
Abstract
Severe fever with thrombocytopenia syndrome (SFTS) is an emerging viral disease in East Asian countries, including China, Japan, and the Republic of Korea (ROK). The causative agent of SFTS is the SFTS virus (SFTSV), which is transmitted by ticks. To investigate the prevalence of SFTSV in the ROK, a total of 9744 ticks were collected from vegetation in five national parks between July and November 2015. Of the collected adult and nymph ticks, Haemaphysalis longicornis (68.44%) was the most abundant, followed by Haemaphysalis flava (29.66%), Ixodes nipponensis (1.56%), and Amblyomma testudinarium (0.34%). Collected larval ticks were of the genera Haemaphysalis (99.61%) and Ixodes (0.39%). One-step RT-PCR and nested PCR were used to detect SFTSV-specific genes from each individual adult and nymph tick and pooled larval ticks. SFTSV was detected in 4.77% (48/1006) in H. longicornis, 1.15% (5/436) in H. flava, 0% (0/23) in I. nipponensis, and 20% (1/5) in A. testudinarium. The infection rate of SFTSV in adult and nymph ticks was 3.61%. The prevalence of SFTSV in adult and nymph ticks was relatively high, compared with previous reports. In larval ticks, the minimum infection rate was 0.31%. SFTSV was detected in ticks collected from both trail and nontrail areas in the national parks, and up to 800 meters above sea level. The sequences obtained showed 99.4–99.7% homology with SFTS virus S segment sequences from Chinese and Japanese ticks.
Introduction
S
The ecological cycle of the SFTSV has not yet been clearly elucidated. However, SFTSV appears to have arthropod vectors and mammalian hosts similar to other Phleboviruses (Liu et al. 2014a). Since SFTSV was discovered, several epidemiological studies have reported that ticks are a key vector of this virus (Yu et al. 2011, Cui et al. 2013, Liu et al. 2014b, 2016). In China and the ROK, SFTSV has been detected in ticks of Haemaphysalis longicornis, Haemaphysalis concinna, Haemaphysalis flava, Ixodes nipponensis, Amblyomma testudinarium, and Rhipicephalus microplus (Zhang et al. 2012, Park et al. 2014, Yun et al. 2014, Luo et al. 2015, Meng et al. 2015, Wang et al. 2015, Liu et al. 2016, Oh et al. 2016, Suh et al. 2016). Among these vectors, H. longicornis is the most dominant in the ROK (Kim et al. 2011, Chong et al. 2013, Park et al. 2014).
Inland national parks in the ROK have relatively well-preserved wildlife, and ticks that parasitize wild animals co-occur in the same habitats. According to a previous study, H. longicornis was the predominant tick collected from wild animals in the ROK (Kim et al. 2014). In addition, SFTSV was detected in the sera and ticks collected from wild animals in the ROK (Oh et al. 2016). Another study reported that SFTSV was detected in the ticks collected from wild reptiles in the ROK (Suh et al. 2016). These findings suggest that the SFTSV might also be detected from ticks collected from the national parks in the ROK. However, to our knowledge, no epidemiological studies of SFTSV have been conducted in the national parks in the ROK. Therefore, we collected ticks from vegetation in five national parks to determine the prevalence of SFTSV in ticks in the ROK national parks.
Materials and Methods
Sample collections
Ticks were collected from five national parks (Bukhansan, Chiaksan, Deogyusan, Juwangsan, and Wolchulsan) in the ROK (Fig. 1) using flags (1 × 1 meters white, cotton, flannel cloth) between July and November 2015. The collection sites were divided into two groups: trail and nontrail (at least 10 meters from a trail) areas. Collected ticks were identified to species level and developmental stages were based on microscopic examinations (Yamaguti et al. 1971). Ticks in adult and nymph stages were identified to species level. Larval ticks were classified to genus level, as identification was restricted by their small size. After identification, all ticks were placed into 2-mL tubes, labeled, and stored at −80°C until assayed. Each tube contained a single adult or nymph tick or pooled larval ticks (at least 10 larval ticks in each tube).
Geographical locations of five national parks in the Republic of Korea where ticks were collected.
RNA preparation
Tick samples were homogenized using the Bead beater TissueLyser II (Qiagen, Hilden, Germany) for 5 min at 30 Hz with a tissue lysis buffer (iNtRON, Seongnam, Korea) and 5-mm sterile stainless steel beads. Then, viral RNAs were extracted from tick lysates using a Viral DNA/RNA Extraction Kit (iNtRON) according to the manufacturer instructions. Extracted genes were stored at −80°C until use.
One-step RT-nested PCR
To detect the SFTSV small (S) segment, each sample was tested using one-step RT-PCR and nested PCR assays. Primary PCR was carried out using one-step RT-PCR premix (Solgent, Daegeon, Korea) with previously designed primers: NP-2F (5′-CAT CAT TGT CTT TGC CCT GA-3′) and NP-2R (5′-AGA AGA CAG AGT TCA CAG CA-3′) (Yoshikawa et al. 2014). The primary reaction was performed with an initial step of 30 min at 50°C and 15 min at 95°C for denaturation, followed by 40 cycles of 20 s at 95°C, 40 s at 52°C, 30 s at 72°C, with a final extension step of 5 min at 72°C. Nested PCR was conducted using 1 μL of the primary PCR product as a template with HiPi PCR premix (Elpisbio, Daejeon, Korea) and previously described primers: N2-F (5′-AAY AAG ATC GTC AAG GCA TCA-3′) and N2-R (5′TAG TCT TGG TGA AGG CAT CTT-3′) (Oh et al. 2016). The reaction for the nested PCR was 25 cycles of 20 s at 94°C, 40 s at 55°C, and 30 s at 72°C. The PCR products were analyzed using gel electrophoresis with 1.5% agarose gel and a 100-bp ladder molecular weight DNA size marker (iNtRON).
Sequencing and phylogenetic analysis
To confirm SFTSV PCR products, positive PCR amplicons were directly sequenced using an Applied Biosystems 3730xl DNA Analyzer (PE Applied Biosystems, Foster City, CA) at Cosmogenetech Co., Ltd. (Seoul, Korea). Sequence homology searches with other deposited sequences were identified using the Basic Local Alignment Search Tool (BLAST) network service. Searched sequences were aligned and analyzed using the program MEGA7 (Kumar et al. 2016). Phylogenetic trees were constructed using the maximum likelihood method. Sequences of the SFTSV S segment that were previously identified in the ROK, China, and Japan were obtained from GenBank for comparative study. The sequences from our study were deposited in GenBank under accession numbers KY497963–KY498000.
Results
A total of 9744 ticks (8274 larvae, 1061 nymphs, and 409 adults) were collected and identified. Among the adult and nymph ticks, H. longicornis was the most frequently collected (68.44%), followed by H. flava (29.66%), I. nipponensis (1.56%), and A. testudinarium (0.34%) (Table 1). All collected larval ticks were identified as one of two genera, Haemaphysalis (99.61%; n = 8242) or Ixodes (0.39%; n = 32) (Table 2).
Infection rate = number of PCR-positive ticks/number of tested ticks × 100.
Minimum field infection rate = number of PCR-positive pools/number of tested ticks × 100.
SFTSV was detected in adult and nymph ticks of H. longicornis, H. flava, and A. testudinarium. Infection rate (the number of SFTSV-positive ticks/the number of tested ticks; IR) in adult and nymph ticks was 4.77% in H. longicornis, 1.15% in H. flava, 0% in I. nipponensis, and 20% in A. testudinarium. The overall IR of SFTSV in adult and nymph ticks was 3.67% (Table 1). SFTSV was detected only in larval ticks of the genus Haemaphysalis. The minimum field infection rate (the number of SFTSV-positive pools/the number of tested ticks × 100; MFIR) in pooled larval ticks was 0.32% (Table 2). The IR of SFTSV in ticks collected from trails was 9.94% in adults and 2.55% in nymphs, whereas the MFIR was 0.11% in larvae. For ticks collected from nontrails, the IR was 3.97% in adults and 2.68% in nymphs, and the MFIR was 0.41% in larvae. Ticks were collected between 69 and 1588 meters above sea level (a. s. l.), and SFTSV was detected up to 800 meters a. s. l.
Of the 80 SFTSV-positive tick samples, 38 were selected from each developmental stage, species, and collection site for sequencing analysis. The sequencing results confirmed that the amplified fragments were very close to the S segment gene sequences of SFTSV. Phylogenetic analysis indicated that the obtained sequences formed three clusters (Fig. 2). The first cluster was closely related to the group of SFTSV strains from humans in Japan with 99.4% homology. The second cluster was closely related to the Korean group with 99.7–100% homology. The third cluster had only one sequence and showed close identity to the SFTSV detected in Chinese ticks with 99.7% homology.
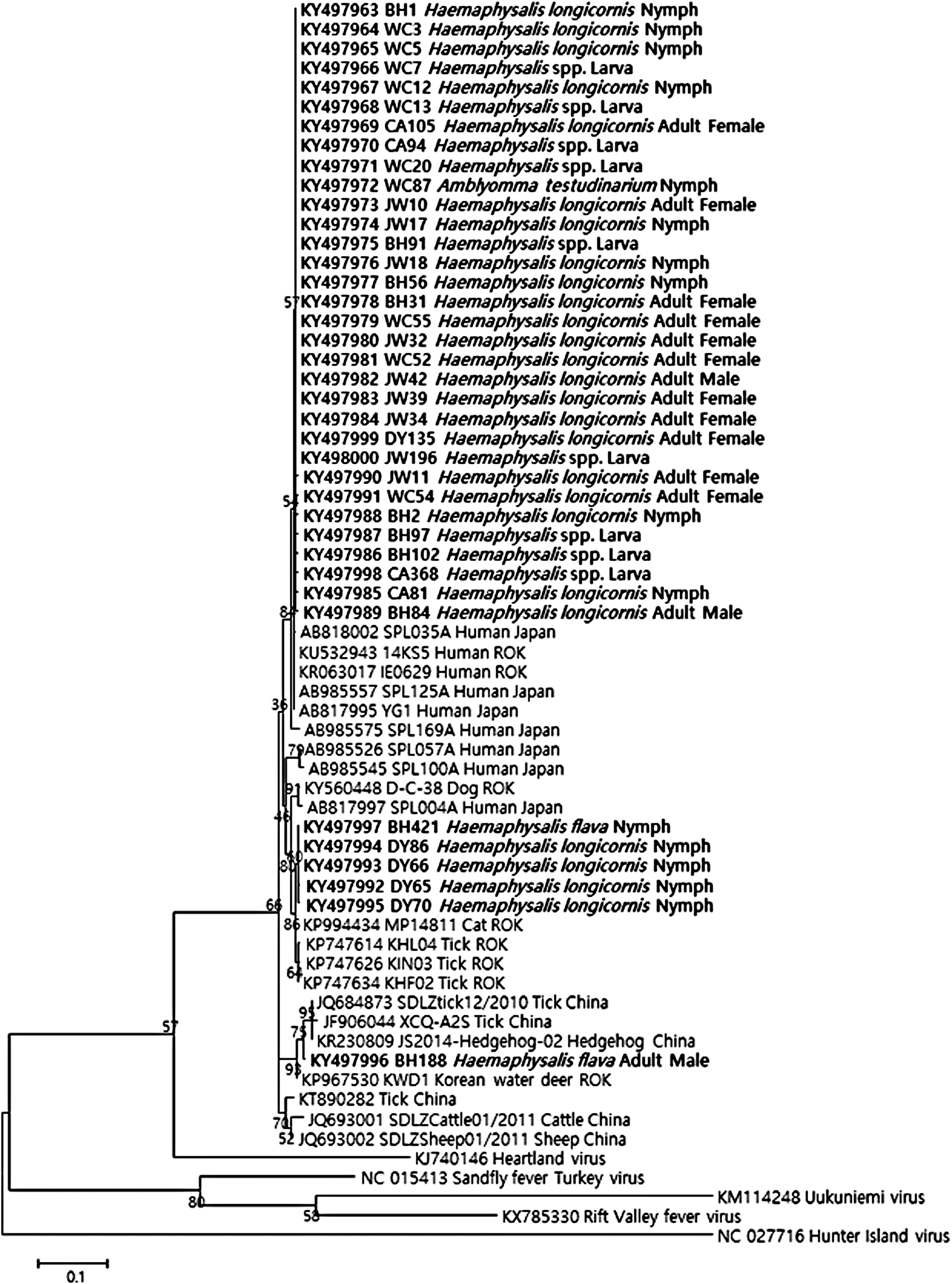
Phylogenetic relationships of SFTSV based on partial S-segment sequences (346 bp). The tree was constructed using the maximum likelihood method based on the p-distance model in MEGA7 (1000 bootstrap replicates). The scale bar indicates nucleotide substitutions per position. Extrabold indicates the original sequences of the SFTSV strains obtained from ticks in this study. BH, Bukhansan; CA, Chiaksan; DY, Deogyusan; JW, Juwangsan; ROK, Republic of Korea; SFTSV, severe fever with thrombocytopenia syndrome viruses; WC, Wolchulsan.
Discussion
There have been many epidemiological studies on the prevalence of the SFTSV in ticks since it was first discovered. In most studies, the prevalence of SFTSV in ticks collected from vegetation has been very low. Two previous studies in China reported the prevalence of SFTSV in ticks collected from vegetation as 0.2% (Luo et al. 2015) and 2.24% (Wang et al. 2015). A study in Japan reported that SFTSV was not detected in 2222 adult and nymph ticks collected from vegetation (Hayasaka et al. 2015). In the ROK, the prevalence of SFTSV in ticks collected from vegetation was 0–0.46% (Ham et al. 2014, Park et al. 2014, Yun et al. 2014). The reason for the low prevalence of SFTSV infection in ticks collected from vegetation is unclear. However, in this study, the SFTSV prevalence rate was estimated as 3.61% in adult and nymph ticks, which was relatively high compared with previous studies. The difference between infection rates in this study and previous studies might have been due to the method of determining prevalence. In our study, the detection of the SFTSV was based on each individual adult and nymph tick and only IR was investigated (due to their small size, larval ticks were pooled to determine MFIR). Conversely, most previous studies attempted to detect SFTSV by pooling a large number of ticks and statistically demonstrating the MFIR. MFIR is a good way to obtain statistical data from a large number of ticks, but it may also underestimate the true IR. Furthermore, several previous studies reported the prevalence of SFTSV was 4.75–6.9% in ticks collected from animals or human bodies, which were tested individually or using fewer pooled numbers of ticks (Yun et al. 2014, Wang et al. 2015, Oh et al. 2016). These findings in this study suggest that the ticks collected from vegetation had a higher prevalence rate of SFTSV than previously reported.
SFTSV was detected in three species of ticks, including H. longicornis, H. flava, and A. testudinarium. Although SFTSV was not detected in I. nipponensis in this study, SFTSV in this species was reported in previous studies (Yun et al. 2014, Oh et al. 2016). Furthermore, SFTSV has also been detected in Haemaphysalis concinna and R. microplus in China (Zhang et al. 2012, Meng et al. 2015). Previous studies indicated that H. longicornis is the predominant tick species in eastern China and the ROK (Chong et al. 2013, Kim et al. 2014, Wang et al. 2015, Coburn et al. 2016), and is the major vector of SFTSV (Yu et al. 2011, Park et al. 2014). Although SFTSV has been detected most frequently in H. longicornis (Liu et al. 2014a, Park et al. 2014), various species of ticks seem to be involved in maintaining this virus.
Our results demonstrated that the viral gene of SFTSV was present in larval ticks (Haemaphysalis) collected from vegetation with an MFIR of 0.32%. These larval ticks did not have any contact with a host and were apparently starved. Recently, SFTSV was detected in all three developmental stages (adults, nymphs, and larvae) of H. longicornis ticks in China, as well as in the eggs (Wang et al. 2015). This suggests that transmission of SFTSV occurs by transovarial routes in ticks. In MFIR addition, Luo et al. (2015) experimentally demonstrated the transstadial and transovarial transmission of SFTSV in H. longicornis ticks in China. However, the prevalence of SFTSV in ticks in nature was not high enough to explain whether vertical infection was completely maintained in the tick population. Although ticks (at least H. longicornis) appear to be the vector and/or reservoir of SFTSV, ticks alone may not be sufficient to maintain this virus in nature. Many of the Phleboviruses in nature are vertically transmitted through their arthropod vectors (Tesh 1988). However, some Phleboviruses do not survive for long periods of time through vertical infection (Fine 1975), and mammalian hosts may be required to maintain the Phleboviruses, including SFTSV.
The occurrence of SFTSV was reported in all provinces of the ROK between 2013 and 2015 (Choi et al. 2016). Furthermore, SFTSV was detected in ticks collected from vegetation of all five national parks surveyed, and also in ticks up to 800 meters a. s. l. In addition, SFTSV was detected in ticks irrespective of whether they were collected from the trails or nontrails of the national parks. Therefore, most areas within the ROK should be considered at risk from SFTSV, including the national parks.
Strains of SFTSV in East Asia are divided into two clades: Chinese and Japanese (Yoshikawa et al. 2015). A previous study demonstrated that many Korean SFTSV strains from humans formed a cluster with the Japanese strains (Yun et al. 2014). Our results show that the subsets of sequences acquired from ticks were classified into three clusters. Most sequences obtained in this study were closely related to the Japanese strains. Our results are similar to previous reports, which indicate that SFTSV isolated from the ROK is closely related to the Japanese strains (Park et al. 2014). However, our results have limited applicability as only the partial sequences of SFTSV were analyzed. Therefore, whole genome sequencing of SFTSV is required for a better understanding of clusters.
This study provides evidence of widespread SFTSV infection in ticks collected from vegetation in the national parks in the ROK. Therefore, forest workers in national parks are at high risk of contact with SFTSV-infected ticks and should take precautions. However, the role of wild/domestic animals as reservoirs and potential additional vectors of SFTSV has not been clarified. Therefore, further study of SFTSV is required for a better understanding of its potential impact on public health.
Conclusions
We determined the prevalence of SFTSV in ticks collected from vegetation in five national parks in the ROK. Our results provide evidence of SFTSV infection in collected ticks and suggest that vertical transmission of SFTSV may occur in tick populations. Moreover, the prevalence of SFTSV in individual ticks collected from vegetation was higher than expected.
Footnotes
Acknowledgments
This study was carried out with the support of “Survey of severe fever with thrombocytopenia syndrome virus in natural environment and study on useful material for prevention of infection I (NIER-SP2015-33),” National Institute of Environmental Research in the Republic of Korea.
Author Disclosure Statement
No competing financial interests exist.