
Abstract
Orientia tsutsugamushi, the causative agent of scrub typhus in humans, is an obligate intracytosolic bacterium transmitted among animals and to humans by some species of larval trombiculid mites (chiggers) and is hosted mainly by rodents. In this study, we attempted detection of O. tsutsugamushi from blood and tissue samples of rodents trapped from different locations in Central India using PCR targeting the 56 kDa outer membrane protein gene and the 47 kDa high temperature transmembrane protein gene. A total of 59 rodent samples comprising 38 of blood collected from domestic and public surroundings and 21 of tissue from agricultural farm were included in this study. The 56 kDa outer membrane protein gene was detected from 10 of 59 samples by PCR, and the 47 kDa protein gene was detected from 4 of 59 samples by nested-PCR. Mites collected from the rodents were identified as Ornithonyssus bacoti, and one of five pooled samples was found to be positive for O. tsutsugamushi using PCR targeting 56 kDa outer membrane protein gene. Thus, perpetuation of O. tsutsugamushi among rodents and mites was detected constituting a potential public health concern.
Introduction
S
The 56 kDa type-specific antigen (TSA) gene codes for a dominant membrane protein, accounting for 10–15% of the total protein of O. tsutsugamushi, with an open reading frame of ∼1600-bp coding for 516–541 amino acids. The protein is generally involved in host cell invasion through the binding of fibronectin (Lin et al. 2011) and has four variable domains I–IV, which majorly contributes for large degree of antigenic variation in this gene. Due to this major antigenic variation, the 56 kDa protein gene (TSA) has become an important factor for molecular typing of O. tsutsugamushi, which led to identifications of several new subtypes, Japanese Gilliam, Japanese Karp, Kawasaki, Kuroki, Boryong TA763, TA636, and Shimokoshi, in addition to previously described Gilliam, Karp, and Kato based on sequence analysis of the gene from patients and rodents in different endemic regions of Scrub Typhus (Blacksell et al. 2008, Nakayama et al. 2010, Yang et al. 2012).
The 47 kDa high temperature requirement protein (HtrA) surface antigen is encoded by a gene within a family of serine proteases that has structural similarity and evolutionary homology with sequences found in almost all major bacterial and eukaryotic lineages, but not necessarily in all bacterial forms (Kim and Kim. 2005, Flannagan et al. 2007). The 47 kDa gene is an outer membrane protein/antigen also known as HtrA.
Many researchers focused on the prevalence of O. tsutsugamushi in chiggers parasitizing on wild rodents (Kuo et al. 2015, Candasamy et al. 2016). The detection of DNA of Orientia in rodents from Europe and Africa had been reported (Cosson et al. 2015). The rate of positivity for O. tsutsugamushi ranged from 0.9% to 5.7% in chiggers collected from wild rodents (Takahashi et al. 2004, Lee et al. 2011). In India, prevalence of scrub typhus has been reported in human cases (Varghese et al. 2015, Usha et al. 2016, Trowbridge et al. 2017). It has been reported that scrub typhus is much more common than previously thought in South India, and overall, the nature of the environment is favorable for scrub typhus (Trowbridge et al. 2017). Abundance and distribution of trombiculid mites and O. tsutsugamushi in rodents and shrews collected from Southern India have been reported (Candasamy et al. 2016). The objective of this study was to detect O. tsutsugamushi from blood and tissue samples collected from rodents trapped from domestic surroundings and agricultural farm in two regions of Central India using PCR.
Materials and Methods
Study site and rodent capture
Rodent study sites included domestic and public place surroundings from Nagpur in Central India, where there was a heavy population of rodents due to poor sanitary situations. Rodents were also trapped from agricultural farm in Raipur where mainly cereal crops (rice and wheat) were grown and had high number of live rodent burrows. Rodents were trapped randomly using traps baited with fish pieces during the month of monsoon, June 2016 to August 2016. At each place of study, numerous trap locations were selected. At each site at least 10 traps were laid. The details of samples collected are given in Table 1.
Ectoparasite collection
Mites, flies, and fleas were recovered from the skin of trapped rodents by Modified Tullgren Apparatus having an ordinary glass funnel comprising a series of four sieves with an incandescent light of 40 W shining above it. The funnel is suspended above a jar of 70% alcohol. The heat produced by the light bulb drives the mites down into the funnel and the mites fall into the alcohol.
Rodent processing
Captured rodents were euthanized using a chloroform saturated chamber. Blood samples were collected by cardiac puncture, aliquoted into tubes containing EDTA which was subsequently used for DNA extraction. The tissue samples were stored at −20°C.
DNA extraction
The blood samples were processed for DNA extraction by Martin's method. Briefly, 500 μL of blood was taken in microcentrifuge tubes, and 1 mL lysis buffer (0.22% NaCl, 0.015% Saponin, 1 mM EDTA, pH 7.5) was added. It was mixed thoroughly and centrifuged at 9300 × g for 3 min. The supernatant was discarded, and the procedure was repeated until pellet was clear of hemoglobin. The pellet was resuspended in 50–100 μL KTT 20 buffer (50 mM KCL, 10 mM tris-HCl, pH 8.0, 0.5% Tween 20, and 100 μg proteinase K per mL). Samples were incubated in water bath at 56°C for 2 h and later stored at −20°C. Spleen tissue samples were processed for DNA extraction using DNeasy Blood and Tissue Kit (QIAGEN, Hilden, Germany) as per manufacturer's instructions and then stored at −20°C till used.
PCR amplification
The PCR targeting the 56 kDa TSA gene was performed using primers OtsuF (AATGCTAGTGCAATGTCTG) and OtsuR (GGCATTATAGTAGGCTGA). This region encompasses ∼410-bp and contains the variable domain I-III hypervariable regions. The reaction mixture (25 μL) was prepared containing 12.5 μL of 2X GoTaq Green master mix (Promega), 1.5 μL of template, and 1 μL each (0.2 μM) of OtsuF and OtsuR primers. The PCR conditions consisted of denaturation at 94°C for 5 min, followed by 40 cycles, each consisting of denaturation at 94°C for 1 min, annealing at 60°C for 1 min, and extension at 72°C for 1 min. The final extension step was carried out for 10 min at 72°C.
The first round of nested PCR for the amplification of the 47 kDa gene was performed in 25 μL reaction volumes consisting of denaturation at 94°C for 5 min, followed by 40 cycles, each consisting of denaturation at 94°C for 30 s, annealing at 51°C for 1 min, and extension at 72°C for 1 min. The final elongation step was done for 10 min at 72°C. Primers used were OtsuFP555 (TCC TTT CGG TTT AAG AGG AAC A) and OtsuRP771 (GCA TTC AAC TGC TTC AAG TAC A) with an expected amplicon size of 238-bp. The second round of nested PCR was performed using the first product as the template DNA and 0.2 μM of each primer OtsuFP630 (AAC TGA TTT TAT TCA ACT AAT GCT GCT) and OtsuRP747 (TAT GCC TGA GTA AGA TAC RTG AAT RGA). The PCR conditions consisted of denaturation at 94°C for 10 min, followed by 35 cycles, each consisting of denaturation at 94°C for 30 s, annealing at 60°C for 1 min, extension at 72°C for 30 s, and the final elongation for 7 min, at 72°C with an expected amplicon size is 118-bp.
The PCR products were visualized with an ultraviolet transilluminator after agarose gel electrophoresis stained with ethidium bromide. Amplicons with a length of ∼410-bp (products of 56 kDa TSA gene) were sent for purification and sequencing to Eurofins India. DNA sequencing was carried out in an ABI 3730XL DNA Analyzer.
Phylogenetic analysis and sequence homologies
The sequences obtained were first trimmed to its appropriate size using ClustalW algorithm and then identified by comparison with sequences available in GenBank using BLAST (
Results
A total of 59 samples (blood and tissue) obtained from 2 regions of Central India were screened for O. tsutsugamushi using PCR targeting the 56 kDa TSA gene with a desired product of 410-bp (Fig. 1). Nested PCR was also performed targeting the 47 kDa HtrA gene with a product size of 118-bp (Fig. 2). Of the 59 samples tested, 10 were found to be positive by PCR targeting the 56 kDa TSA gene yielding a desired product of 410-bp with a prevalence of 16.94%. Out of 10 positive samples, 7 (18.42%) turned out to be positive from domestic and public surroundings of Nagpur, and 3 (16.66%) were found to be positive from agricultural farm at Raipur (Fig. 3).

Amplification of the 56 kDa TSA gene product. Lane M, 100-bp DNA ladder; lane 1, positive control; lane 2, test sample; lane 3, negative control; lanes 4–9, test samples. TSA, type-specific antigen.
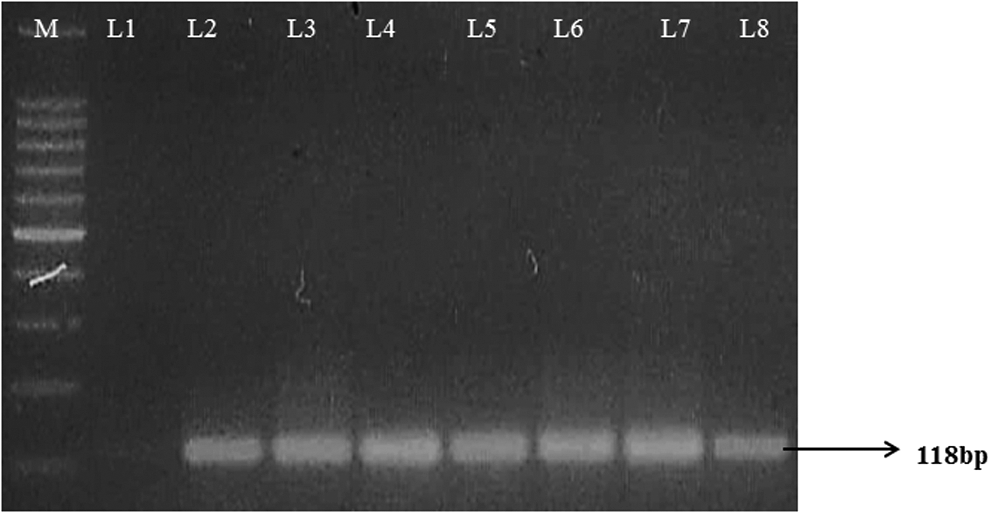
Amplification of the 47 kDa gene product of nested internal PCR. Lane M, 100-bp DNA ladder; lane 1, negative control; lanes 2–8, test samples.
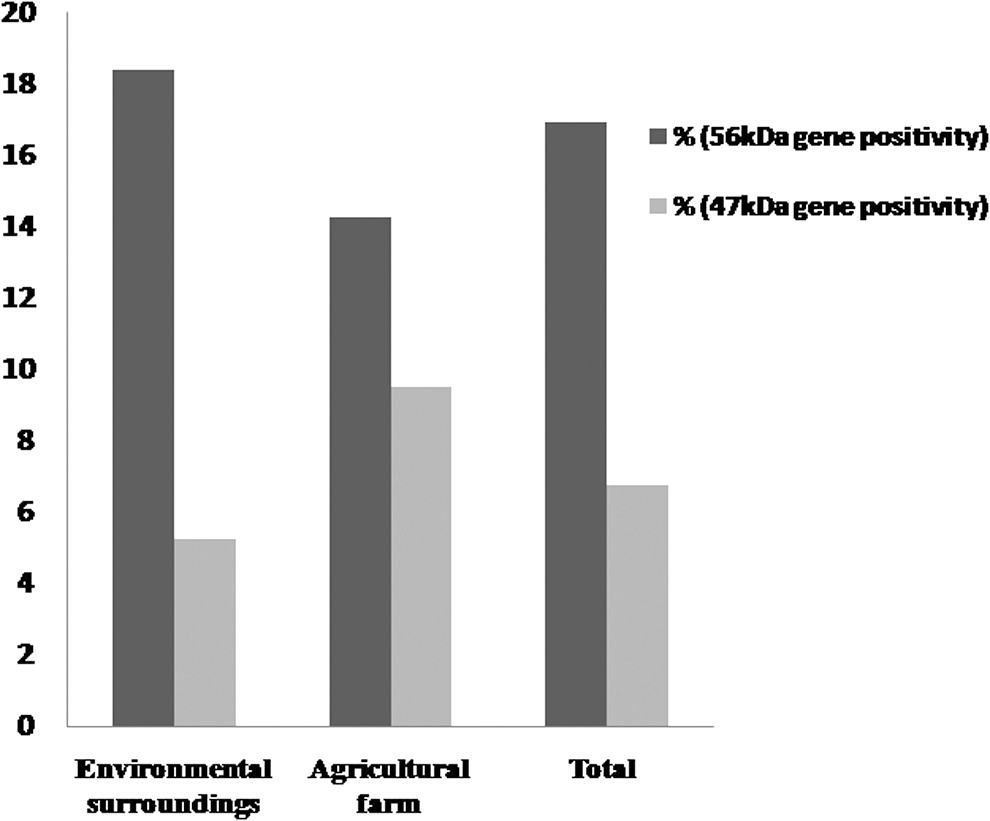
Prevalence of scrub typhus in rodent by PCR for the 56 and 47 kDa genes.
Nested PCR targeting the 47 kDa transmembrane gene was carried out. Four (6.77%) samples turned to be positive, of which two each were positive from domestic surroundings and agricultural farm.
Five pools of chigger mites were prepared from rodents trapped from domestic and public place surroundings. The mites were identified as Ornithonyssus bacoti by microscopic examination (Fig. 4). Out of five pools of mites isolated from rodents, one pool was found to be positive for O. tsutsugamushi by PCR targeting the 56 kDa TSA gene. Flies and fleas recovered were found to be negative for O. tsutsugamushi by PCR.

Lateral view of mites, Ornithonyssus bacoti, recovered from trapped rodents and identified by microscopic examination.
PCR products positive for the 56 kDa TSA gene from rodents and a PCR product of a mite pool were sent for sequencing for further confirmation.
The nucleotide sequence analysis of four samples and the mite pool for the gene encoding the 56 kDa TSA showed that these isolates were related to Karp-like strains, indicating predominance of Karp-like strains of O. tsutsugamushi in Nagpur region. O. tsutsugamushi detected in this study was closely related (94–97% similarity) to the strains isolated from Taiwan and Thailand (Fig. 5).

Phylogenetic tree showing the relationships among the 56 kDa TSA gene sequences of test isolates: R12, R26, R33, R37, Nag mite 1, and the reference strains.
Discussion
Rodents are important reservoirs of a number of human pathogens transmitted through arthropod vectors. These pathogens can cause acute febrile diseases in humans worldwide; however, the actual burdens of the diseases appear to be underestimated, as differential diagnosis on the basis of clinical signs and symptoms is confused by the presence of other tropical infectious diseases with similar signs and symptoms (Hotta et al. 2016). Human cases of scrub typhus are reported every year from various parts of India, including Central India (Rathi et al. 2011, Saxena et al. 2014). However, information on the presence of causative agent O. tsutsugamushi and its vectors from Central India is lacking.
In our study, we demonstrated the prevalence of scrub typhus by detection of O. tsutsugamushi in rodents by PCR from Central part of India. The overall prevalence observed in the present study for detection of O. tsutsugamushi in rodents was found to be 16.94% by PCR targeting the 56 kDa antigen gene. The prevalence rates ranging from 52.3% to 55.9% have been reported earlier in Thailand and Taiwan (Chareonviriyaphap et al. 2014, Kuo et al. 2015). The high prevalence reported earlier compared to the present study might be attributed to the high endemicity of the disease in Thailand and Taiwan.
Data based on nested PCR targeting 47 kDa transmembrane protein gene gave an overall prevalence of 6.77%. Earlier studies conducted in various parts of world reported the prevalence range of 15.2–55.9% (Song et al.1998, Lerdthusnee et al. 2008, Chareonviriyaphap et al. 2014, De et al. 2015, Kuo et al. 2015). The difference in the prevalence rate might be due to the seasonal variation of occurrence of mite in rodents' body, less sample size included in this study, habitat of the rodents, or places from where these were collected.
We identified circulating genotypes of scrub typhus in Nagpur region which included the analysis of variable domain of the 56 kDa TSA gene. The PCR amplicons of the samples collected from Nagpur were sequenced and analyzed. It was found that all the four sequences were related to Karp-like strain of O. tsutsugamushi isolated from Taiwan and Thailand with 94–97% similarity (Qiang et al. 2003, Wongprompitak et al. 2013).
Diagnosis of scrub typhus at molecular level is usually performed by PCR targeting the 60 kDa GroEL gene, which codes for a heat shock protein, the 47 kDa transmembrane protein gene, and the 56 kDa surface antigen gene (Jiang et al. 2004). Previous study (Qiang et al. 2003) reported phylogenetic characterization of O. tsutsugamushi isolated in Taiwan according to the sequence homologies of 56 kDa TSA genes. In the present study, strains detected were related to Karp strains, which were more predominant among the isolates from Taiwan. Kawasaki-related and STA-07 genotypes and distribution of O. tsutsugamushi in scrub typhus patients and rodents were reported in Shandong, northern China (Zheng et al. 2015); however, no Karp-related genotypes were reported. Genotypic analysis of eschar samples collected from patients with scrub typhus using the 56 kDa type-specific PCR and sequencing revealed predominance of Kato-like strains (61.5%) especially in the South and Northeast India, followed by Karp-like strains and Gilliam and Ikeda strains (Varghese et al. 2015). The phylogenetic analysis revealed that Karp-like strains might be predominant in Central part of India, although a higher number of rodent data collection needs to be done to confirm the predominance of the strain.
Considerable prevalence of O. tsutsugamushi was observed in domestic rodents. The mite identified as O. bacoti by microscopic examination is a tropical mite reported for the first time as a source of transmission of scrub typhus, contrasting the earlier reported mites responsible for the disease. It had been hypothesized that these rodents might play a role in transmission of scrub typhus by parasitizing chigger mites (Zhang et al. 2013). Vector species of chigger mites reported for the transmission of the disease included Leptotrombidium akamushi, Leptotrombidium pallidum, Leptotrombidium scutellare, Leptotrombidium deliense, and Leptotrombidium imphalum in Japan, Taiwan, Thailand, and China (Chang 1995, Frances et al. 1999, Takahashi et al. 2004, Kuo et al. 2011). In Korea, the most predominant species of chigger mites reported were L. scutellare, followed by L. pallidum, Leptotrombidium orientale, Leptotrombidium palpale, Neotrombicula tamiyai, Eushoengastia koreaensis, Neotrombicula gardellai, and Cheladonta ikaoensis (Lee et al. 2012). Schoengastiella ligula as the vector of scrub typhus outbreak in Kurseong, Darjeeling, India has been reported (Tilak et al. 2011). Since the collection of rodents was done randomly, rodent species could not be identified. One pool of mites was found to be positive for O. tsutsugamushi by PCR targeting the 56 kDa antigen gene. This confirmed that rodents played a primary role of transmission through the vector, chigger mites. Along with the chigger mites, flies and fleas were also collected to determine the exact mode of transmission which was found to be negative by PCR. Since a pool of mites turned to be positive by PCR, results confirmed mites as a transmitting vector. The results of PCR were confirmed by sequencing which showed Karp-like strain as a circulating genotype in mites also. This further confirmed Karp-like strain as a predominating strain of O. tsutsugamushi in rodents transmitted by chigger mites.
In conclusion, we detected O. tsutsugamushi genotype in rodents trapped from Central India which was Karp-like strain, focusing toward a potential public health concern. We also identified a mite, O. bacoti, for the first time as a vector of scrub typhus in addition to previously reported mite species. Further studies involving a large number of samples from rodents and chigger mites would give a clear scenario of transmission scrub typhus in Central India. The results suggested routine vector/pathogen surveillance at hot spots to initiate appropriate preventive measures.
Footnotes
Acknowledgments
This study was funded by Indian Council of Agricultural Research under Niche Area of Excellence for the project “Centre for Zoonoses” (2014/EP and HS) to S.P.C. The authors thank Dr. K.C. Munegowda, Kasturba Medical College (Manipal, India) for sharing the DNA of O. tsutsugamushi.
Author Disclosure Statement
No competing financial interests exist.