
Abstract
Erythema migrans (EM) rashes once considered pathognomonic of Lyme disease (LD) have been reported following bites of arthropods that do not transmit LD and in areas with no LD. Also, EM rashes have been reported in association with organisms other than members of Borrelia burgdorferi sensu lato complex. Arthropod saliva has chemicals that have effects on the host and pathogen transmission. Tick saliva has protein families similar to spiders and scorpions and even substances homologous to those found in snakes and other venomous animals. Ticks “invertebrate pharmacologists” have a sophisticated arsenal of chemicals that assist in blood feeding, pathogen transmission, and suppressing host defenses. No organisms have been isolated from many EM rashes. We propose that tick salivary toxins may play a role in the causation of rashes and laboratory abnormalities in tick-borne diseases. The role of tick salivary toxins needs further exploration.
Cases of Lyme-like EM rashes referred to as STARI (Southern Tick-Associated Rash Illness) following bites of the lone star tick, Amblyomma americanum, in the United States have been reported predominantly in Southeastern Missouri and a few in South Carolina, North Carolina, Georgia, and one case each in Mississippi and Long Island, New York. Although there is one report of Borrelia lonestari in a patient with a rash, biopsies of 31 cases of STARI, with cultures and PCR, failed to show a relationship. Distribution of A. americanum, whose bites are associated with STARI, now extends along the East Coast of the United States, including New Jersey, up to the Canadian border. As far as we are aware, there have been no prior reports of Lyme-like rashes in New Jersey. In this study, we present case examples of 2 Lyme-like rashes, variations of EM rashes, and a brief review of studies that suggest a role of tick salivary toxins in tick-borne diseases.
Introduction
E
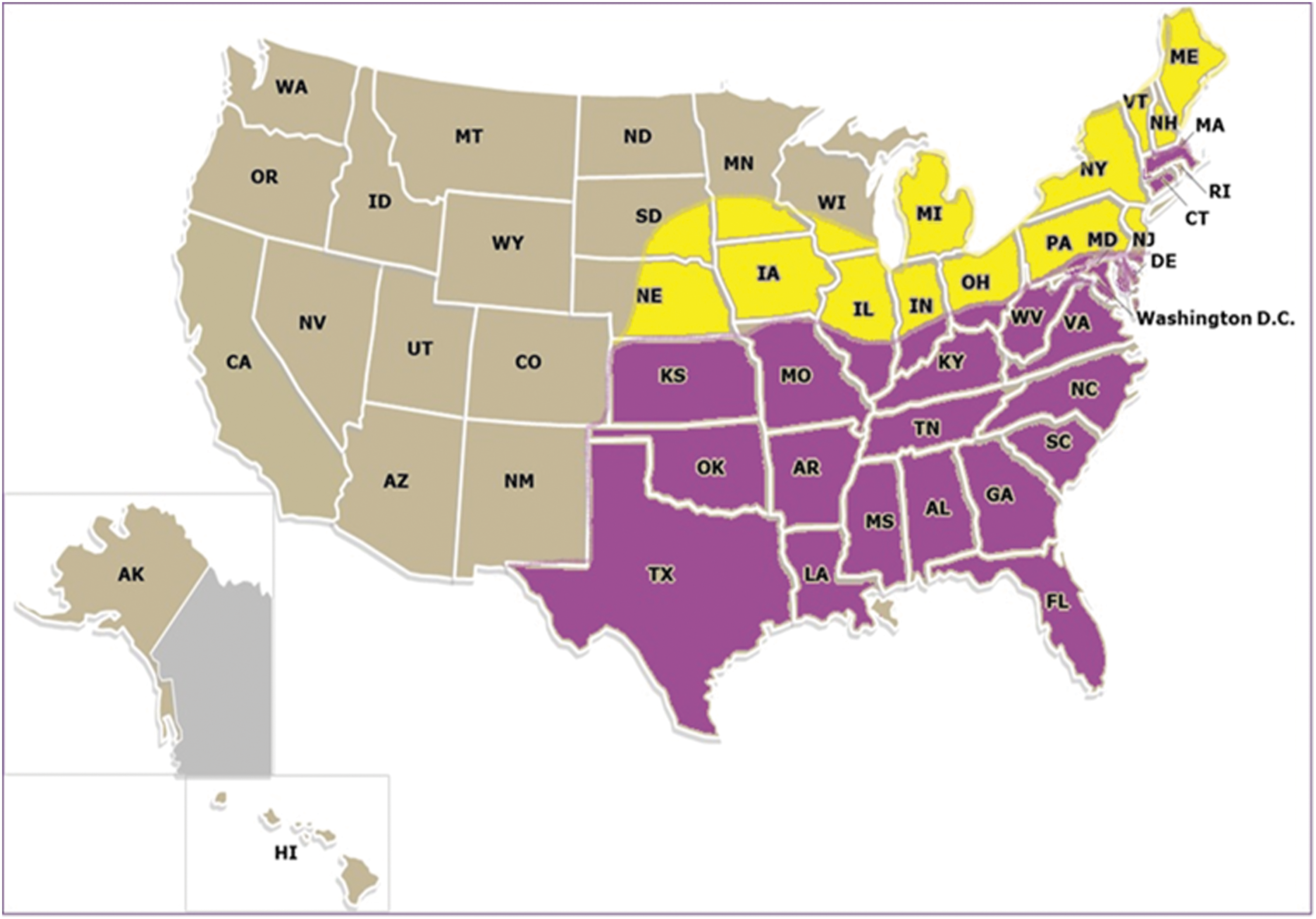
Map showing northward spread of lone star tick Amblyomma americanum since 1986.
Ixodid Tick-Associated Rashes, Where Salivary Compounds, Including Toxins, May Play a Role
SENLAT, scalp eschar and neck lymphadenopathy syndrome; DEBONEL, dermacentor-borne necrosis, erythema, and lymphadenopathy; TIBOLA, tick-borne lymphadenopathy; SFTS, severe fever with thrombocytopenia syndrome; ?, unidentified.
LD is the most common cause of EM rashes, but recent literature shows that it is not unique to LD. According to CDC 2017 guidelines, EM rashes in a state with more than 10 confirmed cases per 100,000 are diagnostic of LD. This has led to misdiagnosis of possible non-LD EM rashes. EM 2017 CDC definition is a skin lesion that typically begins as a red macule or papule and expands over a period of days to weeks to form a large round lesion, often with partial central clearing and must reach at least 5 cm across the largest diameter. This definition has obvious limitations as it does not consider many variations reported in recent literature and even does not explain five of the seven pictures of EM rashes in the CDC website (red expanding lesion with central crust and irregular borders, multiple red lesions with dusky centers, red oval shaped plaque on trunk, bluish hue without central clearing, and red blue lesion). Recently reported vesicular, bullous, hemorrhagic, and necrotic lesions (Mazori et al. 2015, Paul et al. 2016) are not included. We are presenting pictures of rashes with varying areas of clearance, varying sizes, borders and shapes, varying locations, single and multiple rashes, purplish blistering lesions, vesiculopustular lesions mimicking poison ivy rashes, and eschar-associated EM (Figs. 2 and 3).

Non-Lyme rashes: case1 and case2.
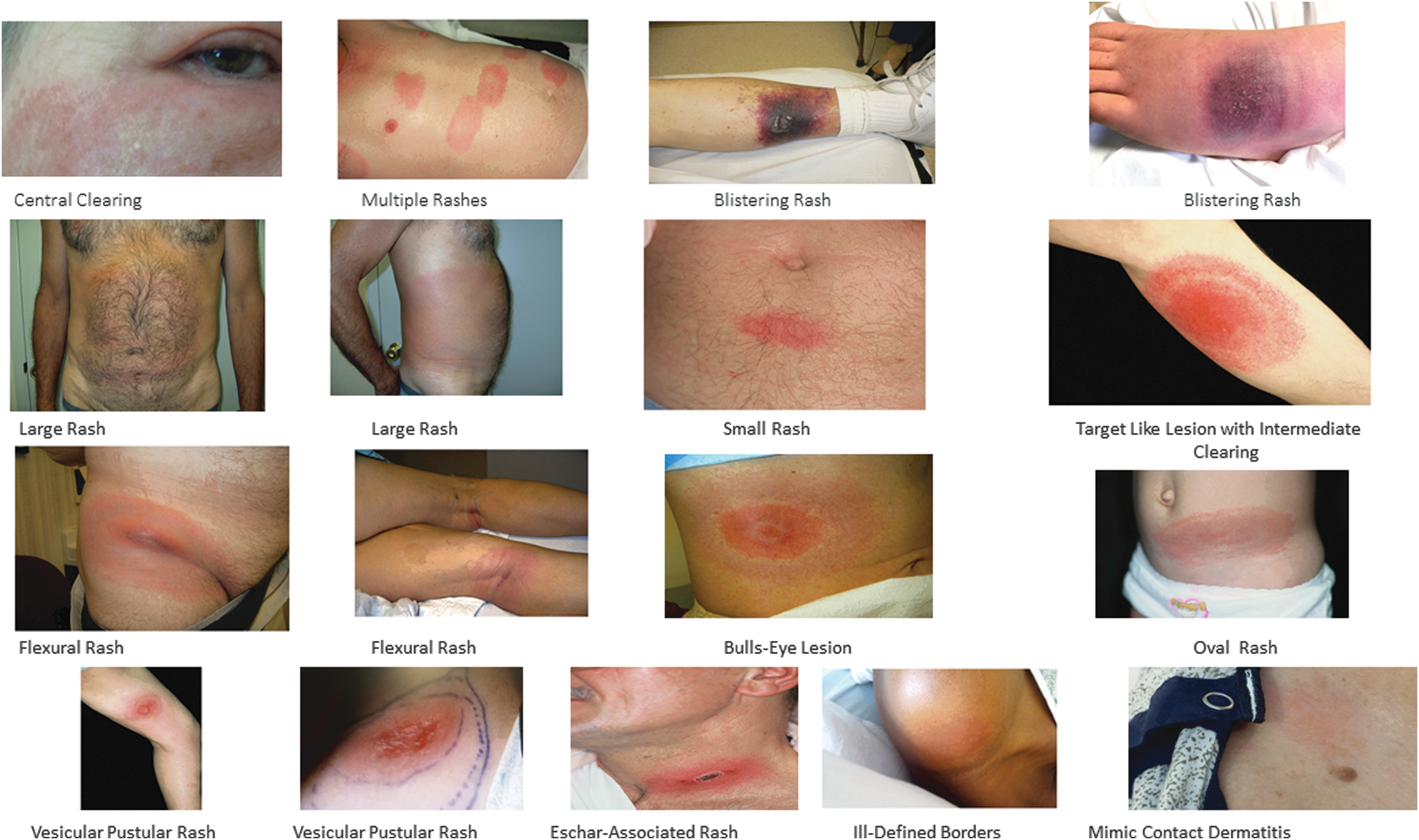
Variations in Erythema migrans rashes.
Candidatus Neoehrlichia mikurensis (CNm) patients in Southern Norway presented with EM, as the only symptom (Quarsten et al. 2017). CNm was found in a surprisingly large fraction of tick bitten persons (diagnosed by PCR). Most EM patients were negative for all pathogens tested. Previously a case of CNm was reported (Li et al. 2012) in China with multiple oval areas of erythema in neck. The tick species reported in Norway study area was I. ricinus. EM-like rashes were reported in Caribbean Islanders bitten by ticks in areas with no LD (Sharma et al. 2010). EM has been found in association with a variety of bites, for example, Dermacentor marginatus associated with Candidatus Rickettsia rioja (Oteo et al. 2004), A. americanum associated with Rickettsia amblyommii (Billeter et al. 2007), and “spotless RMSF” due to Rickettsia rickettsii (Hughes 1995). There are two reports of “erysipelas like” lesions in cases of tick bite fever, caused by R. africae following Amblyomma variegatum bites (Bouvresse et al. 2008). A recent fatal case of deer tick virus in a patient bitten by ticks (Ixodes scapularis) 1 month and 7 days before admission presented with EM, but the Lyme test failed to seroconvert (Cavanaugh et al. 2017). This patient died 10 days after admission.
EM seems to correlate more with the arthropod than the associated pathogen (Table 1). The common belief is that EM is caused by B. burgdorferi. The presence of an organism in a skin biopsy culture or PCR does not necessarily mean that it is the cause of EM. The majority are associated with I. ricinus complex (I. scapularis, Ixodes pacificus, I. ricinus and Ixodes persulcatus). Few have been reported with Amblyomma spp. and rare reports due to Dermacentor spp. EM-like rashes have not been reported with other spirochetes that cause human infections namely Leptospira and Treponema or relapsing fever causing Borrelia transmitted by soft ticks. EM-like rashes have been reported in association with African trypanosomiasis (Ezzedine et al. 2007) caused by Trypanosoma gambiense transmitted by tsetse flies. Tsetse fly saliva is known to contain at least 20 pharmacologically active compounds (Awuoche 2012). There are reports of lesions resembling brown recluse spider bites in areas where they are absent (Vetter et al. 2003). LD has been misdiagnosed as lesions caused by brown recluse spider bites (Rosenstein and Kramer 1987, Osterhoud et al. 2002). Tick bite alopecia and eschars (Table 1) likely due to tissue damage from tick toxins have been attributed by some to an organism, rather than a mere association. It is likely that tick salivary toxins play a role in EM, eschars, and tick-bite alopecia (Castelli et al. 2008). Alopecia has been reported following ant, flea, bee, and tick bites (Mortazavi 2003, Lynch et al. 2016, Sperling et al. 2016). Recently reported “Renbök phenomenon” (Dumas et al. 2017) where a patient who developed a generalized allergic skin rash had sparing of a previous site of EM suggests destruction of cells mediating the allergic skin response by lytic properties of tick saliva (Simo et al. 2017). This phenomenon has been reported in an area of skin damaged by radiation (Cochran and Wilkin 1981).
There are reports of EM worsening while on recommended LD treatment (Feder et al. 2011, and two of our cases) and EM developing in seven patients after adequate treatment, following a tick bite (Maraspin et al. 2002). These rashes are likely non-Lyme. Leukopenia and thrombocytopenia are common in tick-borne infections (Table 3). Many unrelated tick-borne infections share similar hematologic and blood chemistry abnormalities (Table 3), suggesting factors other than the pathogen, may play a role in the etiology. In this study, we present 2 cases of non-Lyme rashes that did not respond to standard LD treatment and examples of a variety of atypical EM rashes (Figs. 2 and 3).
Case Reports
Case 1. A 27-year-old female hair dresser, with no medical history presented to the emergency room with a history of fever, body aches, and a circular patch of localized redness and swelling on the lower lateral right thigh (Fig. 2). Approximately 5 days earlier, she noticed a small bite-like lesion, which was painful and pruritic, on the lower lateral right thigh. The patient used over-the-counter Benadryl (diphenhydramine) cream which helped her pruritus, but the area of redness became progressively larger. She gave a history of sitting on a bench outside in a tick-infested area. She was seen at an urgent care center with a 2-day history of body aches, fever of 102.6 F, and chills. She was prescribed amoxicillin and Bactrim and took the medications as directed. Five days after noticing the bite, she arrived at the emergency room with fever, chills, body aches, and vomiting. She smoked half pack/daily × 12 years, admitted to social drinking, and occasional marijuana use. Medical, surgical, and family histories were unremarkable. Vital signs on admission: T 102.1 F, BP 93/54, P 96/mt, RR 22/mt. The 8–10 cm erythematous rash on admission was warm and tender to touch. She was alert, oriented, appeared to be in good physical condition with a normal physical examination, except for the abnormal vital signs and the rash. There was no local lymphadenopathy. White blood cell count (WBC) was 5400 (4800–10,800) with 0% bands and Platelets 101,000 (130,00–400,00). Liver function, creatinine kinase, coagulation profile, urinalysis, and chest x-ray were normal. Blood chemistry was normal except for a sodium level of 133 (136–140 mM) and potassium of 3.4 (3.5–5.3 mM). Blood and urine cultures were negative. She received 1 gram of intravenous Ceftriaxone and a 500 CC bolus of normal saline in the emergency room, and doxycycline was added after admission. On day 6, temperature was 99.4, BP 80/40. WBC dropped to 3100 with 11% bands and the platelet count decreased to 85,000. The rash increased to a maximum diameter of 12 cm, while on adequate therapy for LD. On day 7, vital signs were normal; blood chemistry returned to normal, but WBC decreased further to 2900 with 20% bands and the platelet count decreased to 83,000. Ehrlichia chafeensis, Anaplasma phagocytophilum, and Babesia microti IgG and IgM antibodies, and PCR tests and LD western blot test were negative. She was discharged on day 8 and continued oral doxycycline at home to complete 2 weeks. Clinical picture did not fit with any known tick-borne illness. The discharge diagnosis was STARI.
Case 2. 70-year-old female was admitted for “facial cellulitis,” with an area of redness below her right eye. She has not responded to treatment with cephalexin (Keflex) as an outpatient. She was treated with intravenous vancomycin in hospital. The rash persisted and 2 days later, an Infectious Diseases Consult was called. The patient had an area of erythema below right eye, patches of erythema on the nose, right ear, behind right ear, right chest wall, and right posterior trunk. She had no fever. She claimed to have been bitten by ticks, fleas, and mosquitoes. EM and LD were suspected. Rest of the physical examination was normal. Her WBC count and liver functions were normal. Blood and urine cultures were negative. She claimed to be allergic to tetracycline. On treatment with Ceftriaxone, rashes changed in morphology and new rashes appeared. Lyme Western Blot and blood PCR was negative. A. phagocytophilum, E. chafeensis, and B. microti PCR were negative. Ceftriaxone was discontinued. The Patient left the hospital on no treatment. She stated she will self-treat at home with “holistic Medicines.” New rashes appearing while on Ceftriaxone ruled out possible LD.
Discussion
Many dermatologic manifestations have been reported in association with LD. The most common are EM, lymphocytoma, and acrodermatitis chronica atrophicans (Vasudevan and Chatterjee 2013, Middelveen et al. 2015, Tas et al. 2015). EM rashes vary in appearance. They usually have an erythematous base. Macular, maculopapular, vesicular, eschar associated, nodular, pustular, bullous, hemorrhagic, crusting, necrotic, and erosive plaques with various shades of red, pink, whitish, or bluish, varying areas of clearing, or a combination of above may be present. Figure 3 shows numerous EM rashes with some of the variations. The “typical bulls eye lesion” is not common. They present in different sizes and shapes, irregular or smooth borders, numbering from single to many patches (Kluger et al. 2007), and occupying different areas of the body or generalized (Pritt et al. 2016). Recent dermatology literature reported vesicles (Mazori et al. 2015), bullae, hemorrhage, crusting, and necrosis in EM lesions (Paul et al. 2016). There is no other rash comparable to “EM.” Tremendous variation in appearance, and accompanying features, unlike any other organism-associated rash, cannot be explained by a pathogen etiology. Ixodid ticks are prolonged feeders (Lynn 2016) with duration of attachment up to 3 weeks. Amount of saliva injected is variable. Vasodilators injected repeatedly to maintain a liquid blood pool for prolonged feeding could explain the erythema. Bluish and purplish color changes such as a bruise, as well as areas of clearing could be explained by the membrane disrupting and lytic properties of chemicals that lyse red blood cells. Immunomodulatory properties of tick salivary compounds, which inhibit cells involved in the host immune response, could explain the poor antibody response and negative antibody-based tests. The two-tier antibody- based testing currently recommended by the CDC is insensitive for diagnosis of early LD. Culture (Berger et al. 1992) or an ultrasensitive PCR-based assay (Aucott et al. 2017) of skin biopsies gave the highest positivity rates, 86% and 76.9%, respectively. Prolonged attachment is required for transmission of LD. This could be due to time taken for accumulation of sufficient immunomodulatory chemicals required to protect the spirochetes. In experimental studies with I. scapularis, attachment for 72 h was required for 70% transmission, with only 10% transmission after 48 h (Eisen 2018). Ticks retain nutrients from the blood meal and spit back water and sodium into the host with other salivary components. The amount of water expelled was reported to be 74% and sodium 96% for Dermacentor andersoni (Simo et al. 2017). An ixodid tick may increase up to 100 times in size after a full feed (Sauer et al. 2000, Nicholson 2009). The amount of fluid injected back to the host with accompanying salivary chemicals could be enormous.
Ticks transmit viruses, bacteria, rickettsia, spirochetes, and protozoa that are not closely related, but there is an inexplicable and striking similarity in the hematologic and blood chemistry findings in these infections. Leukopenia, thrombocytopenia, abnormal liver function tests, and hyponatremia are common with many tick-borne pathogens (Table 3). Hyponatremia is more frequent in tick-borne encephalitis than with nontick-borne viral meningitis and bacterial meningitis (Czupryna et al. 2016). No attention has been paid so far, to the pharmacologic effects of many chemicals in tick saliva (Table 2) as most of the information is relatively new. Tick salivary proteins have anticoagulant (Narasimhan et al. 2002), antiplatelet, vasodilatory, anti-inflammatory, lytic, and immunomodulatory activities (Kazimirova 2008, Kazimirova and Stibraniova 2013, Simo et al. 2017). Chemicals in tick saliva may play a role in the rashes they produce and clinical and laboratory findings in tick-transmitted diseases. The rash in the two cases presented got worse while on adequate therapy for LD.
Relevant Protein Families in Tick Saliva Found in Other Venomous Animals
Laboratory Abnormalities in Tick-Borne Diseases
Pancytopenia: Lyme disease (Mehrzad and Bravaco 2014), Tick-borne relapsing fever (Castilla-Guerra et al. 2016, Leen et al., 2017).
+, reported blood abnormality.
All hematophagous arthropods have at least one anticoagulant, antiplatelet factor and a vasodilator (Ribeiro 1995, Cordeiro et al. 2015). Many tick salivary protein families and their functions are homologous to proteins found in scorpion, spider, snake, platypus, and bee venoms (Cabezas-Cruz and Valdes 2014) (Table 2). Serine proteinases also found in snake venom affect coagulation, fibrinolytic, and kallikrein-kinin systems and platelets (Serrano and Maroun 2005). Ticks, scorpions, and spiders (Table 2) share a sophisticated salivary cocktail of potent toxic pharmacologic compounds that have dermonecrotic, neurotoxic, and hemorrhagic properties and compounds that inhibit host immune response (Mathias et al. 2011, Stibraniova et al. 2013, Kotal et al. 2015, Simo et al. 2017), neutrophil function (Guo et al. 2009), T and B lymphocytes, platelet aggregation, clotting factors (Zhu et al. 1997a, 1997b), interferon production, and production of IL2 and hemolysins (Simo et al. 2017). Protein families shared by spiders, scorpions, and ticks include neurotoxins, defensins, hyaluronidase, antimicrobial peptides, phospholipases, and proteinases. Alanine-valine-isoleucine-threonine protein and sphingomyelinase have been found in spiders and ticks. Cystatins, lipocalins, and peptidase S1 are only found in ticks (Cordeiro et al. 2015). Phospholipase A2 found in snake venoms display a variety of biological effects, including neurotoxic, myotoxic, cytotoxic, edema inducing, artificial membrane disrupting, anticoagulant, platelet aggregation inhibiting, hypotensive, and other functions (Soares and Giglio 2003). Phospholipase A2 in A. americanum with hemolytic properties is known to hydrolyze membrane phospholipids.
Immunomodulatory components of Ixodid tick's saliva include well-characterized antihistamines, kininases, antioxidants, anticoagulants, prostaglandin E2 that inhibit dendritic cell maturation and Salp15, which inhibits CD4+ T-cell-mediated immune responses in vivo and inhibits killing of spirochetes. Polymorpho-nuclear-leukocytes (PMN) are the first cells to arrive at the site of a B. burgdorferi infection, and tick saliva inhibits critical PMN functions such as phagocytosis and superoxide production. Disintegrins in tick saliva inhibit platelet aggregation. Salp 15 has been shown to enhance spirochete survival and effect joint inflammation in a murine model in vivo. (Kazimirova and Stibraniova 2013, Simo et al. 2017)
The tick salivary microbiome includes many organisms, majority nonpathogenic (Bonnet et al. 2017) and predominantly Proteobacteria (Narasimhan and Fikrig 2015). Their role at present is not fully elucidated, but may facilitate, limit, or block transmitting pathogens (Bonnet et al. 2017) to the host and their establishment. Pathogen transmission and their multiplication is relatively ineffective in the absence of tick saliva. Saliva-assisted transmission has been reported for Thogoto virus (Jones et al. 1989), Tick-borne Encephalitis virus (Labuda et al. 1993), Powassan virus (Hermance and Thangamani 2015), African Swine fever virus (Bernard et al. 2016), Rickettsia conori (Milhano et al. 2015), A. phagocytophilum (Chen et al. 2012), Borrelia afzelii (Pechova et al. 2002), Borrelia lusitaniae (Zeidner et al. 2002), B. burgdorferi (Machackova et al. 2006), and Francisella tularensis (Krocova et al. 2003).
Conclusions
Not all EM rashes are associated with LD. Arthropods that do not transmit LD spirochetes and arthropods other than ticks produce similar rashes. Also, rashes that do not fit the classic EM definition could be easily misdiagnosed as non-Lyme.
The best diagnostic modalities for EM at present are not laboratory tests, with many false negatives. History of a tick bite, clinical manifestations, and presence or absence of a response to LD treatment are more helpful.
Further investigation is needed to determine the role of tick salivary toxins in the etiology of rashes and laboratory findings.
Unusual laboratory findings, history of bites by non-Lyme vectors, and lack of response to Lyme treatment should alert clinicians to the possibility of alternate etiologies of EM rashes.