
Abstract
Plague, a flea-borne disease, hampers efforts to restore populations of black-footed ferrets (Mustela nigripes), which occupy colonies of prairie dogs (Cynomys spp.) in North America. Plague is managed by infusing prairie dog burrows with DeltaDust® 0.05% deltamethrin, a pulicide that kills fleas. Experiments are needed to identify pulicides that can be used in rotation with DeltaDust for integrated plague management. In South Dakota, USA, we tested the efficacy of four pulicide dusts when applied at a rate of 8 g per burrow on colonies of black-tailed prairie dogs (Cynomys ludovicianus): Sevin® 5% carbaryl; Dusta-cide® 6% malathion; Alpine® 0.25% dinotefuran with 95% diatomaceous earth; and Tri-Die® 1% pyrethrum with 40% amorphous silica and 10% piperonyl butoxide. We also tested systemic 0.005% fipronil, which was distributed as ½ cup of laced grain per burrow. We sampled prairie dogs on 3294 occasions and detected 10,041 fleas. Sevin and Dusta-cide suppressed fleas but only for 1 month. Neither Alpine nor Tri-Die had any noticeable, consistent effect on fleas. Fipronil suppressed fleas by 97–100% for 3 months. The residual effect of fipronil persisted for ∼12 months. Efficacy of fipronil seems comparable with DeltaDust, which exhibited a residual effect for ∼10 months in prior studies. Continued research is needed to optimize fipronil treatments for plague management on prairie dog colonies.
Introduction
E
Plague is most commonly managed by infusing PD burrows with DeltaDust®, a waterproof powder formulated with 0.05% deltamethrin. The active ingredient opens sodium ion channels and causes hyperexcitation of flea nerve membranes, resulting in flea mortality. DeltaDust can suppress fleas on PD colonies for at least 10 months, and can improve annual survival rates for adult PDs by 31–45% (Biggins et al. 2010) and for adult BFFs by 240% (Matchett et al. 2010). As a result, deltamethrin dust is the primary tool to manage plague on PD colonies. Due to evolution of deltamethrin resistance in fleas, for example, as observed on two PD colonies in South Dakota (Eads et al. 2018), additional tools are needed to manage plague, including pulicides with different active ingredients for attempts to develop an integrated approach to plague management.
In addition to deltamethrin, multiple types of pulicides have been tested with PDs, including 2% carbaryl dust (Barnes et al. 1972), 0.5% permethrin dust (Beard et al. 1992), 0.15% natural pyrethrin 0.3% permethrin dust (Hoogland et al. 2004), systemic 0.025% imidacloprid on edible grain (Jachowski et al. 2011, 2012), and systemic 0.005% fipronil on grain (Poché et al. 2017). To date, fipronil grain has provided the most promising results. In Colorado, fipronil suppressed fleas on black-tailed PDs (BTPDs, Cynomys ludovicianus) (Poché et al. 2017).
Experiments are needed to identify pulicides, in addition to deltamethrin, that can suppress fleas on PDs over the long term. In 2015–2017, we tested the efficacy of five pulicides, including four dusts and systemic fipronil, when used on colonies of BTPDs. Each of the pulicides has a different mode of action on fleas (Table 1). Ultimately, we aimed to identify at least one pulicide in addition to deltamethrin that can suppress fleas on BTPDs for ∼11–12 months.
Pulicides Used in This Study of Fleas Parasitizing Black-Tailed Prairie Dogs on Buffalo Gap National Grassland, South Dakota
Materials and Methods
Study site
Research was conducted under U.S. Geological Survey IACUC protocol 2015-07 (Fort Collins Science Center, Colorado). We studied BTPDs and their fleas on Buffalo Gap National Grassland, South Dakota, during July 10 to October 20, 2015, May 31 to October 26, 2016, and June 5 to October 21, 2017 (43°46′ N, 102°18′ W). Pulicides were tested on three geographically separated BTPD colonies: Cutbank (2015–2017), South Exclosure (2015–2017), and Big Foot (2017). Before our study, burrows in the South Exclosure had been treated with DeltaDust on an annual basis from 2005 through 2014 (Griebel 2014). By 2014, fleas at that colony had developed some resistance to deltamethrin (Eads et al. 2018). Burrows at the South Exclosure were not treated with DeltaDust in 2015–2017. Cutbank and Big Foot had never been treated with a pulicide before 2015 and 2017, respectively.
Study colonies, pulicides, and sampling fleas
We define a site, each of differing shape, as a collection of juxtaposed burrow openings that may be treated with a pulicide or remain nontreated. Dusts were infused at a rate of ∼8 g per burrow using pump-operated dusters (Birchmeier Sprühtechnik AG, Switzerland) and crank dusters (DustinMizer, EarthDuster LLC, Elkins, Arkansas). Fipronil was applied at a rate of ½ cup of grain per burrow opening. For all treatments, pulicides were distributed 50 m beyond the locations where PDs were trapped. Within each colony each year, the edges of the 50 m buffers around the sites were all separated by ∼100 m. Each year, the effective area treated with each particular pulicide was <4 ha due to spatial clustering of BTPD burrow openings.
In 2015, we tested Sevin®, Dusta-cide®, and Alpine®. At Cutbank, all dusts were applied on August 26: Sevin in 851 burrows, Dusta-cide in 531 burrows, and Alpine in 1085 burrows. At the South Exclosure, Sevin was applied to 768 burrow openings on August 27, Dusta-cide was applied to 593 burrows on August 25, and Alpine was applied to 452 burrow openings on August 25.
In 2016, we tested Tri-Die®, which was applied on August 19 at Cutbank (531 burrows) and the South Exclosure (452 burrows) at sites where Dusta-cide and Alpine failed to suppress fleas in 2015, respectively. We also tested fipronil grain, which was applied on July 27 at Cutbank (1085 burrows) and the South Exclosure (768 burrows) at sites where Alpine and Sevin failed to suppress fleas in 2015, respectively.
We consider the evaluations in 2015 and 2016 to be pilot trials. Fipronil proved effective for 3 months at Cutbank and the South Exclosure in 2016. Thus, in 2017, we evaluated the residual effect of fipronil on fleas 11–12 months post-treatment. In addition, to further evaluate the short-term efficacy of fipronil, we retested it on new sites at Cutbank (292 burrows treated July 25), the South Exclosure (136 burrows treated July 24), and Big Foot (198 burrows treated July 24).
Each year, we collected fleas from live-trapped BTPDs during three periods: before pulicide treatments, from 1 to 30 days post-treatment (using the earliest date of any treatment on a colony in a given year as the starting date), and from 31 to 91 days post-treatment. Hereafter, we refer to these periods as Before, After-1, and After-2. We anesthetized each BTPD and combed them as thoroughly as possible for 30 s to collect fleas (Eads et al. 2018). Each BTPD was allowed to fully recover from anesthesia and was released at its capture location. The majority of fleas (>99%) were likely to have been Oropsylla hirusta (Eads et al. 2018).
Statistical analyses
We analyzed data on flea abundance, defined as numbers of fleas on BTPDs, including counts of zero. We conducted separate analyses for the short-term efficacy of pulicides in 2015 and 2016 (pilot trials) and 2017 (extended fipronil research) and the long-term efficacy of fipronil from 2016 to 2017. In each case, we used negative binomial models to compare flea abundance on no-treatment and treatment sites. For interpretation, we present model projections of flea abundance (point estimates) and 95% confidence intervals (“GLIMMIX,” SAS® version 9.3, SAS Institute, Inc.).
We assessed the short-term efficacy of pulicides under a before-after-control-impact (BACI) design. For each year separately, we fit a model with effects for TREATMENT (i.e., no treatment and each pulicide), PERIOD of treatment (i.e., Before, After-1, and After-2), COLONY, and all possible two- and three-way interactions (which helped account for colony differences while assessing flea changes relative to both TREATMENT and PERIOD). If a pulicide was effective over the short term, then flea abundance on a treated site should have declined immediately after treatment and remained low (relative to the nearby no-treatment site) until the end of the field season.
To assess the long-term efficacy from 2016 to 2017 on Cutbank and the South Exclosure, we ran a BACI analysis similar to the assessment described above. We fit a model with effects for TREATMENT, COLONY, and PERIOD (in this case, before treatment = June–July 2016, 11 months post-treatment = June 2017, and 12 months post-treatment = July 2017). We included all possible two- and three-way interactions. If fipronil was effective over the long term, then fleas should have been less abundant on the fipronil sites than on the control sites in June and July 2017.
For the 2015 and 2016 pilot evaluations, multiple treatments on multiple colonies were assessed in single statistical models, increasing the complexity of interpreting interactions. In contrast, fipronil was the sole treatment in 2017, and we assessed the long-term efficacy of fipronil alone. The singular best evidence for an effect in the BACI evaluations of fipronil should be detections of significant TREATMENT × PERIOD interactions that are produced by declines in flea abundance on the treated sites after treatment contrasted with no declines, or minimal declines, on the paired no-treatment sites.
Results
In the 2015 pilot trials, we combed BTPDs on 1276 occasions and detected 5037 fleas. Before treatments at both Cutbank and the South Exclosure, fleas were similarly abundant on the no-treatment, Alpine, and Sevin sites. Fleas were less abundant on the Dusta-cide site than on the no-treatment site at Cutbank, but more abundant on the Dusta-acide site than on the no-treatment site at the South Exclosure (Table 2). In the After-1 period, fleas had increased in abundance on the no-treatment and Alpine sites at both colonies. Flea abundance had declined on the Dusta-cide and Sevin sites at both colonies. In the After-2 period, fleas remained relatively abundant on the no-treatment and Alpine sites at both colonies. Flea abundance had increased on the Sevin sites at both colonies and the Dusta-cide site at the South Exclosure. Flea abundance declined further, but slightly, on the Dusta-cide site at Cutbank (Table 2).
Predicted Abundance of Fleas (95% Confidence Intervals) on Black-Tailed Prairie Dogs at Big Foot, Cutbank, and South Exclosure Colonies, Buffalo Gap National Grassland, South Dakota, 2015–2016
Live-trapped prairie dogs were anesthetized and combed for fleas on 2842 occasions. Data were collected on no-treatment sites, and nearby sites treated with pulicides. Data were collected before pulicide treatments (Before), 1–30 days after treatments (After-1), and 31–91 days after treatments (After-2). Data in this table are available as figures in the Supplementary Data.
CI, confidence interval.
In the 2016 pilot trials, we combed BTPDs on 1566 occasions and detected 3978 fleas. Before treatments at both Cutbank and the South Exclosure, fleas were similarly abundant on the no-treatment and fipronil sites; in comparison, fleas were less abundant on the Tri-Die sites (Table 2). In the After-1 period, flea abundance had increased on the no-treatment sites at both colonies. Flea abundance had increased on the Tri-Die site at Cutbank but declined on the Tri-Die site at South Exclosure. Fleas had nearly disappeared from BTPDs on the fipronil sites at both colonies. In the After-2 period, flea abundance had increased on the no-treatment site at Cutbank and declined slightly on the no-treatment site at the South Exclosure. Flea abundance had declined slightly on the Tri-Die site at Cutbank and increased on the Tri-Die site at South Exclosure. Fleas were still scarce on BTPDs at the fipronil sites (Table 2 and Supplementary Data). Fipronil had suppressed fleas by 97–100% for 3 months. After fipronil grain was distributed at Cutbank, only 8 fleas were collected during 265 capture events at the fipronil site (compared with 793 fleas during 266 capture events at the no-treatment site). After fipronil grain was distributed at the South Exclosure, only 2 fleas were collected during 185 capture events (compared with 1782 fleas during 204 capture events at the no-treatment site).
In the short-term evaluation of fipronil in 2017, we combed BTPDs on 739 occasions and detected 1907 fleas. The treatment effect, assessed by the TREATMENT × PERIOD interaction, was dramatic for Big Foot (p = 0.016) and Cutbank (p < 0.001). The interaction was not supported for the South Exclosure (p = 0.999), where fleas were scarce on the fipronil site before treatment, making it difficult to assess an effect of fipronil (Fig. 1). In the After-1 period, flea abundance had declined slightly on the no-treatment sites at all three colonies. In contrast, fleas had nearly disappeared from BTPDs on the fipronil sites (Fig. 1). In the After-2 period, flea abundance had increased to ∼3–10 fleas per BTPD on the no-treatment sites but remained very low on the fipronil sites (Fig. 1). Fipronil had suppressed fleas by 97–100% for 3 months. After fipronil treatments, 2 fleas were collected during 44 capture events on the fipronil site at Big Foot (compared with 608 fleas during 71 capture events at the no-treatment site), 5 fleas were collected during 169 capture events on the fipronil site at Cutbank (compared with 253 fleas during 111 capture events at the no-treatment site), and no fleas were collected during 15 capture events on the fipronil site at the South Exclosure (compared with 277 fleas during 34 capture events at the no-treatment site).
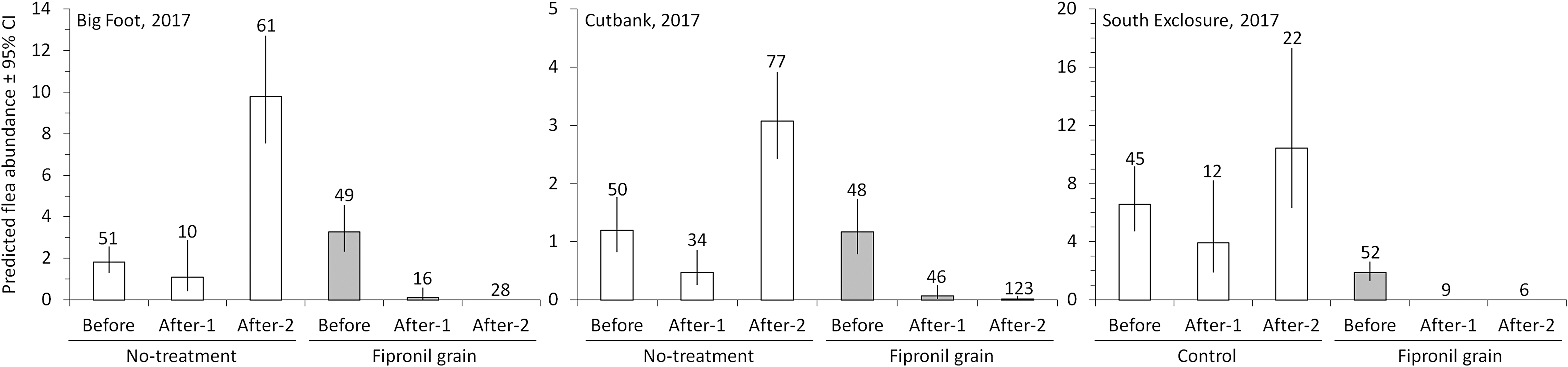
Short-term efficacy of fipronil grain in 2017. Predicted abundance of fleas (95% CIs) on BTPDs at Big Foot, Cutbank, and South Exclosure colonies, Buffalo Gap National Grassland, South Dakota. Data were collected at no-treatment sites and nearby sites treated with fipronil grain during periods before fipronil treatments (Before), 1–30 days after treatments (After-1), and 31–89 days after treatments (After-2). Live-trapped prairie dogs were anesthetized and combed for fleas. Numbers of combing events are presented above the 95% CIs. BTPD, black-tailed prairie dog; CI, confidence interval.
In assessment of long-term fipronil efficacy, we combed BTPDs on 522 occasions (287 of which were from the no-treatment and fipronil sites in June to July 2017, which were included in the short-term evaluation for 2017). We detected 1283 fleas (881 of which were from the control and fipronil sites in June to July 2016). The treatment effect was clearly supported by significant TREATMENT × PERIOD interactions for both Cutbank (p = 0.005) and the South Exclosure (p < 0.001). In June to July 2016, before fipronil treatments, fleas were similarly abundant on the no-treatment and fipronil sites at both colonies. In June and July 2017, 11–12 months after treatments, fleas were significantly less abundant on the fipronil sites than on the no-treatment sites (Fig. 2). At Cutbank in 2017, 17 fleas were collected during 66 combing events at the fipronil site (compared with 60 fleas during 50 combing events on the no-treatment site). At the South Exclosure in 2017, 29 fleas were collected during 74 combing events on the fipronil site (compared with 296 fleas during 45 combing events on the no-treatment site).
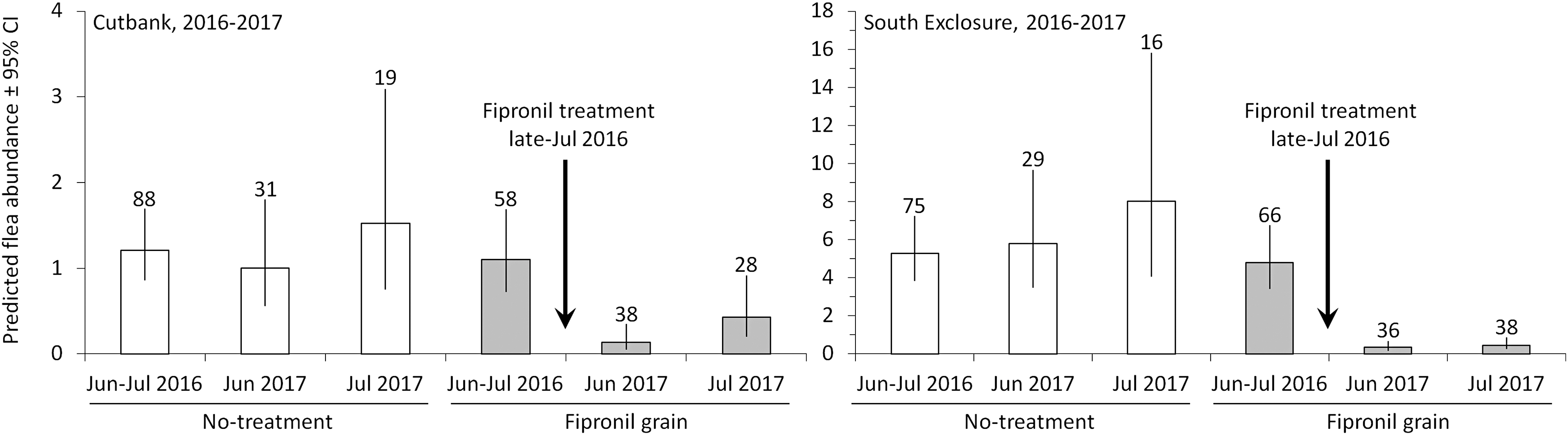
Long-term efficacy of fipronil grain from 2016 to 2017. Predicted abundance of fleas (95% CIs) on BTPDs at Cutbank and South Exclosure colonies, Buffalo Gap National Grassland, South Dakota. Data were collected at no-treatment sites and nearby sites treated with fipronil grain. Data were collected before fipronil treatments (June–July 2016), 11 months post-treatment (June 2017), and 12 months post-treatment (July 2017). Live-trapped prairie dogs were anesthetized and combed for fleas. Numbers of combing events are presented above the 95% CIs.
Discussion
If a pulicide is effective within a BTPD colony, it should cause a rapid decline in flea densities from 1 to 30 days post-treatment, relative to changes in flea densities on the corresponding no-treatment sites. During our study, fleas tended to maintain or increase in abundance after burrows were treated with Alpine or Tri-Die, similar to the patterns observed on paired no-treatment sites. Thus, the two products were ineffective when applied at a rate of 8 g per BTPD burrow opening using pump- and crank-operated dusters.
Both Sevin and Dusta-cide suppressed fleas as evaluated by the BACI design, but only for ∼30 days. Barnes et al. (1972) tested 2% carbaryl Sevin on a colony of BTPDs; the product suppressed fleas for ∼1 month. They infused 85–95 g of the product into each burrow opening, compared with the 8 g of dust we used. Tirador et al. (1967) treated ∼100,000 BTPD burrows with 5% malathion dust; it is unclear how much of the dust was infused into each burrow and efficacy of the dust as a pulicide was not reported.
Carbamates like Sevin are known to degrade quickly under moist conditions (Barnes et al. 1972). During our study, a relatively large front of thunderstorms deposited precipitation at Cutbank and the South Exclosure ∼1 week after Sevin was applied to burrows in 2015. Perhaps, the pulse of moisture reduced the efficacy of Sevin. Moreover, the product's efficacy might be reduced inside BTPD burrows characterized by high humidity (Wilcomb 1954); further, carbaryl exhibits a half-life of only ∼40 days in soil (Johnson and Stansbury 1965, Laveglia and Dahm 1977).
Like other organophosphate dusts, Dusta-cide is somewhat sticky. When applied to burrows using pump- or crank-operated dusters, Dusta-cide fell into burrows as a clumped mass. As BTPDs moved within the burrows, they might have carried some of the Dusta-cide into their burrow systems, thereby causing a brief reduction in flea numbers. However, our inability to coat the inner surfaces of tunnels in BTPD burrows with Dusta-cide might have limited the product's efficacy after 1 month. In addition, organophosphates exhibit a short half-life in soils; their concentrations in soils can decline >90% within a few days (Laveglia and Dahm 1977).
Fipronil was much more effective than the other products as a pulicide during 2016 and 2017. After treatment of sites with fipronil grain in those years, fleas nearly disappeared from BTPDs for 3 months. By June to July 2017, 11–12 months post-treatment, fleas were common on the no-treatment sites but relatively scarce on the fipronil sites. Efficacy of fipronil was comparable with the efficacy of DeltaDust, which suppressed fleas by 98% for 3 months at one of our study colonies in 2015 (on a site at Cutbank; Eads, unpublished data) and suppressed fleas for ∼10 months in a prior study involving three species of Cynomys (Biggins et al. 2010).
Two nonexclusive mechanisms might help explain why fipronil was effective over the long term. First, mammals store fipronil in their fat, and fipronil may be released slowly into the bloodstream, resulting in long-term control of hematophagous fleas. Second, rodents may excrete up to ≥75% of fipronil in their feces (Jackson et al. 2009), and BTPDs deposit feces in their burrows (Wilcomb 1954) where larval fleas develop and feed on organic debris (Krasnov 2008). Larval fleas might consume fipronil-laced feces from BTPDs and die, resulting in long-term flea control (Hinkle et al. 1997, Davis 1999). We are evaluating these hypotheses with field and laboratory studies.
Fipronil grain is appealing as a management tool because it is easily distributed (Poché et al. 2017), most BTPDs readily consume the bait (T. Tretten and J. Hughes, U.S. Fish and Wildlife Service, personal communication), and the product can suppress fleas on BTPDs for ∼12 months. Experiments are needed to evaluate any toxicity of fipronil to BTPDs, BFFs, and nontarget species; fipronil is toxic to some vertebrates (Gibbons et al. 2015). Preliminary results suggest that 0.005% fipronil grain has no noticeable, negative effects on BTPDs and BFFs, but continued study is needed (T. Tretten and J. Hughes, U. S. Fish and Wildlife Service, personal communication).
DeltaDust is an important and established tool for plague management, and our results suggest that fipronil may be added to the toolbox. Research is needed to determine the application rates of DeltaDust and fipronil needed for long-term efficacy and for minimizing the development of resistance in fleas. Current evidence suggests that for prophylactic plague management, either DeltaDust or fipronil should be applied annually. Insecticide resistance in fleas may be reduced if pulicides are rotated over time (Brogdon and McCallister 1998).
In the future, it might be useful to formulate fipronil-treated pellets that can be distributed through all terrain vehicles (ATVs) or aerial drones, methods that have been contemplated since at least the 1970s (Miller et al. 1975) and are available for testing (Krieger et al. 2017). Ultimately, fipronil might be combined with deltamethrin, other pulicides, and supplemental tools for integrated plague management. By protecting PDs against plague, pulicides can help protect a prey base for BFFs, but may not always protect ferrets against plague infection (Livieri et al. 2013). Ferrets can be vaccinated against plague as a proactive measure.
Footnotes
Acknowledgments
Funding and logistical support were provided by the US Geological Survey (USGS); US Forest Service; National Park Service; US Fish and Wildlife Service; Colorado State University (CSU); Prairie Wildlife Research; National Fish and Wildlife Foundation; and World Wildlife Fund. This research was also supported by Grant/Cooperative Agreement Number G14AC00403 from the USGS. We thank many volunteers for their efforts in the field; R. Poché, D. Poché, S. Valentine, L. Polyakova, and Scimetrics Limited Corp. for kindly providing the fipronil grain and for many productive discussions; M.R. Matchett and J. Hughes for discussions on plague management; T. Tretten for assistance; and J. Hoogland and two anonymous reviewers for comments on the article. D. Eads thanks M. Antolin and the CSU Department of Biology for their support. Data from this article are available from Eads (![]() ). Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the US Government.
). Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the US Government.
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Data
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.