
Abstract
Cryptosporidiosis is a parasitic zoonosis implicated in severe diarrhoea in pets and humans. This study aimed to determine the prevalence and genotypes of Cryptosporidium spp. in household dogs and in-contact children, and the risk factors associated with infection in children in Sharkia Province, Egypt. Fecal samples of 100 children (2–12 years old) and 50 dogs (3 months–1 year old) were randomly collected from both rural (children: n = 85, dogs: n = 40) and urban (children: n = 15, dogs: n = 10) households. Initial parasite detection was done by light microscopy, while, genotyping was based on molecular diagnostic assays. The overall prevalence of Cryptosporidium spp. infection in children was 35% using microscopy and 14% using nested polymerase chain reaction (PCR). In dogs, it was 34% using microscopy and 24% using nested PCR. Cryptosporidium spp. from children were identified as distinct genotypes, with the predominance of human genotype I (Cryptosporidium hominis) over the zoonotic genotype II (Cryptosporidium parvum). Moreover, only zoonotic genotype II (C. parvum) was identified in dog samples. The significant risk factors associated with the prevalence of Cryptosporidium infection in children were the presence of diarrheal episodes during time of survey, improper disposal of garbage, and dog feces and contact with other livestock (p ≤ 0.05). This study concluded that the existence of C. parvum in children and dogs residing the same households confirm the zoonotic transmission and its public health significance. Also, the study recommended the necessity of hygienic disposal of dog feces and preventing direct contact of dogs with other livestock.
Introduction
C
In Egypt, cryptosporidiosis is common in children and has been known as a primary agent of childhood diarrhea (Antonios et al. 2001, Youssef et al. 2008). The infection can be transmitted to human through close contact with dogs (Hunter and Thompson 2005). Cryptosporidium canis and C. parvum can infect both dogs and humans (Fayer et al. 2000, Xiao and Fayer 2008).
The Cryptosporidium oocyst wall protein (COWP) is one of the most target genes used for molecular identification of Cryptosporidium spp.; and differentiating C. parvum genotypes from other Cryptosporidium spp. on the basis of polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) genotyping (Spano et al. 1997, Sulaiman et al. 1999). Of interest, the utilization of COWP primers was sensitive enough to carry out a routine detection study (Kato et al. 2003). Also, nested-PCR of COWP gene could be a reliable method to identify natural infection of animals with Cryptosporidium and its genetic diversity (Kato et al. 2003). Genotyping of Cryptosporidium allows species identification and elucidates the epidemiology of cryptosporidiosis (Hajdusek et al. 2004, Abe et al. 2006).
Humans may be infected with Cryptosporidium hominis (anthroponotic genotype), C. parvum (zoonotic or bovine genotype), and C. canis (dog genotype) (Abe et al. 2006, Ryan et al. 2014). Moreover, dogs are infected with the host-adapted genotype (C. canis) and the zoonotic genotype (C. parvum) (Hajdusek et al. 2004). In Egypt, so far there is no available literature on genotyping of Cryptosporidium spp. isolated from household dogs and in-contact children and the risk factors associated with zoonotic transmission. This study was done to determine the prevalence and genotypes of Cryptosporidium spp. in household dogs and in-contact children. In addition, the risk factors associated with human cryptosporidiosis were assessed.
Materials and Methods
Ethical statement
The study was approved by Institutional Animal Care and Use Committee, Zagazig University (ZU-IACUC) under the following number:ZU-IACUC/2/F/10/2018. Fecal samples of dogs were only collected after spontaneous defecation upon an informed consent of their owners. The medical ethics were in accordance with the declaration of Helsinki. Informed and written consent was obtained from the parents of the children involved. Several meetings were carried out to explain the aim and protocol of this study to their parents, and to emphasize that participations were voluntary and the withdrawal from this study at any time was permitted without any reason.
Sampling
Fecal specimens of 100 children (2–12 years old) and 50 dogs (3 months–1 year old) were randomly collected from both rural (children: n = 85, dogs: n = 40) and urban (children: n = 15, dogs: n = 10) households at Sharkia Province, Egypt. The majority of householders in rural areas and few in urban ones are rearing cattle, sheep, and dogs on small-holder farms in close vicinity to homesteads. Householders were asked to collect fresh fecal samples immediately after defecation from their own dogs and complete questionnaire sheet with data regarding age, sex, health status, and history of medication for their children and dogs.
Direct smear method
Fecal sample (2 mg) was emulsified in a drop of saline (0.85%) in the center of a clean glass slide until the contents became homogenous. The field was covered with a coverslip and examined under low power ( × 10 objective) of a light microscope.
Formalin-ether sedimentation method
Cryptosporidium spp. Oocysts were concentrated in each of the fecal samples using formalin-ether sedimentation technique (Cheesbrough 2005).
Staining techniques
Two fecal smears were prepared from each of the positive sample using formalin-ether sedimentation technique. One smear was stained by the modified Ziehl-Neelsen stain (MZN) (Dubey et al. 1990) and the other one was stained with dimethyl sulfoxide-modified acid fast stain (Bronsdon 1984). The stained smear was then microscopically examined for the presence of Cryptosporidium oocysts under the oil immersion lense ( × 100 objective lense).
Sheather's sugar floatation technique
The technique was used to concentrate Cryptosporidium oocysts for molecular typing. Each positive fecal sample (2 gm) was homogenized in 10 mL of modified Sheather's sugar solution (specific gravity = 1.27) and then strained into a centrifuge tube through a tea strainer. The centrifuge tube with the filtrate was filled with the floatation solution and centrifuged at 1200 rpm for 5 min (Dryden et al. 2005). A drop of the float/supernatant was stained with MZN to confirm the presence of Cryptosporidium spp.oocysts. The oocyst density was graded according to Hamnes et al. (2007) into low 1+ (<5 oocysts per slide), mild 2+ (5–10 oocysts per slide), and high 3+ (>10 oocysts per slide). The oocyst-positive supernatant was resuspended in two volumes of phosphate-buffered saline (PBS) and then centrifuged at 1500 rpm for 5 min. The supernatant was discarded and oocyst pellet was resuspended in PBS to a volume of 1 mL and stored at −20°C until the molecular identification.
Molecular identification
Cryptosporidium spp. were molecularly confirmed in all purified samples by Sheather's sugar floatation using nested PCR targeting COWP gene (Pedraza-Díaz et al. 2001). DNA was extracted using FavorPrep™ stool DNA isolation Mini Kit (Favorgen Biotech corporation ping-Tung 908, Taiwan). The primers were BCOWPF: (5′-ACCGCTTCTCAACAACCATCTTGTCCTC-3′), BCOWPR: (5′-CGCACCTGTTCCCACTCAATGTAAACCC-3′) in the primary reaction and Cry-15: (5′-GTAGATAATGGAAGAGATTGTG-3′), Cry-9: (5′-GGACTGAAATACAGGCATTATCTTG-3′) in secondary reaction. The conditions for both PCR reactions included an initial denaturation at 95°C for 5 min, followed by 35 cycles of denaturation at 94°C for 30 s, annealing at 55°C for 30 s, and extension at 72°C for 30 s with a final extension at 72°C for 10 min. PCR products and 100 bp DNA Ladder (Fermentas, Cat#SM0371) were run on 2% agarose gel and then visualized on a UV transilluminator. Positive control C. parvum genotype from human was kindly provided by Department of Medical Parasitology, Kasr Al-Ainy, Faculty of Medicine, Cairo University.
Cryptosporidium genotyping by PCR-RFLP
The nested PCR products of Cryptosporidium spp. COWP gene and C. parvum positive control were digested with RsaI restriction enzyme (Fermentas, Lithuania) according to the manufacturer's instructions (Spano et al. 1997). The digested PCR products were analyzed by electrophoresis on 2% agarose gel and visualized on a UV transilluminator.
Statistical analysis
Differences in proportions between variables were estimated using Chi-square test. Risk factors related with cryptosporidiosis in the investigated children were determined by Univariate logistic regression model. Data analysis was performed by SPSS version 22 for windows (IBM Corp. 2013, Armonk, NY) to compute p values for pearson chi-square and odds ratio (OR). A statistical significance was considered when the p value ≤0.05. Level of agreement between diagnostic techniques was estimated using Kappa (κ) test. κ-values <0.75 indicate an excellent agreement and those >0.40 represent a poor agreement (Altman 1991).
Results
The frequency distribution of Cryptosporidium spp. in 100 stool samples of children was 35% by microscopy and 14% by nested PCR. The infection rate was significantly higher (p ≤ 0.05) in diarrheic versus nondiarrheic children (Table 1). The overall frequency of Cryptosporidium spp. in household dogs using microscopy and PCR was 34% (17/50) and 24% (12/50), respectively. The percentage of infection was significantly higher in diarrheic than nondiarrheic dogs (p ≤ 0.05). By microscopic analysis, the infection rate with Cryptosporidium spp. was significantly higher in dogs ≤6 months compared with dogs >6 months–1 year of age (p = 0.003). However, the frequency of Cryptosporidium spp. was higher in male dogs than female and in rural than urban ones, with no statistically significant difference in both comparisons (p > 0.05) (Table 2). The oocyst density was low in all positive samples after concentration. A statistically significant difference existed between microscopic analysis and nested PCR method (p < 0.001), and a poor agreement found between them (κ-value = 0.173).
Significant at p ≤ 0.05.
PCR, polymerase chain reaction.
Significant at p ≤ 0.05.
PCR-RFLP genotyping of the nested PCR positive human samples (n = 14) revealed the existence of human genotype I (C. hominis; n = 7), zoonotic genotype II (C. parvum; n = 4), and mixed genotype I + II(n = 3). Genotyping of nested PCR positive dog samples (n = 12) showed the presence of only genotype II (C. parvum) in five samples and the restriction fragments banding patterns of this genotype were identical to C. parvum genotype in the children (Table 3, Fig. 1). Meanwhile, the other seven samples of dogs showed no digestion pattern. Among the risk factors analyzed by univariate logistic regression model, prevalence of Cryptosporidium infection in household children was significantly associated with the presence of diarrheal episodes during time of survey, improper disposal of garbage and dog feces, and contact with other livestock (p ≤ 0.05) (Table 4). The other risk factors showed no significant relevance (p > 0.05).
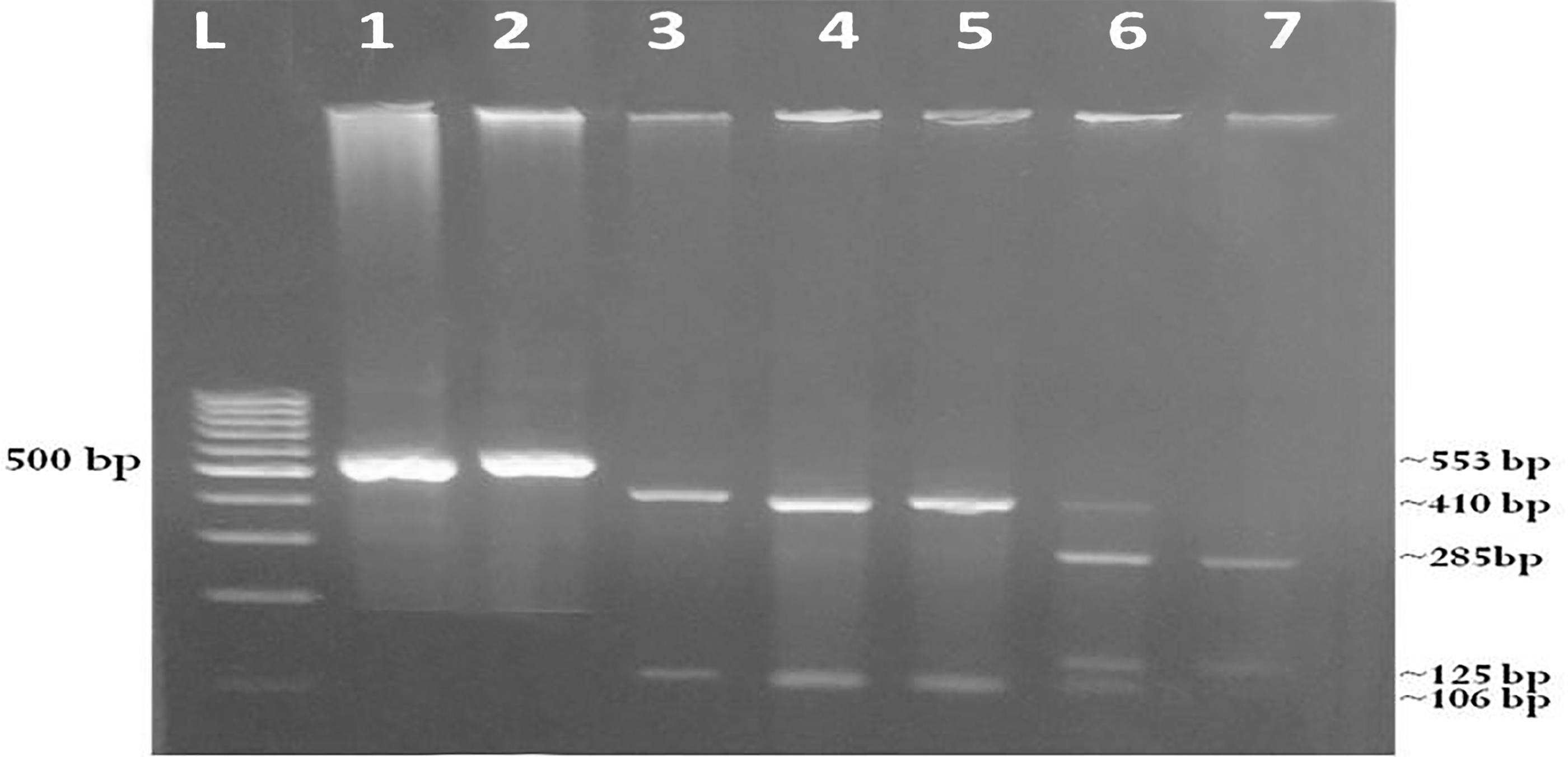
Agarose gel electerophoresis of nested-PCR products of COWP gene and PCR-RFLP patterns for Cryptosporidium genotypes after digestion with restriction enzyme RsaI. Lane L: DNA ladder (100 bp). Lanes 1–2: Amplified nested PCR products targeting 553 bp of Cryptosporidium spp. COWP gene. Lanes 3–4: products of genotype II (Cryptosporidium parvum) from human and dog samples (34, 106, and 410 bp). Lane 5: Positive control for genotype II (C. parvum) from human. Lane 6: PCR-RFLP products of mixed genotypes C. parvum and Cryptosporidium hominis from human sample (34, 106, 125, 285, and 410 bp). Lane 7: PCR-RFLP products of genotype I (C. hominis) from human sample (34, 106, 125, and 285 bp). The band 34 bp in all genotypes was very small, faint, and difficult to visualize. PCR, polymerase chain reaction; COWP, Cryptosporidium oocyst wall protein; PCR-RFLP, polymerase chain reaction-restriction fragment length polymorphism.
Significant at p ≤ 0.05.
Discussion
This study is the first to investigate the prevalence and genotyping of Cryptosporidium spp. among household dogs and in-contact children in Egypt. Also, this work is unique in that the risk factors associated with Cryptosporidium infection were assessed in children who had a potential contact with those pets. Cryptosporidium infection could cause diarrhea in humans and dogs. Cryptosporidium spp. in household dogs have a potential risk of zoonotic transmission (Itoh et al. 2014). This study using microscopy (35%) compared with PCR (14%) showed a higher percentage of Cryptosporidium spp. in children. Nearly similar percentage was previously recorded in Tanzania (Tellevik et al. 2015). However, a lower rate was found in rural Ghana (Eibach et al. 2015). In this study, distribution of Cryptosporidium infection showed higher significance in diarrheic children (62.5% by microscopy and 37.5% by nested PCR) than those in nondiarrheic children (26.3% by microscopy and 6.6% by nested PCR). This was inagreement with Tellevik et al. (2015); whereas the prevalence of C. parvum/hominis by multiplex real-time PCR technique was significantly higher in diarrheic cases (16.3%, than in children without diarrhea (3.1%) in Tanzania (Tellevik et al. 2015). A lower Cryptosporidium prevalence (15%) was recovered from diarrheic children in Fayoum, Egypt (El-Mohamady et al. 2006). The prevalence of Cryptosporidium infection by nested PCR method was 25.8% in diarrheic children and 16% in asymptomatic ones in Aboul-Reesh Cairo University Pediatrics Hospital, Egypt (Abdelrazek et al. 2016). By microscopy methods, lower prevalence (3.5%) in Egypt and (8.9%) Kenya were reported in diarrheic children by Elfadaly et al. (2018) and Mbae et al. (2013), repectively. However, higher prevalence (41.2%) was detected by microscopic analysis in diarrheic children at Abu Rish Hospital, Egypt (Hassanain et al. 2015). Moreover, Latif and Rossle (2015) cited different percentages of Cryptosporidium infection in diarrheic children: 8.5% in Iraq; 37.3% in Jordan, and 4.62% in Malaysia. The discrepancies in the prevalence of Cryptosporidium spp. in children in this study compared with other studies might be attributed to differences in the sensitivity of diagnostic assays; variations in sociodemographic, behavioral, and environmental factors; and the sample size (O'connor et al. 2011, Abdelrazek et al. 2016). The higher prevalence of Cryptosporidium infection observed in diarrheic children in our study might be attributed to the close contact of the investigated children with infected household dogs and other infected livestock or to the drinking of contaminated groundwater with animal excreta and human sewage. This finding was supported by Yu et al. (2004), where the contact with infected animals posed a risk for zoonotic transmission of C. parvum to farmers.
Among the examined 50 dogs, the overall prevalence of Cryptosporidium spp. was 34% by microscopy and 24% by nested PCR. Nearly similar PCR-based prevalence (19.3%) was reported in Thailand (Tangtrongsup et al. 2017). However, lower prevalence of Cryptosporidium infections were cited in dogs elsewhere: 1.7% in Egypt (Ahmed et al. 2014); 8.7% in The Netherlands (Overgaauw et al. 2009); 1.7% in Costa Rica (Scorza et al. 2011), 1.23% in Germany (Sotiriadou et al. 2013), 5.4% in Nigeria (Olabanji et al. 2016); 5.5% in Northern Spain (de Lucio et al. 2017); 7.2% in Japan (Itoh et al. 2014); and 4.1% in Northern Spain (Gil et al. 2017). On the contrary, Samie et al. (2013) found higher prevalence of Cryptosporidium spp. (41.7%) in home-based dogs in South Africa. The global prevalence of Cryptosporidium spp. in dogs could vary depending on the sensitivity of used diagnostic techniques, geographic locations, and the tested population (Lucio-Forster et al. 2010). In our study, the higher prevalence of Cryptosporidium in household dogs compared to the prevalence in previous studies might be attributed to ingestion of dog feces or cattle feces (dog coprophagia), contaminated soil, poor sanitation, and the pack behaviors of dogs that facilitate the transmission of Cryptosporidium infection through contact with infected animals. Those hypotheses were previously supported (FitzGerald et al. 2011, Samie et al. 2013, Ahmed et al. 2014.). The prevalence of Cryptosporidium was significantly higher (p ≤ 0.05) in 8 (53.3%) diarrheic dogs by using each of microscopy and nested PCR, as compared to 9 (25.7%) and 4 (11.4%) in the corresponding dogs without diarrhea. Similarly, the prevalence of Cryptosporidium spp. was significantly higher in diarrheic dogs than in nondiarrheic ones in Northeast Spain (Causapé et al. 1996) and Nigeria (Olabanji et al. 2016). On the contrary, higher and nonsignificant prevalence of Cryptosporidium was found in nondiarrheic (32.6%) than in diarrheic (23.5%) dogs in Thailand (Tangtrongsup et al. 2017). In this study, the higher infection of Cryptosporidium in diarrheic dogs was augmented by Lallo and Bondan (2006), who cited that high prevalence of cryptosporidiosis is only clinically manifested in diarrhea in cases of animal weakness.
The frequency distribution of Cryptosporidium spp. was significantly higher (p = 0.003) in 50% of younger dogs (≤6 months of age) compared to 10% in older dogs (>6 months–1 year of age), when their fecal samples were analyzed microscopically. Similarly, prevalence of Cryptosporidium was significant higher (8%) in dogs <90 days than in dogs >360 days in China (Jian et al. 2014). In France, Osman et al. (2015) recorded higher significant prevalence in dogs <1-year age (7.7%) than the age >1 year (0%). This finding might be attributed to immature immune systems of juvenile dogs, whereas these pets could be more venerable to Cryptosporidium infection (Samie et al. 2013, Gil et al. 2017). There was no significant association (p > 0.05) between prevalence of Cryptosporidium among dogs and age groups (Olabanji et al. 2016, Gil et al. 2017). There was no significant difference between prevalence of Cryptosporidium and sex of dogs in both samples examined by microscopy and PCR (p > 0.05). The distribution of Cryptosporidium infection was higher in male household dogs than females. Two previous studies conducted in South Africa and Spain found similar findings (Samie et al. 2013, Gil et al. 2017). However, other studies in Nigeria and Thailand reported higher prevalence of Cryptosporidium infection in females than males with no statistical association (Olabanji et al. 2016, Tangtrongsup et al. 2017). This referred that sex is not a major factor influencing prevalence of Cryptosporidium in dogs. The distribution of Cryptosporidium infection showed no significant increase in 16 (40%) and 11 (27.5%) dogs in the rural locality with the use of microscopy and PCR, respectively, compared to 1 (10%) in the corresponding animals in the urban one (p > 0.05). This finding coincided with the results of Adriana et al. (2010) and Olabanji et al. (2016). The high prevalence in household dogs from the rural area might be attributed to the residence of most household dogs in rural villages under poor hygienic level, and ingestion of contaminated water and food in rubbish during outdoor time. In addition, dogs live with other livestock in small farms close to households for security purposes. Therefore, dogs could acquire Cryptosporidium infection, particularly C. parvum through consumption of water contaminated with infected cattle feces or runoff from farms. This interpretation was supported by finding of Adriana et al. (2010), who reported that higher prevalence from rural areas could be correlated with the living of dogs close to farms of cattle and sheep.
Upon comparison of the two diagnostic methods, there was a significant difference between microscopy and nested PCR method (p < 0.001), and a poor agreement existed between them (κ-value = 0.173). It appeared that the higher prevalence of Cryptosporidium infection was detected by the microscopic analysis in comparison to the nested PCR technique. This study indicated that the nested PCR was less sensitive than the microscopic analysis. This finding could possibly be attributed to the low number of Cryptosporidium oocysts in positive fecal samples that yield insufficient DNA concentration to be identified by nested PCR (Abd El Kader et al. 2012, de Lucio et al. 2017). Also, one of the major limitation for sensitivity of nested PCR technique in our study was the presence of PCR inhibitors. This was in line with the finding of Wiedenmann et al. (1998). In addition, PCR inhibitors such as bilirubin, bile salts, and polysaccharides induced inhibition for the activity of Taq polymerase and blocked the columns of DNA extraction (Monteiro et al. 1997, Wells et al. 2016). In our study, PCR inhibition assay was conducted to check whether any inhibitors existed in the fecal samples that dampened the amplification using nested PCR of COWP gene as previously described (Khalil et al. 2016). Briefly, DNA of two samples, that was positive by microscopic analysis and negative by nested PCR, was first subjected to serial dilution (1:10) up to five dilutions, and then amplified using nested PCR assay. The dilution of genomic DNA resulted in dilution of inhibitors present in fecal samples. Second, all fecal samples that were negative by PCR technique including the former two microscopically positive specimens were retested for the amplification by using DNA of COWP as a positive control. The positive control DNA of COWP gene was added to DNA from all negative specimens and the amplification technique was repeated again. The amplification was detected in all specimens except the two samples that were positive by microscopy. This result confirmed presence of PCR inhibitors in these samples that prevented amplification of Cryptosporidium DNA by nested PCR. Therefore, PCR should not be used as a primary test for clinical diagnosis of Cryptosporidium spp. but should be utilized as a confirmatory method.
Cryptosporidium genotyping was carried out to understand transmission routes and epidemiology of cryptosporidiosis and to implement strategies for prevention and surveillance of the disease (Jex et al. 2008). The nested PCR products targeting COWP gene was digested with restriction enzyme Rsa I. Among 14 positive children samples, PCR-RFLP analysis revealed presence of 2 genotypes: 7(50%) had human genotype I C. hominis; 4 (28.5%) had zoonotic genotype II C. parvum, and 3 (21.4%) had mixed genotype I + II. In our study, the predominance of anthroponotic genotype (C. hominis) over the zoonotic or bovine genotype (C. parvum) in household children was in agreement with the previous studies in Egypt (Abd El Kader et al. 2012, Abdelrazek et al. 2016), Mexico (Valenzuela et al. 2014), and Tanzania (Tellevik et al. 2015). Our results indicated that household children could aquire Cryptosporidium infection through anthroponotic and zoonotic transmission. However, RFLP analysis clarified that 8/9 (88.9%) children samples from Egypt were genotype 1 (C. hominis) (Sadek, 2014).
Out of 12 positive dog samples, 5 (41.6%) had genotype II C. parvum and 7 (58.3%) had untyped genotype. Only the genotype II C. parvum (bovine genotype) was recovered from dogs; while Cryptosporidium isolates of the other seven positive dog samples did not show any banding patterns. Those untypeable genotypes were most likely related to the inability of the RsaI restriction enzyme to digest the nested PCR products of COWP gene. Of interest, C. parvum genotypes from dog samples exhibited the same banding profiles displayed by C. parvum genotypes from children samples. These findings indicated that household dogs are potential reservoirs for C. parvum (bovine genotype) with the possibility of direct or indirect zoonotic transmission to humans in close contact with dogs and other domestic livestock. Similarly, Fayer et al. (2001) reported C. parvum (cattle genotype) in a dog. Our finding was augmented by the results of Sotiriadou et al. (2013), whereas the household dogs are reservoirs for C. parvum (zoonotic genotype). C. parvum genotype was previously identified in household dogs in The Netherlands (Overgaauw et al. 2009), Costa Rica (Scorza et al. 2011), and Thailand (Tangtrongsup et al. 2017). Conversely, C. canis (canine genotype) was the only identified genotype in household dogs in Northern Spain with no evidence of zoonotic transmission of cryptosporidiosis among pet dogs and their owners in this geographic region (de Lucio et al. 2017).
In our study, several risk factors were investigated to estimate the association with prevalence of Cryptosporidium among household children. The presence of diarrheal episodes during time of survey was a significant risk factor associated with cryptosporidiosis among studied children. An OR of 8.52 (95% CI: 2.498–29.059; p = 0.000) was associated with presence of diarrheal episodes during time of survey. These results were in agreement with the findings reported in India (Sarker et al. 2014), Kenya (Helen et al. 2015), and United States (Roy et al. 2004). Our data confirmed that Cryptosporidium infection causes persistent diarrhea in children in Egypt. Different risk factors such as age, sex, residence, washing hands before drinking or eating, mother education, contact with pets, and others revealed no significant association with the prevalence of cryptosporidiosis in children (p > 0.05). Similarly, no significant associations were noticed between Cryptosporidium infection and any hygienic habits including washing hands before meal or after defecation and having a household lavatory (Yang et al. 2017). Significant association was found between dog contact or cat contact and cryptosporidiosis in Australia (Robertson et al. 2002) and in Spain (Cardona et al. 2011). In this study, Cryptosporidium infection was found to be significantly associated with sanitary disposal of garbage and dog feces with an OR of 0.151 (p = 0.044). This finding might be attributed to the improper disposal of dog feces and existence of wet and contaminated floors, which facilitate the survival of Cryptosporidium oocysts and the spread of cryptosporidiosis among dogs and children as previously supported by Abebe et al. (2008). However, there was no significant association between childhood cryptosporidiosis and presence of environmental contaminants such as garbage dumps or animal sheds (Sarkar et al. 2014). Also, significant association was observed between cryptosporidiosis and contact with other livestock with an OR of 4.32 (95% CI, p = 0.049). This finding was in agreement with previous studies carried out in Egypt (Helmy et al. 2013), Australia (Robertson et al. 2002), and United Kingdom (Lake et al. 2007). The infected livestock animals (cattle and sheep) have the potential to shed large number of Cryptosporidium oocysts, contaminate the environment and infect children who live in close proximity to these animals especially in rural areas (Casemore 1990). Those livestock became infected through contaminated water and agricultural sources (Lake et al. 2007). Our result contrasted the findings of Helen et al. (2015) and Quihui-Cota et al. (2017), where there was no significant association between cryptosporidiosis and contact with livestock.
Conclusion
This study was the first to investigate genotyping of Cryptosporidium spp. in household dogs and in-contact children in Egypt. C. hominis genotype I (anthroponotic genotype) was predominant in children over C. parvum genotype II (bovine genotype). The presence of C. parvum genotype II (bovine genotype) with identical PCR-RFLP patterns in both dogs and children confirmed that household dogs are potential reservoirs of C. parvum (bovine genotype) with the possibility of zoonotic and interspecies transmission. Improper disposal of dog feces and the contact with livestock present in the homesteads were significant risk factors associated with cryptosporidiosis in children. Therefore, a better structured molecular study and effective control measures including hygienic disposal of dog feces, disinfection of houses floor, and preventing contact with livestock, are needed to prevent transmission of Cryptosporidium spp. to vulnerable children.
Footnotes
Author Disclosure Statement
No conflicting financial interests exist.