
Abstract
The beef tapeworm Taenia saginata, which causes taeniosis in humans and cysticercosis in cattle, is of medical and economic importance. Understanding the parasite's genetic population structure may help to analyze transmission patterns and aid in the development of control measures. As information on sequence variability is scarce for European isolates, this study aimed to elucidate the intraspecific genetic variability of T. saginata cysts from German cattle by sequence comparison of the mitochondrial cytochrome c oxidase subunit 1 (cox1) and NADH dehydrogenase subunit 5 (nad5) genes, in relationship to sequences from other geographical origins. Cysts were collected from northern German, Swiss, and Belgian cattle. Moreover, proglottids from an adult T. saginata specimen from Palestine were included. Amplification and Sanger sequencing of the cox1 gene was successful for 57 samples (45 German, 9 Swiss, 2 Belgian, 1 Palestinian), whereas 32 sequences were obtained for the nad5 gene (21 German, 10 Swiss, 1 Palestinian). For German isolates, sequence comparison revealed minor genetic variability with two polymorphic sites and mutations in both genes. Three haplotypes with haplotype diversity of 0.088 for cox1 and 0.186 for nad5, as well as nucleotide diversities of 0.00028 and 0.00095, respectively, were observed. Comparison of the cox1 gene sequence of German isolates with other European, African, American, and Asian isolates obtained from National Center for Biotechnology Information (total of 71 sequences) raised 11 polymorphic sites and mutations as well as 10 haplotypes (haplotype diversity: 0.239; nucleotide diversity: 0.00097). Although nad5 sequence comparison comprised less sequences (N = 33), analyses revealed 11 polymorphic sites, 12 mutation sites, and 7 haplotypes (haplotype diversity: 0.335, nucleotide diversity: 0.00391), indicating a better resolution of genetic variability compared to cox1. Thus, nad5 may be particularly useful for in-depth studies on genetic divergence of T. saginata.
Introduction
T
Cattle become infected via ingestion of food or water contaminated with parasite eggs originating from human feces. After ingestion, the oncosphere hatches and migrates through the circulatory system to skeletal and myocardial muscles, where it develops into the infective metacestode, known as Cysticercus bovis, within 10–16 weeks. Cysticerci usually remain infective for about 9 months until they die and calcify (Froyd 1964). Humans acquire infection by ingesting the viable cysticerci within raw or undercooked beef. Typically, taeniosis is asymptomatic in humans, but may manifest with abdominal pain, nausea, weight loss, diarrhea, or obstipation (Dorny and Praet 2007).
Knowledge on the genetic population structure of parasites may shed light on their evolutionary history and aid in epidemiological studies (Campbell et al. 2006, Anantaphruti et al. 2013). For example, Knapp et al. (2010) used microsatellite markers to study the genetic variability of Echinococcus multilocularis in Europe and found higher genetic diversity in the historical endemic zone compared with peripheral zones. Peripheral zones were each dominated by a certain genotype, which can be explained by founder events. Furthermore, understanding the population variants of the parasite helps to analyze transmission patterns (Pajuelo et al. 2017). As tapeworms are capable of self-fertilization, most offspring cysts have the same genotype as the parental tapeworm. Thus, Pajuelo et al. (2017) were able to trace T. solium infections in pigs to human tapeworm carriers, which may ultimately facilitate control of this parasitosis. Furthermore, previous studies in T. solium have shown that infectivity and pathogenicity of tapeworms may differ among genetic variants (Campbell et al. 2006). Finally, knowledge on the genetic structure within and between populations is informative with regard to potential future evolutionary change (Campbell et al. 2006).
To gain information on intraspecific genetic variability, suitable molecular markers need to be identified. For example, microsatellite markers have been used to study population genetics and trace transmission pathways of T. solium and E. multilocularis (Knapp et al. 2010, Pajuelo et al. 2017). However, microsatellite markers have not been used in studies on T. saginata to date. In addition to microsatellites, mitochondrial gene sequences provide outstanding molecular markers to generate deep-branch taxonomic phylogenies (Boore et al. 1995, Boore 1999), as they evolve at a faster rate than nuclear genes and mutation and gene rearrangement is normally uncomplicated and taxon-specific (Wolstenholme 1992, Boore 1999). Previous studies have investigated genetic variation in taeniids based on the mitochondrial genes cytochrome c oxidase subunit 1 (cox1) (Okamoto et al. 1995, de Queiroz and Alkire 1998), cytochrome b and NADH dehydrogenase subunit 1 (Bowles and McManus 1993, Gasser et al. 1999).
cox1 sequences are relatively well preserved among taeniids. On an interspecific level, the cox1 gene showed 2.5–18% nucleotide differences among nine Taenia spp. (Gasser et al. 1999). Intraspecific analyses revealed sequence variation of 0.3–9.5% within T. taeniaeformis isolates (Okamoto et al. 1995). T. saginata isolates from Asia, South America, and Africa displayed 2.5% nucleotide variation in the cox1 gene, while only minor cox1 sequence variation was detected among T. asiatica specimens (Anantaphruti et al. 2013). In contrast, the NADH dehydrogenase subunit 5 (nad5) has been identified as one of the most variable mitochondrial genes on an inter- as well as intraspecific level, showing up to 18.5% nucleotide variation among T. taeniaeformis isolates (Jeon et al. 2007, Jia et al. 2012).
Since genetic information on European T. saginata isolates is rare, this study aimed to assess the intraspecific variation and population genetics of Cysticercus bovis isolates obtained from cattle in Northern Germany by sequence comparison of the cox1 and nad5 genes. Furthermore, obtained sequences were analyzed in relationship to T. saginata isolates from other European, South American, Asian, and African countries.
Materials and Methods
Parasite sampling and molecular investigation
One hundred twenty-eight T. saginata cysts were collected from cattle slaughtered at an abattoir in Northern Germany. Cysts or cyst-containing tissue was collected during routine meat inspection procedures. Parasite materials were kept frozen at −20°C or refrigerated at 4°C for further investigation. Cysts were extracted from tissue and homogenized using the Precellys® 24 (Peqlab Biotechnologie, Germany) by two cycles of 60 s at 6500 rpm. DNA was isolated using the NucleoSpin® 96 Tissue kit (Macherey-Nagel, Germany), following the manufacturer's instructions. In addition, 16 T. saginata cysts collected from different animals at an abattoir in Switzerland (cyst DNA was kindly provided by Ramon Eichenberger, Institute of Parasitology, University of Zurich) and 4 cysts from Belgium (cysts were kindly provided by Pierre Dorny, Department of Biomedical Sciences, Institute of Tropical Medicine, Antwerp). Belgian cysts originated from experimentally infected cattle and were stored in ethanol before DNA extraction. Apart from that, Belgian cysts were treated in the same way as German cysts. Furthermore, one proglottid from an adult T. saginata specimen from Palestine was included, which had been donated from a human patient who had been diagnosed with and treated against taeniosis. After isolation from the feces, the proglottid was washed with phosphate-buffered saline and stored at −18°C before DNA extraction as described for the T. saginata cysts.
PCR amplification of the cox1 gene was performed using primers T1F (5′-ATATTTACTTTAGATCATAAGCGG-3′) and T1R (5′-ACGAGAAAATATATTAGTCATAAA-3′) (Cho et al. 2014). For amplification of the nad5 gene, primers T.s. nad5 for (5′-TTGGTTTTCTATGCGTTATGATTA-3′) and T.s. nad5 rev (5′-CACACCCAAACAAACTACTACTCC-3′) were designed with the PrimerSelect software (DNASTAR, version 5.06; GATC Biotech, Germany). Thermocycling conditions were as follows: 95°C for 3 min, followed by 40 cycles of denaturation at 95°C for 60 s, annealing at 50°C for 60 s, extension at 72°C for 60 s, and final extension at 72°C for 10 min. Amplicons were visualized on a 1% agarose gel and PCR products showing distinct bands at 491 bp (cox1) or 749 bp (nad5) were sequenced by a customized sequencing service (Seqlab Sequence Laboratories, Göttingen, Germany).
Population genetic analyses
Reference sequences were obtained from GenBank (accession numbers provided in Figs. 1 and 2), while multiple sequence alignment (MUSCLE) and phylogenetic as well as molecular evolutionary analysis were conducted using MEGA version 7.0 (Kumar et al. 2016). Phylogenetic classification was achieved using the neighbor-joining method. The Bootstrap consensus tree was inferred from 1000 replicates. Genetic diversity values, including polymorphic sites between populations (S), haplotype numbers (h), haplotype diversity (Hd), nucleotide diversity (π), theta-ω, and theta-π as estimators to measure DNA polymorphism, were calculated using DnaSP version 5.10 (Rozas et al. 2003). The genetic structure of the parasite was elucidated under the population expansion effect via Tajima's D test, Fu's and Li's D* and F* statistics, and Fu's Fs test. All these statistics are used to test the null hypothesis of neutral mutation. Tajima's D is calculated by comparing the mean pairwise differences between two sequences and the number of segregating sites (Tajima 1989). Under the null hypothesis of neutral mutation, D does not differ significantly from 0, while a significantly negative value indicates positive selection or population expansion, and a significantly positive value indicates balancing selection or a sudden population contraction. Fu's and Li's D* and F* are based on the number of recent versus ancestral mutations (Fu and Li 1993), while Fu's Fs test estimates the probability of observing the number of alleles in question under the observed level of diversity given neutral mutation (Fu 1997). Here too, negative values indicate population expansion.
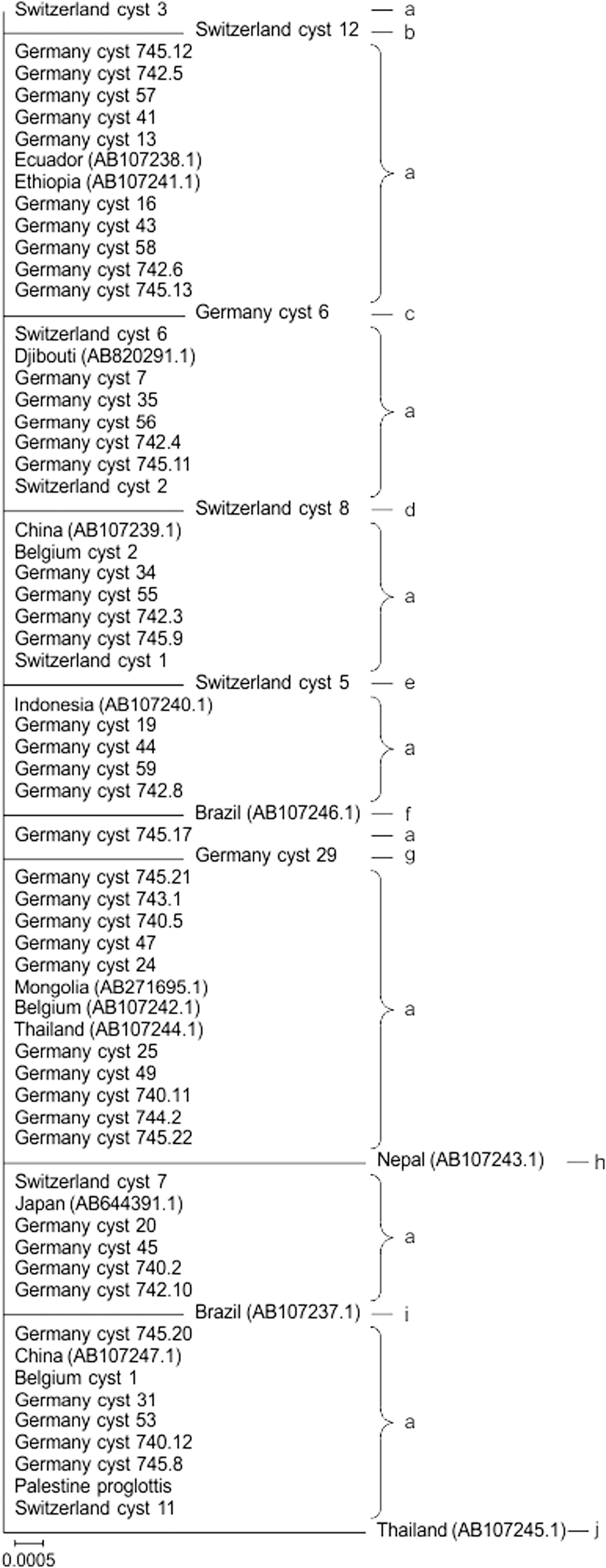
Phylogram of the partial cox1 gene of Taenia saginata from different countries. Lower case letters indicate haplotypes. cox1, cytochrome c oxidase subunit 1.

Phylogram of the partial nad5 gene of T. saginata isolates from Germany, Switzerland, and Palestine in comparison to an African sequence used as reference. Lower case letters indicate haplotypes. nad5, NADH dehydrogenase subunit 5.
Results
Parasitic material, PCR, and sequencing
Quality of T. saginata cysts varied considerably from viable cysts (clear and defined cysts) to degenerative cysts (degraded and surrounded by connective tissue, sometimes filled with purulent exudate). Each cyst originated from a different animal, with the exception of cattle no. 740, 742, and 745, which contributed multiple cysts (four, six, and nine cysts, respectively) to the sequence analysis. Cysts removed from the same cattle got the same first number, followed by a dot and an individual number (e.g., 745.8). Good-quality nucleotide sequences could not be obtained from all cysts, most likely due to DNA degradation in dead and calcified cysts. From 149 collected samples, 57 cox1 sequences (45 German, 9 Swiss, 2 Belgian, and 1 Palestinian sequence) were obtained. From 31 of these 57 samples, nad5 sequences were obtained as well (21 German, 9 Swiss and 1 Palestinian sequence). For 26 cysts (24 German and 2 Belgian cysts), only cox1 sequences were available, whereas from one Swiss cyst only a nad5 sequence could be generated.
Genetic characterization of German T. saginata isolates
Alignment of the 45 cox1 and 21 nad5 sequences from Germany revealed two polymorphic sites and mutations and three haplotypes in both genes (Table 1). Concerning cox1 sequence analysis, one dominant haplotype was found, comprising 38 of the 45 sequences, while only four sequences were assigned to the second, and three sequences to the third haplotype. All four isolates analyzed from cattle no. 740, five of six isolates from cattle no. 742, and all nine isolates from cattle no. 745 clustered together in the same haplotype. Among nad5 sequence isolates, one dominant haplotype was found, comprising 19 isolates, while the other two haplotypes comprised only one cyst each (cyst 57 and 740.12, respectively). All isolates analyzed from cattle no. 745 and 742 clustered together in the dominant haplotype.
without an outgroup; §statistically significant (p ≤ 0.05).
Haplotype diversity of German T. saginata isolates was low for cox1 (0.088 ± 0.057), while it was higher for nad5 (0.186 ± 0.110). Low nucleotide diversities were detected for both genes (cox1: 0.00028 ± 0.00018; nad5: 0.00095 ± 0.00058). Pairwise (p) distance among partial cox1 sequences ranged from 0.000 to 0.007 (overall mean distance: 0.001). Among partial nad5 sequences, p-distance ranged from 0.000 to 0.017 (overall mean distance: 0.007). Although Tajima's D neutral indexes were negative for both investigated genes, no statistical significance was found (p > 0.1).
Genetic diversity of T. saginata isolates of worldwide origin
Sequence comparison of the 57 cox1 sequences obtained in this study with 14 selected cox1 sequences from different geographical areas (source: National Center for Biotechnology Information [NCBI] GenBank) resulted in 11 polymorphic sites and mutations. A total of 10 haplotypes with high haplotype (0.239 ± 0.068) and low nucleotide (0.00097 ± 0.00031) diversity were identified (Table 1). The largest cluster consisted of isolates originating from Europe, Asia, Africa, and South America, showing no correlation of T. saginata isolates to geographical origin (Fig. 1).
As only one T. saginata nad5 sequence was available in NCBI GenBank at the time of analysis, nad5 sequence comparison included only 33 sequences, namely the 32 sequences obtained in this study and one further African sequence (Jeon et al. 2007). nad5 sequence comparison revealed 11 polymorphic sites, 12 mutation sites, and 7 haplotypes (Fig. 2, letter a–g), with high haplotype (0.335 ± 0.106) and low nucleotide (0.00391 ± 0.00163) diversity. Similar to cox1 sequence analyses, no geographic structure was found, as the largest haplotype group was composed of German, Swiss, Palestinian, and African isolates.
p-Distance among partial cox1 sequences ranged from 0.000 to 0.012 (overall mean distance: 0.001). Among partial nad5 sequences, p-distance ranged from 0.000 to 0.030 (overall mean distance: 0.010). Tajima's D neutral indexes were significantly negative (p < 0.01) for both investigated genes, indicating population expansion.
Discussion
Taeniid tapeworms are of medical and veterinary significance worldwide (Craig and Pawlowski 2002, Hoberg 2002). Although T. saginata only causes mild disease in humans, presence of the metacestode in cattle has a considerable economic impact (Gonzalez et al. 2000, Yamane et al. 2013). Knowledge on the genetic population structure may help to analyze parasite transmission patterns on different geographic scales, and may ultimately aid in improving control measures (Campbell et al. 2006, Anantaphruti et al. 2013), as demonstrated, for example, by studies on E. multilocularis (Knapp et al. 2010) and T. solium (Pajuelo et al. 2017). Studies on the genetic population structure of T. saginata have been conducted in Asia and South America based on mitochondrial genes (Anantaphruti et al. 2013, Solano et al. 2016, and Sanpool et al. 2017), however, no information on genetic variability of European T. saginata isolates has been available to date. Therefore, the present study aimed to characterize the genetic population structure of T. saginata cysts from Northern Germany based on the mitochondrial cox1 and nad5 genes, in relationship to isolates of worldwide origin.
Although seroprevalence of T. saginata cysticercosis in cattle from Germany (Federal state of Lower Saxony) was estimated at 8.83% (Abuseir et al. 2010), collection of cysts is difficult due to low detection rates during meat inspection. Furthermore, cyst quality showed considerable variation, which affected sequencing success due to degradation of DNA in nonviable cysts. Consequently, only 38.3% of the 149 collected cysts yielded high-quality cox1 sequences and only 20.8% nad5 sequences.
Comparison of the 45 cox1 sequences from Germany revealed two polymorphic sites and three haplotypes, with low haplotype diversity (0.088). In contrast, other studies on the genetic divergence of T. saginata in Asia have revealed considerably higher haplotype diversity in the cox1 gene (Anantaphruti et al. 2013, Sanpool et al. 2017). The comparably lower intraspecific diversity inferred from partial cox1 sequences in this study may result from different sequence lengths. In the present study, partial sequences of 504 bp length were used, compared with the 924 bp partial sequences analyzed by Anantaphruti et al. (2013) and the complete cox1 sequences (1620 bp) analyzed by Sanpool et al. (2017).
While the cox1 gene shows a relatively high degree of conservation, nad5 was reported to be one of the most variable mitochondrial genes in taeniids (Jeon et al. 2007). Higher haplotype diversity was also found amongst nad5 than cox1 sequences in the present study, both amongst the German isolates and in the analysis comprising isolates from other geographical areas, despite the fact that a lower number of nad5 than cox1 sequences, with a more limited geographical distribution, was available. This suggests that the nad5 gene provides a finer resolution and may be particularly suitable to study intraspecific population structure of taeniids. However, it was more difficult to obtain nad5 sequences than cox1 sequences, probably due to DNA degradation in nonviable cysts. Thus, successful generation of nad5 sequences may be dependent on viable or dead but clear cysts.
Notably, with one exception, all cysts originating from the same individual cattle always clustered together in the same haplotype. Although the sample size was rather low as only three cattle contributed multiple cysts, this finding might indicate that these cattle acquired infection mainly by direct transmission, for example, via presence of a carrier on the farm, rather than by more indirect transmission via surface water-contaminated environment. Furthermore, it can be assumed that the cysts originated from a single or short-term infection event, as on the one hand, the intermediate hosts rapidly acquire protective immunity upon first exposure to taeniids (Lightowlers 2010). Thus, when immune cattle are reexposed, invading oncospheres are killed rapidly, preventing establishment of superinfections. On the other hand, exposure to T. saginata eggs may occur only sporadically in Europe, so that most animals only encounter an infection once in their lifetime, with increasing risk of exposure with the age of animals (Dorny and Praet 2007, Dupuy et al. 2014).
Both among the German and among the global sequences, haplotype distribution was not related to geographical origin, neither regarding the cox1 nor the nad5 gene. Similarly, comparison of T. saginata cox1 sequences from Thailand (Anantaphruti et al. 2013) as well as sequences of diverse global origin (Solano et al. 2016) revealed mixed phylogeny among localities. This indicates gene flow both at a regional and at a global scale, possibly mediated by cattle trade. Likewise, populations of E. granulosus, which also infect cattle as well as other ungulates as intermediate hosts, show little genetic differentiation between localities (Casulli et al. 2012, Sharma et al. 2013, Yan et al. 2013). In contrast, geographic population structure is evident among cox1 sequences of T. solium, the pork tapeworm (Solano et al. 2016), possibly because pig trade is more restricted.
Significantly, negative Tajima's D, Fu's and Li's D* and F* indexes, as well as low nucleotide but high haplotype diversity for both the cox1 and nad5 sequences of worldwide origin indicate population expansion of T. saginata, which supports previous findings (Anantaphruti et al. 2013, Sanpool et al. 2017). Evidence of recent population expansion on a local as well as global level was also found for E. granulosus (Casulli et al. 2012, Sharma et al. 2013, Yan et al. 2013). In this study, no significant divergence from neutral evolution could be found for the analyzed German samples; however, this might be due to the smaller sample size. It has been speculated that the underlying cause for the population expansion may be anthropogenic introduction of infected hosts, such as cattle, sheep and dogs, into new geographic regions, following their domestication (Nakao et al. 2010, Sharma et al. 2013). For E. granulosus, the pattern of haplotype distribution among continents seems to reflect the spreading of livestock domestication (Casulli et al. 2012, Sharma et al. 2013). More extensive sampling in diverse geographical regions is necessary to investigate whether a similar pattern can also be observed for T. saginata.
Conclusions
Analysis of partial cox1 and nad5 sequences from T. saginata cysts obtained from German cattle revealed the presence of three haplotypes in each gene. Haplotype diversity was higher among nad5 than cox1 sequences, indicating that nad5 may prove especially useful for the analysis of genetic population structure in taeniids. However, high-quality nad5 sequences may be more difficult to obtain from cysticerci than cox1 sequences. Alternatively, complete cox1 sequences rather than partial sequences, or microsatellite markers may be more informative. When sequences from other geographical origins were included in the analyses, population expansion of T. saginata was detected, confirming the results of previous studies.
Footnotes
Acknowledgments
The authors thank Sandra Buschbaum, Marie-Kristin Raulf, and Annika Timme for technical assistance. T. saginata-DNA from Switzerland was kindly provided by Ramon Eichenberger of the Institute of Parasitology, University of Zurich. The authors are thankful to Sarah Gabriël (Department of Veterinary Public Health and Food Safety, Faculty of Veterinary Medicine, Ghent University) and Pierre Dorny (Department of Biomedical Sciences, Institute of Tropical Medicine, Antwerp) for supplying T. saginata cysts from Belgium.
Author Disclosure Statement
No conflicting financial interests exist.