared to our sampling dates, could be related to lowered reproductive success of ticks (Dobson et al. 2011, Hancock et al. 2011, Estrada-Peña and de la Fuente 2014).
Our results on tick spatial distribution regarding roads can be compared with those reported by Smith et al. (1946) and McEnroe (1971), in that ticks appeared to increase population densities near roads. Smith et al. (1946) marked a number of ticks that quested in parallel along roadsides and across the road right of way. In another study, ticks in the vicinity of a road or path system migrated toward the edge following a chemotactic CO2 gradient originated by road traffic, being “drained” within 30 meters of the road edge, and they also would respond positively to heat transmission from road surfaces (McEnroe 1971).
We detected adult ticks displaying an intense questing activity along rural asphalt verges and asphalt narrow roads with low traffic, in Quercus woodlands (Sierra Norte, Seville). Ticks staying prolonged time along road verges could increase their chances of finding hosts. Higher abundance of hosts near roads compared to more undisturbed grounds could explain, in part, differences in tick density near these corridors (Forman et al. 2002). Barely, there is also some early-published evidence of a road-crossing (over the asphalt right of way) activity in parasitic ticks (Smith et al. 1946). Nymphs of Ixodes pacificus have been shown to “ambush” (i.e., staying close to the freeing spot and moving only few centimeters a day) when seeking for hosts (Lane et al. 2009). Falco and Fish (1991) also described very limited recruitment distances (<2 meters) for Ixodes dammini. Adult ticks, however, can sometimes disperse longer distances and are capable of notable movements even with no vertebrate involvement, crossing ecosystem interfaces (Carroll and Schmidtmann 1996). Asymmetries in landscape structure between both sides of a road (Didham 2010) would be related with variation in vertebrate faunal composition. This would affect tick numbers and concentrations (Van Buskirk and Ostfeld 1998) creating differences in tick assemblage composition and abundance between landscapes surrounding both road edges.
Ticks are, nevertheless, more efficiently dispersed through their own hosts' movements (Ruiz-Fons and Gilbert 2010). Vertebrates moving across road-fragmented landscapes may suffer vehicle collisions (Trombulak and Frisell 2000), which could have some impact on the number of ticks passively crossing roads on vertebrate hosts (Nelder and Reeves 2005). Thus, some effect of roads either aiding or limiting tick movements would be expectable also on epidemic rates of their tick-borne pathogens. Vertebrates moving across habitats and tick biotopes can be an important dispersal vector for ticks during unfavorable climate periods (Lorusso et al. 2011). Crossing roads on vertebrate hosts can be an important mean of tick redistribution across contrasting landscapes in all seasons, but it can be the only or most preeminent way of moving for ticks during the climatically unfavorable season. On the other extreme, and interacting with the heat island effect of roads, global warming could create new areas apt for ticks and extend the climatically favorable breeding and questing season (Eisen 2008).
Tick abundance in the proximity of roads and road passages would be dependent on particular environmental features (Randolph 2000, Gilbert et al. 2010). However, as suggested by our results, distance to roads and road underpasses and the differentiated roadside habitat structure and microclimate could be also influential. Faunal passages, that is, underpasses, allowing increased permeability of transport infrastructure for small animal movements (Yanes et al. 1995, Clevenger et al. 2001, Grilo et al. 2008), could also intensify tick dispersal through host mobility, and hence parasite dissemination. Tick road-crossing rates (both on vertebrates and questing individuals) and colonization patterns of different areas would be worth studying, to assess effects of host-crossing rates, their own capabilities during the maximum questing season, and seasonality. Prospects for infection by tick pathogens and emergent diseases transported by road-crossing animals would be worthy of a more detailed evaluation, to establish the role of these linear elements which fragment the landscape and change vertebrate-parasite-pathogen relationships.
Tick-borne pathogens
Our results on pathogenicity are of public health relevance, since B. burgdorferi s.l. and Bartonella sp. are both emerging and underdiagnosed pathogens associated to severe diseases (Anderson and Neuman 1997, Sobrino and Cortázar 2008).
Our rates of infection by B. burgdorferi s.l. (5.7%) are lower than other studies on continental European tick populations (e.g., 18.6% prevalence for Ixodes ricinus) (Rauter and Hartung 2005). Nevertheless, lower infection rates have been detected for North American deer ticks Ixodes scapularis carrying Borrelia miyamotoi (ca. 1% of ticks) (Han et al. 2016).
Bartonella sp. is endemic in mainland Spain. Previous articles have shown higher prevalence than that found in this study (1.9%), concretely in carnivore fleas and ticks in Northern Spain (5.7%), where Bartonella henselae and Bartonella rochalimae were identified (Gerrikagoitia et al. 2012) or in small mammals (26.8%) in Basque Country, Catalonia, and Madrid (Gil et al. 2010). However, studies performed in humans and their pets in Madrid, where B. henselae DNA was detected, showed lower prevalence (0.15%) (Ayllón et al. 2012).
Conclusions
We found preliminary evidence of increased relative tick abundance next to roads and in the vicinity of road underpasses. Plant cover type was related to tick presence in this road context. Implications of road permeability and edge effect in patterns of vertebrate–tick relationships in road-fragmentation contexts are discussed. The potential role of ticks found in the transmission of Borrelia and Bartonella is confirmed, and our results, even based on low sample size, reveal an ample distribution within the Seville province. Roads may play a critical role in future expansion of these tick-borne diseases.
Footnotes
Acknowledgment
This research was partially funded by the grant “Ayuda p.p. 11.10, Grupos Emergentes” of the Universidad Pablo de Olavide, Seville, Spain, to J.D.D.
Author Disclosure Statement
No competing financial interests exist.