
Abstract
The Sari-Dzhas natural mountain focus of plague with an area of 5000 sq. km is located mainly in Kyrgyzstan. This enzootic area belongs to a group of Tien-Shan mountain plague foci and crosses the boundaries of Kazakhstan, Kyrgyzstan, and China. After collapse of Soviet Union, this area was not investigated fully, thence we have estimated current status of hosts and vectors comparing to 1977 in the Sari-Dzhas focus. For period 1977–2014, the number of the gray marmot (Marmota baibacina) as main host of plague has decreased twice due to regular chemical suppression of focal area, but now there is a tendency for population recovery. If negative impact of human activity (hunting, livestock, and tourism) will be minimized, the population density should rise in the near future, related to high reproduction score of animals. Significant increase of number of narrow-skulled vole (Microtus gregalis) observed on focus area in 1977–2017 can effort a risk of pathogen transmission in cross-populations of marmots provided by recovery of vector number between different hosts. For whole observation period in the Sari-Dzhas focus were described 33 species of marmot fleas with dominated Oropsylla silantiewi (67% from whole amount of fleas) and Radinopsilla li ventricosa (6.5%). Deep dustation of marmot burrows by 10% dichlorodiphenyltrichloroethane (DDT) conducted in 1980s caused significant decrease of ectoparasite abundance: the number of specific fleas is strongly diminished, while recovery of ticks and lice number is faster. At the same time, Yersinia pestis constantly circulates in the focal area, which is comfortable for interacting of human populations and pathogen hosts. Last plague epizootic was recorded in 1983 and recent outbreaks among gray marmot and its vectors with varying intensity are registered again starting from 2012. Thus, the Sari-Dzhas natural plague focus remains active with all links of plague epizootic cycle (marmot-vector-rodent) and needs strong monitoring every year.
Introduction
T
Marmots are well known as a host of many zoonotic diseases, but greatest danger to humans is plague (Pole 2006). Ectoparasites of marmots are actively involved in the transmission of the plague pathogen between animals (Bibikov 1967, Suntsov and Suntsova 2006). Marmot fleas are represented mainly by the following species: Oropsylla silantiewi; Rhadinopsylla li ventricosa; Ceratophyllus lebedewi; and Pullex irritans: among them, the two first species are specific (Rall 1960, Suntsov and Suntsova 2006).
The Sari-Dzhas plague focus was active for a long time starting from 1913 till 2016 (Abdikarimov et al. 2017). However, after the collapse of the Soviet Union, this territory was not monitored due to the destruction of unified antiplague system.
In this term, the purpose of this study was to estimate current ecological and epizootic status of populations of the plague hosts and vectors in the Sari-Dzhas plague focus by comparing it with a situation in 1977.
Materials and Methods
In total, the article includes information of analyses of 684 marmots, 313 small rodents and other wild animals, and 1240 ectoparasites.
Field work. The field surveys were conducted in different sites of the Sari-Dzhas focus in 2014, 2015, 2016, and 2017. All activities were organized by the Republican Center of Quarantine and Dangerous Infections (RCQDI), conducted by the Karakol Anti-plague Branch (KAPD) during July 2014, July–August 2015, June–July 2016, and June–July 2017. For a comparison purpose, we used archived data of epizootological survey of the Sari-Dzhas focus conducted in 1977.
All manipulations with wild and laboratory animals and handling plague strains were conducted according to the protocols approved by the Kyrgyz government regulations (Burgasov 1967, Aubakirov et al.1990, Kuregyan 1995, Korneev 2002, Aytkuluyev 2010, Ezhlova 2011). The animal work was performed according to the Regulation of the State Agency for Environmental and Forest Protection Facilities at the Government of the Kyrgyz Republic. Animal work in the field was conducted according to the following protocols: protocol 000144-KC for trapping marmots, other rodents, and analysis of their nests approved on May 6, 2014; protocol # 000168-KC approved on May 22, 2015; protocol #000173-KC approved on May 17, 2016; and protocol #000003-KC approved on March 5, 2017. Each permit to collecting a certain number of animals (400 marmots, 400 rodents of other species, and ten nests of marmots and rodents) was issued and used during the fixed period of time (from June 1 to August 30).
Each field work was conducted at a certain area selected based on historical data for previous plague activities (Weaver et al. 2014). Each field work episode included a survey of natural habitats of marmots within 800–1000 sq. km of plague-endemic territory.
To estimate the number of animals, direct and indirect methods were applied: trapping of marmots by traps and visual counting of active animals and burrows used by animals during the current season. The visual estimation of animals was carried out during period of highest activity of animals (during clear days from 6 to 11 and from 17 to 20 h). All observed marmots were counted in each site following a path of minimal length of 5 km and the 200 m wide strip. Due to the fact that some of the animals remained in burrows and could not be observed, the correction index proposed by Aytkuluyev (2010) was used to improve the calculation of animals per 1 sq. km.
A number of small rodents, such as mice, voles, and hamsters, were estimated by counting animals captured by snap traps (Aubakirov et al. 1990, Ezhlova 2011). The predators were estimated by the same way (trapping and visual observing). Each animal trapped during the night was analyzed by zoological and bacteriological methods.
Physiological status of animals
All caught animals were weighed, their sex and age group were recorded, and length of the body was measured. The inner fat, stomach, and intestines were extracted and weighed separately. For determining weight of rodents, weight of the stomach and intestines was subtracted from the total weight of the animal. The accumulation of inner fat was estimated as a ratio of inner fat to the body weight.
For estimation of reproductive activity and retrospective analysis of generative processes in populations, we used such traits as a number of embryos per pregnant female and percentage of reproductively active adult females among total number of females.
The reproductive status of marmots (R) was estimated as an equation proposed by Ezhlova (2011); specifically, the average number of embryos per pregnant female was multiplied by percentage of reproductively active females.
Parasitological analysis
The ectoparasites of marmots were collected by combing caught animals (“body” ectoparasites) and from rodent burrows and nests (“off-host” ectoparasites, Ezhlova 2011). The number of ectoparasites was estimated as Index of Abundance (AI): by dividing the total number of ectoparasites collected from animals by total number of inspected trapped animals.
Identification of ectoparasites was performed by using entomological keys (Ioff and Skalon 1954, Ioff et al. 1965).
Bacteriological analysis
After identification of the species, insects were prepared for group or individual inoculation on selective medium for plague as Hottinger agar, pH 7.2 (Ioff et al. 1965, Suleymenov and Abdirasilova 2014).
Identification of isolated bacteria was carried out by standard microbiological microscopy of smears, Pokrovsky's test for Yersinia pestis, pseudo tuberculosis bacterial phages, and bacteriophage for Y. pestis (L-413-C) produced by the Kazakh Scientific Center for Quarantine and Zoonotic Diseases (KSCQZD); cultivation on selective medium; and serological indirect hemagglutination assay with erythrocyte immunoglobulin diagnosticum and erythrocyte antigen diagnosticum produced by KSCQZD.
The map of landscape-epizootic regions (LER, Aubakirov et al. 1990) of the Sari-Dzhas focus of plague was designed using Geographic Information System MapInfo Professional 7.8. For their construction, we used the geographic objects digitized from topographic maps of a scale of 1: 100,000, as well as the Digital Elevation Model GDEM2. The maps are constructed in Universal Transverse Mercator—the Mercater projection (WGS 84). The boundaries of the LER are plotted along the boundaries of the sectors (Shabunin et al. 2017). The places of animal trapping are marked by the following data: 1977 and 2014—according to the archive report of KPAD; and 2015, 2016, and 2017—for GPS points obtained during the surveys of area.
Results
Field epizootical and epidemiological survey were made in 2014–2017 in the area of three local sites of the Sari-Dzhas natural plague focus: Inilchek-Kaindy, Kok-Dzhar, and the Sari-Dzhas (Fig. 1). The Inilchek-Kaindy site occupies 900 sq. km of southern part of the Sari-Dzhaz focus in the basins of Inilchek and Kaindy rivers. The typical landscapes are high-mountainous steppes and semideserts separated by mountain ridges. The climate is warmer compared to other sites. The vegetation varies from dry mountainous steppe and variety of cereals on syrts to birch, spruce, and various species of shrubs. The Kok-Dzhar site (750 sq. km) is located in the basins of the Turuk and Zhynalach rivers (Fig. 1). The terrain is slightly intersected by intermontane valleys with varietal meadow plants; there are also bushes and spruce forests. The climate is sharply continental. The Sari-Dzhas site locates in the Sari-Dzhas river basin within an area 1220 sq. km at an altitude of 2.700–3.600 m (Mamatkanov et al. 2014). The climate is severe: winter is long and cold, summer is cool, and the precipitation rate is 300 mm per year. In 2014–2017 we have inspected total 2.400 sq. km of focus area. In 1977, when the antiplague field works were supported and organized by the Soviet Government, only a 1-year expedition had worked 60–90 days and covered 1000 sq. km. (Fig. 1).
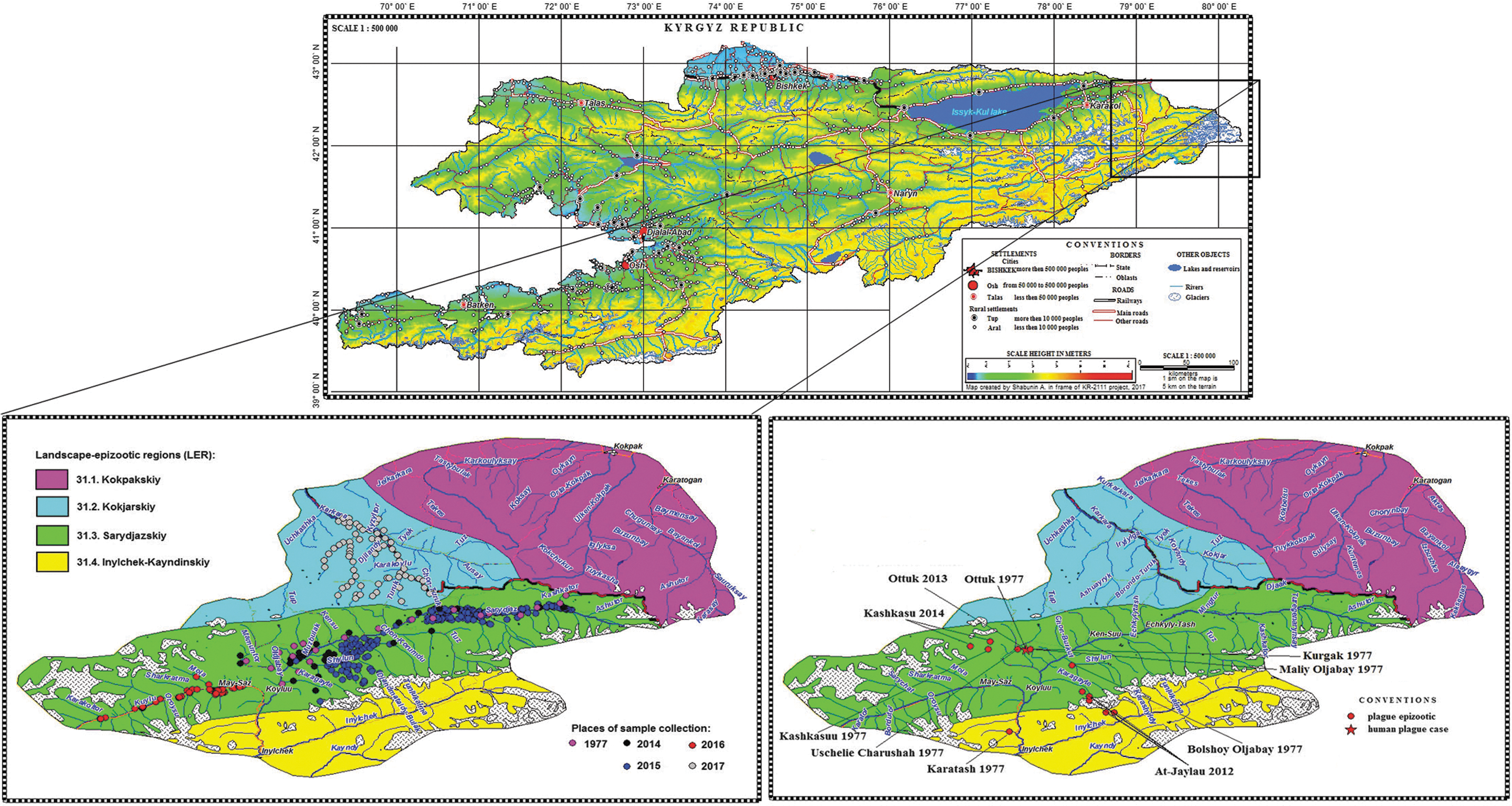
Localization of the Sari-Dzhas natural plague focus, conducted field activities, and isolated plague strains. Figure 1 can be seen in greater detail at
The number and distribution of the main host of plague–gray marmot
In 2014–2017, the visual number of marmots on all sites was similar to those in 1977 (Table 1).
The average number of marmots on the surveyed territory varies by year: in 1977, this trait was relatively high; in 2014, on the same sites, it fell twice, and then increased in 2015, while the maximum number on certain sectors was 51.7, and returned to the level of 2014 in 2017 (Table 1).
Nutritional status of marmots
Table 2 presents the results of accumulation of internal fat of marmots during observation.
The accumulation of internal fat in all age groups over the entire period (4 years) was 0.5–2.0% (Table 2). This indicator in young animals of all groups was slower as by subadult and adult. The data of recent years do not differ much from those of 1977, excepting nonreproductive females, in which accumulation of internal fat was by 1.3% higher than now.
Population composition
In 2014–2017 adult males and females made 26% and 22% of the total population, respectively; among them young marmots (♂ ♀ sub-adult and juvenis) have 51.2%, and the proportion of young females and males varied by years. In contrast to young animals there was large number of adults (64.0%), including more females in 2015. In 2017, there were also more females. For comparison, in 1977, adult males and females made 19.5% and 32.2% of the total population, respectively. Young marmots made 48%: among them, appositive, males were almost two times more than females (Fig. 2).
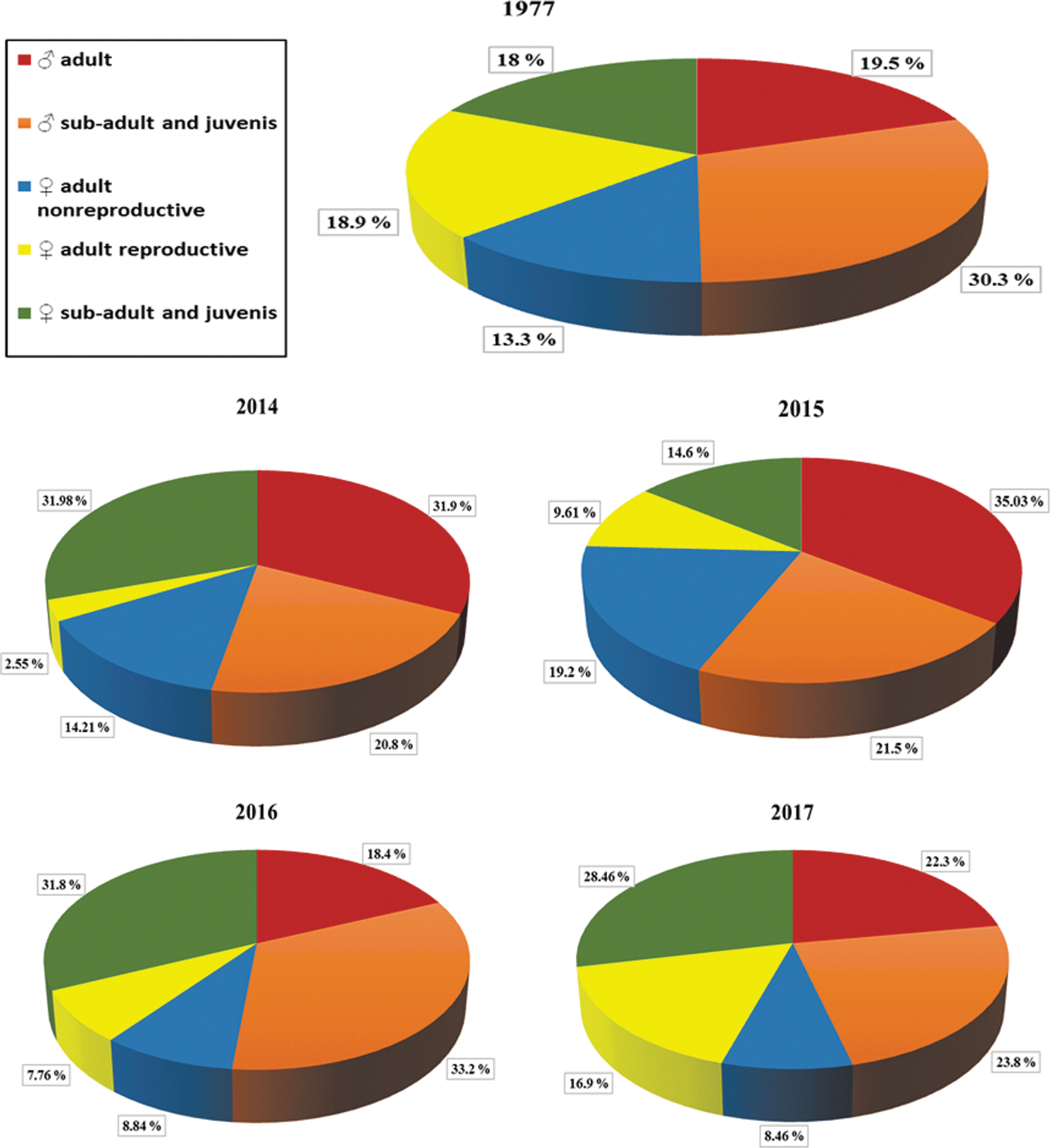
The composition of marmot population in 1977 and 2014–2017.
Reproductive status
In 1977, the total number of females was very high, but during the period of 2014–2017, this parameter decreased significantly (Fig. 3). The percentage of reproductive adult females in 1977 was 71%, while in 2014, it was 15%, but almost reached the previous level in 2017 (Figs. 2 and 3). The dynamics of nonreproductive adult females shows an increase in 2014 compared to 1977, but later on, returned to the level of 1977 (Fig. 3). In average, one pregnant female had 6.3 embryos. Starting from 2014, there is an evident tendency to increase the relative reproduction score, which almost reaches the level of 1977.
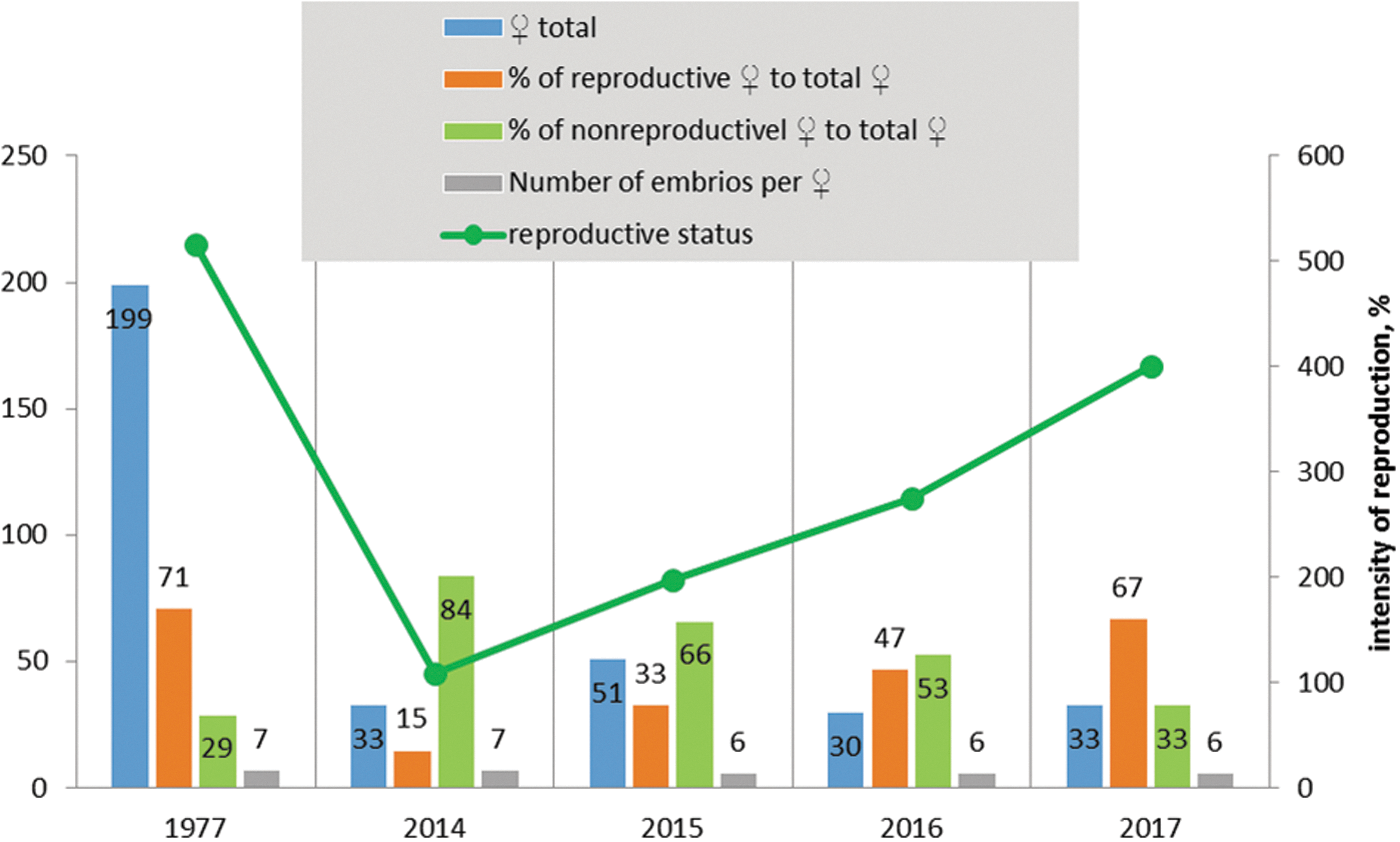
The dynamics of marmots' reproduction in the Sari-Dzhas natural plague focus in 2014–2017.
The number of small mammals
Among mouse-like rodents, the most numerous remains Microtus gregalis and its number is significantly increasing in recent years. All other mouse species, such as the forest mouse, gray hamster, and pika, were significant less (Table 3).
Recording of the number of vectors of plague in the Sari-Dzhas natural focus
In 1977, the AI for all “body” ectoparasites (fleas, ticks, and lice) of marmots has shown very high values (Table 4). This was the main reason for deep dustation of the focal area with 10% dichlorodiphenyltrichloroethane (DDT), conducted in 1980 s by RCQDI and KPAD. As a result, the AI of all ectoparasites decreased significantly (Table 4). In 2014–2017, the average AI for fleas and ticks remains still reduced, while the number of lice has fully recovered (Table 4). The composition of specific body fleas has also changed—in 1977, the main vectors of marmot plague were O. silantiewi and R. li ventricosa (65% and 30% of all fleas, respectively, Table 4). In 2014–2017, the percentage has shifted toward O. silantiewi, which dominates with average 92% and R. li ventricosa has only 8% (Table 4). As to annual dynamics, the Or. silantiewi had maximal AI in 1977, which dropped by four times in 2014, although now it has reached the previous level on the whole observed area. AI of R. li ventricosa in the body of marmot decreased from 0.41 in 1977 to 0.03 in 2014 and is still low in most areas of the focus, excepting few sectors (Boron-Turuk valley and Molo gorge) where it has fully recovered in 2016–2017.
p < 0.05.
More precise changes were identified by the number of nest ectoparasites of marmots: the AI for fleas in 1977 was extremely high with evident prevalence of R. li ventricosa (Table 4). During 40 years, this trait has decreased to four to five times: only in 2014 and 2016 were observed both specific fleas with prevalence of R. li ventricosa. The ticks were found once in 2016.
Epizootic activity of Sari-Dzhas natural plague focus in 1977 and 2012–2017.
In total, for whole observed period in Sari-Dzhas natural plague focus, 45 plague strains were isolated (Fig. 1) with maximum in 1977: from marmots—10; ectoparasites—16; among them O. silantiewi—8; R. li ventricosa—1, thick—7 (Table 5).
Starting from 2012, the sporadic plague epizootics were recorded again. Among them was one case of human bubonic plague in 2013 (Popov 2013).
Discussion
Density of gray marmot populations in the Sari-Dzhas plague focus was significantly reduced after active control measures during the period from 1956 till 1967 using cyanide (archived data from KPAD, unpublished; Pole 2003). Similar preventive measures were widely used against the gray and red marmots in the Verkhnenaryn and Alai foci of Kyrgyzstan, Siberian, and Mongolian plague foci (Pole 2003, Popkov et al. 2013). Our data showed that average number of marmot in the Sari-Dzhas focus after 40 years of intensive marmot control activities has still not reached the level of 1977. It is possible that various unfavorable environmental factors (deficit precipitation rate, droughts) and changes in human activity (livestock grazing) lead to transformation of local ecosystems accompanied by decrease in number of marmots. In general, irregular spatial density and the nutritional status of marmots estimated as accumulation of internal fat in the Sari-Dzhas plague focus are correspondent to biotic (landscape, climatic conditions, and forage) and abiotic factors (increased human activity like hunting, livestock grazing, and tourism). Livestock density and recreational activities intensified very much for recent years in Kyrgyzstan. Hunting on marmots has also increased because of their commercial value (Aubakirov et al. 1990, Pole 2015) and has been officially authorized by Kyrgyz Government in 2015.
Previously it was mentioned a better nutritional status of nonreproductive adult females comparing to those reproductive after hibernation (Bibikov 1967, Armitage 2014). We suppose that conversely rate of fat accumulation in these two groups showed in 2014–2017 can be related to depletion of fodder base due to concurrence with livestock animals grazing in the same areas as marmots. In general, based on high percentage of females participating in reproduction this year (40.5%), a part of nonreproductive females (12.7%), as well as high proportion of young females (26.8%), we can predict an increase in number of marmots in the near future, in case a negative impact of human activity is minimized.
Significant increase in density of narrow-skulled voles (M. gregalis) in 2014–2017 in the Sari-Dzhas focus can present a risk of plague transmission between populations of marmots. The degree of contact between narrow-skulled voles and marmots is very high as their colonies overlap colonies of marmots (Krylov 1964, Bibikov 1967, Aubakirov et al. 1990, Pole 2015). In some cases, there were shown facts of ectoparasites exchange between voles and marmots (Ioff 1949, Sludskiy 1978, Suntsov and Suntsova 2006). Six plague strains isolated from M. gregalis in different years and one strain from their flea (C. migratorius) in 2012 indicate that mouse-like rodents are involved in the circulation of plague pathogens in the Sari-Dzhas natural focus.
Fleas are key vectors of pathogen transmission in plague foci (Bibikov 1967, Suntsov and Suntsova 2006). Totally, for an entire period of observation in the Sari-Dzhas focus, there were 33 species of marmot fleas found with dominant O. silantiewi and R. li ventricosa (Krylov 1964, Bibikov 1967, Suntsov and Suntsova 2006, Kolesnikov 2011, Pole 2015). Before intensive application of DDT in this territory in 1980 s, the level of vectors was much higher: average annual AI of fleas was 4.2 and for R. li ventricosa 3.05 (Aubakirov et al. 1990). This study demonstrated that deep total dustation of marmot burrows has a long-term effect on plague circulation within Sari-Dzhas focus that manifested in a decrease of number of plague vectors and isolated strains. The same long-term effect of DDT on ectoparasites population was described in the Alai natural focus of Kyrgyzstan (Altybaev and Kulnazarov 2004), and in the Siberian foci of Russia (Popkov et al. 2013), Mauritania, and Morocco (Malek et al. 2016). The prevalence of R. li ventricosa in the nests of marmots after long-term effect of dustation could be explained by the fact that this flea is a nest ectoparasite and withstands starvation significantly longer in the case of host death compared to O. silantiewi, which needs more frequent bloodfeeding (Sludskiy 2014, Kolesnikov 1997, Suntsov and Suntsova 2006). These physiological properties provide fleas to overwinter and keep the pathogen persistence for long period. Probably, unique diversity of landscapes historically formed in the Sari-Dzhas area contributes to such complicated effect of dustation on recovery of ectoparasite numbers (Mamatkhanov et al. 2014, Bibikov 1967).
Thus, the activation of plague focus accompanied with a recovery of carriers and vectors, observed in 37 years after chemical control and dustation of marmot populations, presents certain epizootological and epidemiological risks. The circulation of plague pathogen across the epizootic territory occurs constantly with different intensity, depending on the current state of the plague parasitic system, abiotic and anthropogenic factors. In Kyrgyzstan, efforts are made to identify factors, which determine long-term plague foci activity, and to strengthen surveillance of animal hosts and their ectoparasites.
Conclusion
The Sari-Dzhaz transboundary area located between Kazakhstan, Kyrgyzstan, and China is the most active among mountain plague foci of Kyrgyzstan. Plague epizooties among wild animals has been recorded here regularly since 1913. In 2014–2017, population density of main plague host, the gray marmot, slowly recovered with high reproduction score, but some conditions (intensive livestock, officially authorized hunting of marmot [Government Resolution of KR, 2015], and tourism) have a negative impact on marmot populations. Increase of number of small rodents and their contact with marmots with exchange of ectoparasites like ticks and lice during last 37 years relate to heterogeneity of local landscape-climatic conditions. Thus, the geography of the area, spatial organization of gray marmots and population density of animals, their family-colonial lifestyle, different ages of animals in the families, the long-term use of the winter burrows, and year-round parasitism of specific fleas (Or. silantiewi and R. li ventricosa) on the marmots provide good conditions for continuing circulation of Y. pestis in the Sari-Dzhas plague focus. Efforts are required to strengthen surveillance of host animals and ectoparasites.
Footnotes
Acknowledgments
This study was supported by the International Science and Technology Center (ISTC). The field epizootical surveys were funded through regular projects #КR-1784 and #КR-2111 by financial support of the Government of Canada, EU, and USA. We thank Dr. Kingsley Amoako (FAI, Canada), Dr. Michael Kosoy (CDC, USA), Dr. Serge Morand (CIRAD, France), and Prof. Vladimir Motin (UTMB, USA) for scientific support of these surveys and their comments on draft of the article.
Author Disclosure Statement:
No conflicting financial interests exist.