
Abstract
Culex quinquefasciatus is the principal vector of West Nile virus (WNV) in the South Central United States, yet limited data on host utilization are available. We evaluated host utilization over a 3-month period in 2013 in a residential landscape in College Station, Texas. PCR sequencing of the mitochondrial cytochrome oxidase 1 gene permitted molecular identification of vertebrate bloodmeals to the species level. Forage ratio analysis identified bird species that were overutilized and underutilized by comparing community feeding index values to expected relative abundance values of bird species, derived from eBird data. Community feeding index values were also used in conjunction with reservoir competence data from the literature to generate reservoir capacity index values, a means of identifying relative importance of vertebrate reservoir hosts. Of 498 blood-engorged Cx. quinquefasciatus, 313 (62.9%) were identified to vertebrate species. The majority (95.5%) of bloodmeals originated from avian species with the remainder from mammals, but not humans. Northern mockingbird (Mimus polyglottos) was the principal host for mosquito feeding in June and July, but northern cardinal (Cardinalis cardinalis) became primary host in August. Forage ratio analysis revealed the overutilization of house finch (Haemorhous mexicanus), American robin (Turdus migratorius), northern mockingbird, northern cardinal, white-winged dove (Zenaida asiatica), and mourning dove (Zenaida macroura). Great-tailed grackle (Quiscalus mexicanus), blue jay (Cyanocitta cristata), and Carolina wren (Thryothorus ludovicianus) were under-utilized relative to availability. Reservoir capacity calculations suggested that northern mockingbird and northern cardinal were the principal amplifiers in the study area. These data identify the primary avian species contributing to the enzootic amplification of WNV in East-Central Texas and reveal that the heavy feeding on moderately competent hosts and no feeding on humans likely limit epidemics in this region.
Introduction
C
In Brazos County, TX (county seat: College Station) we are developing a thorough knowledge of vector natural history for arboviruses such as WNV, because recent major outbreaks of WNV have occurred nearby in Dallas to the north and Houston to the southeast (Chung et al. 2013, Randle et al. 2016, Martinez et al. 2017). Currently, the only host utilization data for Cx. quinquefasciatus in Texas are from the more urbanized environments in and around the City of Houston and adjacent regions of Harris County, TX (Molaei et al. 2007). In other parts of Texas, vertebrate amplifiers for WNV have been poorly studied and no bloodmeal host utilization data for Cx. quinquefasciatus are available. Blood-engorged mosquitoes represent contact between vectors and vertebrate reservoirs for arboviruses, so determining which vertebrate species are overutilized by mosquito vectors can help elucidate which bird species are candidate viral amplifiers, and possibly shed light on landscape features that promote vector-amplifier contact.
To generate data pertinent to zoonotic disease control and IPM strategies in Brazos County, Texas, we used DNA sequencing methods to determine the vertebrate hosts that Cx. quinquefasciatus mosquitoes utilize for bloodmeals. These results allowed us to measure community feeding index values for local bird species that we identified from the engorged blood. Furthermore, we used avian relative abundance data generated by citizen scientists through eBird to estimate the expected frequency of vertebrate bloodmeals to test if mosquito-host interaction was purely opportunistic. Differences between observed and expected bloodmeal frequencies (community feeding index and avian relative abundance, respectively), derived from forage ratio analysis, may reveal that certain avian host species are overutilized for bloodmeal acquisition, and indicate species that may be more important WNV amplifiers than others in Brazos County, Texas.
Finally, we combined community feeding index measurements with reservoir competence index estimates from the literature to generate relative estimates of reservoir capacity for candidate amplifier species of birds. Reservoir capacity estimates the relative number of infectious vector mosquitoes derived from naturally infected birds, assuming all birds are susceptible (i.e., not immune due to previous exposure to the virus of interest).
Methods
Mosquitoes were collected in CDC gravid traps (32 locations), CDC light traps (32 locations), and BG Sentinel® traps (16 locations), visited weekly during the period Jun–Aug, 2013, in a residential area of College Station, TX, adjacent to Texas A&M University's main campus (a region encompassing about 9 km2), as previously described (Medeiros et al. 2017). Mosquitoes were sorted by sex, species, and trap (Darsie and Ward 2000). Blood-engorged Cx. quinquefasciatus mosquitoes with Sella score of 2 (fully engorged; Detinova 1962) were separated from collections for bloodmeal identification, following methods previously described (Kent et al. 2009).
Briefly, abdomens were individually macerated in 550 μL PBS with a zinc-coated ball bearing in a Retsch MM300 mixer-mill (Retsch GmbH, Haan, Germany), and clarified by centrifugation. Nucleic acid was extracted by robot (Qiagen Bio Robot Universal System, Qiagen GmbH, Hilden, Germany) and amplified in a PCR reaction using a cocktail of primers for the mitochondrial cytochrome oxidase 1 (CO1) gene (Ivanova et al. 2007). CO1 amplicons were gel-purified and sequenced by Sanger methodology. Sequences were compared with reference CO1 sequences from GenBank and the Barcode of Life Database (BOLD) for bloodmeal identification to the species level.
The observed frequency of bloodmeal host utilization (“community feeding index,” B) was compared to the expected frequency (relative abundance, A) by forage ratio analysis, where B/A > 1 indicates a frequency greater than expected (host overutilization) and B/A < 1 indicates a frequency lower than expected (host underutilization). The forage ratio is neutral if the 95% confidence interval includes unity.
Avian relative abundance data were generated from transformation of eBird checklist frequency data for 268 bird observation checklists from 27 eBird hotspots (Supplementary Table S1; Supplementary Data are available online at
where x is the mean frequency (proportion of checklists that include the species), σ is standard deviation, and n = 12.
For each of the 64 avian taxa represented in the eBird checklists, the eBird mean frequency was transformed to relative abundance (A, “expected frequency”) by dividing each mean frequency by the sum of all mean frequencies. The confidence limits were also divided by this sum, to generate confidence limits for the relative abundances. For each species, the observed frequency of bloodmeals (B, also called “community feeding index”) was the number of species-specific bloodmeals identified divided by all identified bloodmeals. Forage ratio was calculated as observed frequency/expected frequency (B/A) and confidence limits were calculated as B/Amin (upper limit) and B/Amax (lower limit).
To calculate the Reservoir Capacity Index (R), for each species j, we used the following equation:
where B is the community feeding index and C is the reservoir competence index (Kent et al. 2009). Values for C were taken from previously published studies (Komar et al. 2003, 2005, Guerrero-Sánchez et al. 2011, Kilpatrick et al. 2013, Panella et al. 2013).
Results
A total of 498 blood-engorged Cx. quinquefasciatus mosquitoes were collected, with 372 coming from gravid traps (32 trap locations), 121 from BG traps (16 locations), and five from light traps (five locations). Of the 498 mosquitoes processed, 313 of these engorged abdomens produced DNA sequence data that matched CO1 sequences for known vertebrate species in NCBI's nucleotide database. Avian species contributed 299 bloodmeals (95.5% of all bloodmeals identified), all of which were from free-ranging species, except for one chicken (Table 1). Non-avian hosts identified included Virginia opossum (Didelphis virginiana, 8), horse (Equus caballus, 2), and one each of cattle (Bos taurus), black rat (Rattus rattus), eastern cottontail (Sylvilagus floridanus), and striped skunk (Mephitis mephitis). Mammals comprised 4.5% of the identified bloodmeals. Overall, northern mockingbird had the highest community feeding index value (23%) followed by mourning dove (14%) and northern cardinal (12%).
Utilization Code: O, overutilization (confidence interval >1.000); N, neutral (confidence interval includes unity); U, underutilization (confidence interval <1.000); NA, not applicable (confidence interval could not be determined). Only avian species identified among bloodmeals and those with relative abundance above 1% (expected frequency >0.01) are included in this table. The lowest limit of detection by eBird was a relative abundance of 0.02% (expected frequency = 0.0002). Confidence intervals of forage ratios not shown.
Indicates that species was not reported to eBird. For these, a relative abundance value (expected frequency) of 0.0001 was estimated. Utilization code could not be determined for these seven species.
Analysis of bloodmeal host utilization by month revealed seasonal patterns (Fig. 1). In June, Cx. quinquefasciatus fed predominantly on northern mockingbird (28%), great-tailed grackle (12%), mourning dove (12%), and white-winged dove (11%). In July, Cx. quinquefasciatus fed predominantly on northern mockingbird (23%) and mourning dove (17%). In August, Cx. quinquefasciatus fed predominantly on northern cardinal (23%) and northern mockingbird (18%).
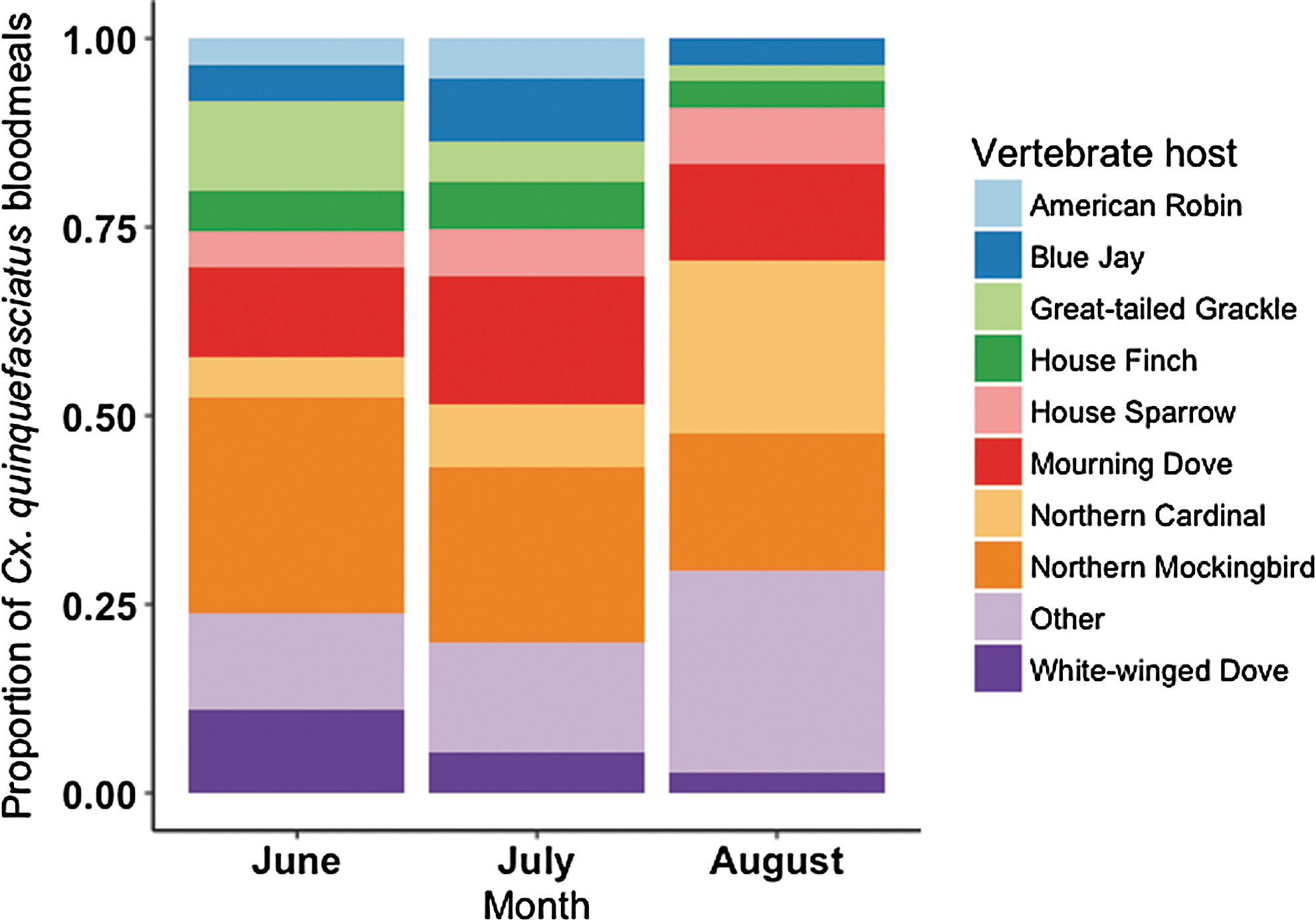
Proportion of host bloodmeals acquired by Culex quinquefasciatus during the months of June (n = 109), July (n = 95), and August (n = 109). Vertebrate hosts aggregated into the category “Other” include barn swallow (n = 1), brown-headed cowbird (n = 4), Carolina chickadee (n = 2), Carolina wren (n = 11), common grackle (n = 2), domestic chicken (n = 1), eastern screech-owl (n = 11), European starling (n = 3), green heron (n = 3), northern bobwhite (n = 1), western kingbird (n = 2), yellow warbler (n = 1), yellow-billed cuckoo (n = 1), black rat (n = 1), cattle (n = 1), eastern cottontail (n = 1), horse (n = 2), Virginia opossum (n = 8), and striped skunk (n = 1).
Forage ratio analysis of bird hosts identified in five or more mosquito bloodmeals revealed, in descending order, the overutilization of eastern screech-owl, house finch, American robin, northern mockingbird, northern cardinal, and mourning and white-winged doves. Fewer than expected bloodmeals were derived from great-tailed grackle, blue jay, Carolina wren, Carolina chickadee, and European starling. Additional bird species that were somewhat abundant (1–5% relative abundance), but were not identified among the bloodmeals presumably were also underutilized. These include killdeer, red-bellied woodpecker, downy woodpecker, and several other species (Table 1). Only two bird species were considered neutral, because their representation among the bloodmeals and their relative abundance in the region were a close match: house sparrow and common grackle (and the calculated forage ratio confidence intervals for these species spanned unity).
Reservoir capacity calculations based on cumulative community feeding index data from across the entire season of the study suggested that northern mockingbird and northern cardinal were the principal amplifiers in the study area (Table 2). Secondary amplifiers were great-tailed grackle, blue jay, house finch, and house sparrow. Monthly analysis of reservoir capacity indicates that northern mockingbird is the primary amplification host in June and July, and northern cardinal emerged as the primary amplification host in August (Fig. 2).
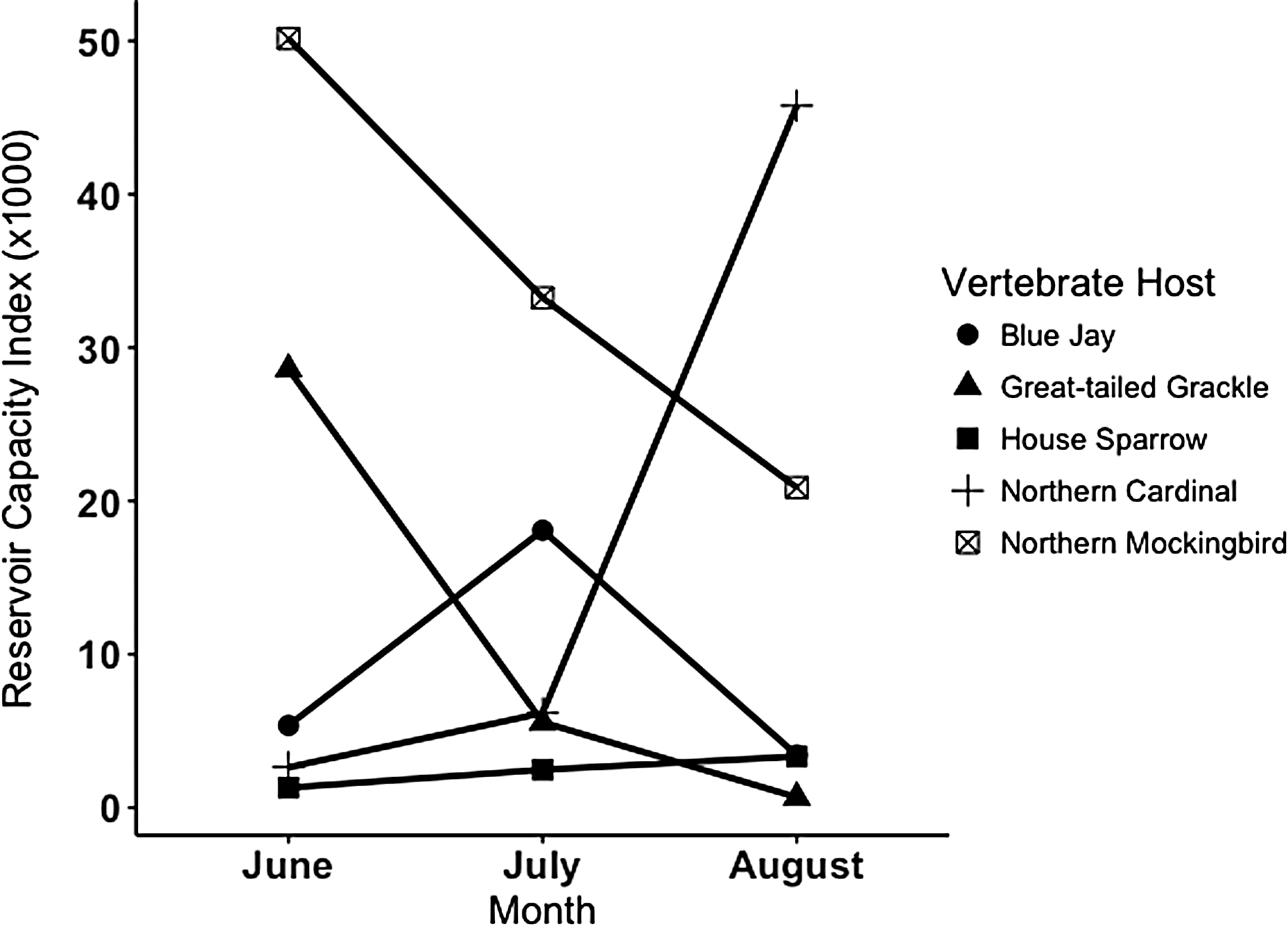
WNV Reservoir Capacity Index values by month for selected avian species in Brazos County, TX. The reservoir capacity index is calculated as Rj = Bj2 × Cj, where B is the monthly community feeding index of Culex quinquefasciatus, and C is the reservoir competence index unique to each avian host (listed in Table 2).
Discussion
We found that the host feeding behavior of Cx. quinquefasciatus in Brazos County was strongly ornithophilic, with over 95% of identified bloodmeals originating from birds, while the remaining bloodmeals originated from a variety of mammals (but none from human). This predominance of avian bloodmeals suggests a feeding profile more similar to Cx. pipiens than Cx. quinquefasciatus, as studies of the latter have detected mammalian bloodmeal proportions from 10% to 70% (Tempelis et al. 1970, Kay et al. 1985, Gomes et al. 2003, Zinser et al. 2004, Molaei et al. 2007, 2010, Levine et al. 2016). Molaei et al. (2007) found that Cx. quinquefasciatus fed on mammals 60.8% and birds 47.4% of 672 identified bloodmeals in and around Houston, TX, during 2005 (their analysis identified 8.3% mixed bloodmeals with both avian and mammal species). Levine et al. (2016) reported on 346 individual feeds by Cx. quinquefasciatus in metropolitan Atlanta, Georgia, during 2010–2011, of which 15.6% derived from mammals and 83.8% from birds.
One explanation for the variation in bloodmeals originating from mammalian hosts is that Cx. quinquefasciatus mosquitoes are opportunistic in their host utilization, as concluded Molaei et al. (2007). Then, if mammals were scarce in our field sites, this would explain the low community feeding index for mammals. However, forage ratio analysis among the avian species selected suggests that most avian species were disproportionately utilized than would be expected based on availability, thus dispelling the opportunism theory.
Although it remains unknown why Cx. quinquefasciatus mosquitoes are highly ornithophilic in Brazos County, low mosquito feeding rates on humans may contribute to the relatively low incidence of human WNV cases in the area compared to the more urbanized regions of Houston (Harris County) and Dallas.
Our results favoring avian over mammalian bloodmeals may also reflect a bias in the study design. Only single-source bloodmeals were identified by the methods used. Mixed source meals would generate mixed sequences, which could not be recognized as matching known sequences. Some mixed bloodmeals may have included mammal blood. In the study of Cx. quinquefasciatus feeding behavior in Tucson, AZ, roughly 5% of the bloodmeals contained mixed avian-mammalian blood (Zinser et al. 2004). As stated earlier, Molaei et al. (2007) detected 8.3% mixed bloodmeals in Houston, while Levine et al. (2016) reported 12.3% mixed bloodmeals. Mixed bloodmeals may be part of the reason why 185 bloodmeals failed to generate a species identification in our study. The other principal explanation is enzymatic degradation of the vertebrate DNA.
Our conclusion that specific host utilization among birds indicates biases for different birds (rather than opportunism) by Cx. quinquefasciatus is predicated on the accuracy of the eBird data for estimating avian relative abundance in the region. While the advent of the eBird project is relatively new compared to that of other citizen science projects like the Christmas Bird Count (CBC) and the Breeding Bird Survey (BBS), we believe that eBird data are less prone to bias than the other data sources.
A recent study found that bird population trends estimated with eBird and BBS were statistically similar (Horns et al. 2018). CBC data reflect winter bird populations, as the data are collected annually between December 15 and January 5. BBS data reflect summer bird populations collected during the breeding season between late May and early July. eBird data allowed us to match the collections of mosquito bloodmeals and presence of birds precisely during the 3-month study period. Thus, an influx of hatch-year birds in late summer due to breeding, or of early fall migrants or postbreeding dispersers in August is reflected by the eBird data, but would not be represented in the other available data repositories for bird populations. However, only 14 checklists of birds were submitted to eBird from the College Station/Bryan metropolitan region between June 1 and August 31, 2013, so to generate more robust estimates of bird populations, we combined eBird data from 10 years of observations.
Utilizing 10 years of data effectively obscures the effects of annual variation, but risks skewing the results of this analysis. We therefore compared the relative abundance estimates and forage ratio estimates for the 2013 dataset and the 10-year dataset, and found that the more robust dataset did not significantly alter the conclusions of the analyses (data not shown).
In Harris County, TX, mourning dove and white-winged dove predominated among the birds most frequently utilized for bloodmeals, followed by several passerine species, including house sparrow, house finch, gray catbird (Dumetella carolinensis), and American robin (Molaei et al. 2007). Our results found principal bloodmeal hosts were passerine species (mockingbird, cardinal), as well as mourning dove. Levine et al. (2016) reported a suppression of WNV transmission due to a shift in feeding from American robin, a moderately competent reservoir, to northern cardinal, a less competent reservoir (Komar et al. 2005). This finding was principally grounded on the concurrent peak of WNV infection in Culex mosquitoes with an observed shift to feeding on cardinals.
In this study, we report the highest community feeding index on low to moderately competent hosts, including northern mockingbird, northern cardinal, and great-tailed grackle. Our calculation of reservoir capacity further suggests that these birds are important in the enzootic cycle of WNV in College Station, TX. Mosquito utilization of northern mockingbird throughout June, July, and August make this abundant species an important amplification host throughout the summer season. Meanwhile, seasonal feeding patterns of Cx. quinquefasciatus suggest great-tailed grackle is more involved in WNV amplification in June and northern cardinal in August. Overall, this analysis suggested that WNV amplification in July was suppressed (in part) by a shift to feeding on less competent avian hosts (Fig. 2). Presumably, the accumulation of immune reservoir hosts late in the summer would also impede transmission.
Blood feeding of mosquitoes on moderately competent bird species might limit the explosive amplification of WNV in the College Station area. During the 2012 WNV epidemic, Dallas County reported 175 West Nile neuroinvasive disease (WNND) cases, which is an incidence of 7.39 per 100,000 (data from Texas Department of State Health Services). Brazos County reported eight WNND cases in 2012, an incidence of 4.12 per 100,000. This suggests that WNV transmission in the enzootic cycle in 2012 was more intense in Dallas County, resulting in more human cases compared to Brazos County. Further studies should identify Culex host feeding patterns in Dallas County to see if the avian community composition and mosquito feeding patterns yield greater amplification potential. Note, however, that these assessments of reservoir capacity do not account for herd immunity, which develops rapidly in the reservoir host populations.
Our analysis of reservoir capacity is the first to find a primary role for northern mockingbird as an amplifier of WNV. Northern cardinal was also important, a finding shared by studies in similar ecosystems in Louisiana and Georgia (Komar et al. 2005, Levine et al. 2016).
Our study identified the primary avian species contributing to the enzootic amplification of WNV in East-Central Texas and revealed that the heavy feeding on moderately competent hosts and no feeding on humans likely limit the epidemics in this region. Our results differ from similar studies in regions where large epidemics occurred, in that important amplifiers are solitary roosters at night when mosquitoes are feeding, rather than communal roosters. Mockingbird and cardinal are solitary roosters. In New York, Chicago, Phoenix, and Los Angeles, communally roosting passerines such as American crow, American robin, house sparrow, house finch, and great-tailed grackle were implicated as important vertebrate amplifiers of WNV (Komar et al. 2001, 2013, Reisen et al. 2006, Hamer et al. 2011). Additional efforts to assess WNV transmission in and around communal roost sites in Brazos County may be warranted.
Footnotes
Acknowledgments
We thank the numerous residents of households in College Station, Texas, that granted us permission to trap mosquitoes on their property. Field and mosquito processing assistance were provided by Emily Boothe, Michael Sanders, Chris Tarrand, Sarah Noe, and Tim Foreman. We appreciate the power of eBird to generate avian relative abundance data, and the hundreds of local citizen science observers who contributed these data. Funding for mosquito collections was provided by the Mosquito Research Foundation. Laboratory testing was funded by the Centers for Disease Control and Prevention.
Author Disclosure Statement
We declare that none of the authors possesses any conflict of interests with respect to the data presented in this article. No competing financial interests exist. The opinions expressed in this article are those of the authors only and do not reflect any official position of either the Texas A & M University or the U.S. Centers for Disease Control and Prevention.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.