
Abstract
Toxoplasma gondii is a zoonotic parasite of major veterinary and public health importance. The parasite is common in many host species in Estonia, however, information about the seroprevalence in the local sheep population is lacking. We estimated T. gondii seroprevalence and evaluated a limited selection of possible risk factors for T. gondii seropositivity in sheep raised in Estonia. Sera from 1599 sheep from 122 flocks, collected during 2012–2013, were diluted to 1:40 and screened for the presence of anti-T. gondii immunoglobulin G antibodies using a commercial direct agglutination test. The cutoff titer for seropositivity was 40. A total of 667 of the 1599 sheep were T. gondii seropositive, yielding an apparent seroprevalence estimate of 41.71% (95% confidence interval 39.31–44.14). At least one seropositive sheep was detected on the majority (88.53%) of the farms. T. gondii seroprevalence was significantly lower in lambs (age <12 months) than in adults (age ≥12 months) (11.11% and 42.16%, respectively). The seroprevalence increased steeply with age in the first years of life. A multivariable random-effects logistic regression model identified being an adult (odds ratio [OR] 9.55) and being from a farm located on the islands (OR 2.51) as risk factors for T. gondii seropositivity, while being of Texel breed appeared as a protective factor (OR 0.34) when compared with the Estonian Blackhead breed. Exposure to T. gondii was widespread among the investigated sheep. Undercooked meat and unpasteurized milk from sheep from Estonia should be considered possible sources of T. gondii infections to other hosts, including humans.
Introduction
T
In addition to public health importance, ovine toxoplasmosis is of major veterinary importance (Blewett and Trees 1987, Buxton et al. 2007, Dubey 2009). In sheep, T. gondii infection can cause clinical signs, such as fever and respiratory distress, and it is one of the main causes of infectious ovine abortions and neonatal mortality worldwide (Dubey and Jones 2008, Dubey 2009, Innes et al. 2009).
In Estonia, northeastern Europe, T. gondii infections are endemic and common in several host species. In our previous studies, more than half (55.8%) of the individuals representing the general human population (Lassen et al. 2016) and more than half (60.8%) of the domestic cats examined were T. gondii seropositive (Must et al. 2015). Furthermore, almost a quarter (24.0%) of examined free-ranging wild boars (Sus scrofa) and moose (Alces alces), which are considered good indicators of environmental contamination with T. gondii, were seropositive (Jokelainen et al. 2015, Remes et al. 2018). In domestic animals, the seroprevalence was estimated to be 5.8% in domestic pigs from breeding herds (Santoro et al. 2017) and 18.6% in cattle (Jokelainen et al. 2017). Ovine T. gondii infections have not been investigated in Estonia before, despite sheep being a relevant host species from several points of view.
We aimed to estimate T. gondii seroprevalence and to evaluate a selection of potential risk factors associated with T. gondii seropositivity in sheep in Estonia. Our primary focus was on animal-level T. gondii seropositivity, whereas flock-level analyses were considered secondary. Our main hypothesis was that exposure to T. gondii would be common in sheep in Estonia.
Materials and Methods
Ethics statement
The sera used in this study were surplus from samples collected for unrelated purposes. No sheep were sampled solely for this study. The sera were stored and analyzed coded. All information was treated confidentially.
Setting, samples, and study design
The target population was mainly adult (>1-year-old) sheep raised in Estonia. At the beginning of 2012, there were a total of 80,189 sheep (of these 45,486 were breeding animals, i.e., adult animals) in 1897 sheep flocks (ARIB 2012).
The samples we used in this study had been primarily collected for unrelated infectious disease surveillance and monitoring purposes and mainly from breeding animals. In brief, 10% of all registered sheep flocks in Estonia were randomly selected for testing for the original purposes. From each selected flock, adequate number of sheep aged 6 months or older were selected to be able to detect at least 20% prevalence of brucellosis in this age group, with 95% confidence and assuming perfect diagnostic test (VFB 2012). The blood samples were collected by authorized veterinarians on the farms, from the jugular vein of live sheep. The samples included in this study were collected between March 14, 2012, and June 5, 2013, and the sera were stored frozen until analysis.
Owing to the study design, this study did not include a risk factor questionnaire. Information on the age and the breed of each sheep, the size of each flock, and the location of each farm were obtained from the Estonian Agricultural Registers and Information Board. Using the samples and data that were available, we conducted a cross-sectional seroepidemiological study to investigate naturally acquired T. gondii infections in sheep in Estonia.
We set as inclusion criteria that the samples were from individual sheep that were raised in Estonia, and that there was a sufficient quantity of serum available for the analysis. From the original sample of 1662 serum samples, we excluded samples of 59 sheep due to insufficient amount of serum for analysis, and the second samples from 4 sheep that had been sampled twice. The sample size in this study was 1599 sheep from 122 sheep flocks (1–65 sheep per flock), which was evaluated to be sufficient for estimating the seroprevalence at animal level but limited for a flock-level estimate. We did not aim to detect or state freedom of T. gondii infection or disease caused by the parasite within each flock. The sample size evaluation was performed with Epitools software (Sergeant 2017) using confidence level of 95%, acceptable error of 5%, 25% as the expected animal-level seroprevalence or 76% as the expected proportion of flocks positive (Jokelainen et al. 2010), and 45,486 breeding animals or 1897 flocks as the population size (ARIB 2012). Ignoring clustering, the minimum required sample size for an apparent animal-level seroprevalence estimate would have been 288 sheep. Owing to expected clustering, we included all 1599 samples that were available, to keep the number of flocks included as high as possible. The minimum required sample size for an apparent flock-level prevalence estimate would have been 245 flocks.
Serology
The sera were screened for the presence of specific anti-T. gondii immunoglobulin G antibodies using a commercial direct agglutination test (Toxo-Screen DA, bioMérieux, Marcy-l'Étoile, France), following the instructions of the manufacturer. The sera were diluted to 1:40. The results were read after 18 h of incubation using a 4-point scale and interpreted conservatively as negative or positive (Jokelainen 2013). The cutoff titer for seropositivity was 40 (Jokelainen et al. 2010) and only clear positive reactions (the agglutination mat covering at least half of the bottom of the well) were considered positive results (Jokelainen 2013). An antigen control and the negative and positive controls provided in the kits, diluted 1:40 and 1:4000, were included on each plate. The antigen control included all reagents except sample serum to control that the antigen solution did not create a false positive reaction by itself.
Statistical analyses
Data recording and coding were performed in Microsoft Excel 2013 (Microsoft Corporation by Impressa Systems, Santa Rosa, California). We used OpenEpi software to calculate 95% confidence intervals (CIs) (Mid-p exact) for the proportions and two-tailed p values (Mid-p exact, or when that was not calculated, uncorrected chi-square) for the Two-by-Two-Table comparisons (Dean et al. 2017). Further statistical analyses and building of models were done with Stata 13.1 software (Stata Corporation, TX). We report p values for all the analyses.
The outcomes were dichotomous: a sheep was defined seropositive if its serum sample yielded a positive result, otherwise the sheep was considered seronegative, and a flock was considered positive if at least one of the tested sheep from the flock tested seropositive, otherwise the flock was considered negative. In addition to simple apparent seroprevalence estimate, an apparent animal-level seroprevalence estimate and 95% CI that were adjusted for clustering (sampling weights) were calculated using the Epitools two-stage prevalence survey analysis calculator (Sergeant 2017). For flocks that were represented by at least five sheep, we also report the proportion of flocks that had at least three seropositive sheep, as well as proportion of sheep that were seropositive within the flocks.
The evaluated potential risk factors for animal-level T. gondii seropositivity were age and breed of the sheep, the size of the flock, and the location of the farm. The evaluated potential risk factors for finding at least one seropositive sheep in a flock were the size of the flock and the location of the farm. Strength of association between the potential risk factors and the outcomes was assessed using odds ratio (OR).
The age of each sheep was rounded to full months. Age was evaluated as a dichotomous variable (lamb: <12 months vs. adult: ≥12 months) (Halos et al. 2010, Tegegne et al. 2016, Deksne et al. 2017), as a continuous variable, and graphically as a categorized variable. The breeds were evaluated as dummy variables; the breeds presented by <10 sheep were categorized as ‘other breed’. The breed represented by the highest number of sheep was used as the reference breed. Flock size was evaluated as a dichotomous variable (small flock: <100 sheep vs. large flock: ≥100 sheep) (Deksne et al. 2017). The location of the farm was evaluated by using counties as dummy variables and Harjumaa County, where the capital city Tallinn is located, as the reference county (Remes et al. 2018) and alternatively as a dichotomous variable: mainland vs. islands (the two large islands, Hiiumaa County and Saaremaa County).
First, univariable logistic regression models for animal-level T. gondii seropositivity were used to evaluate the potential risk factors separately. Next, multivariable logistic regression models were built using backward elimination: all variables were included in the model, and then those that were nonsignificant and did not act as confounders were removed, starting from the highest p value, until only variables with p values <0.05 and any confounders were left. If elimination of a variable from the model altered the measure of association of another variable by >20%, this was considered an indication of confounding. The variable “farm” was included as a random factor in all models. The age of the sheep and the location of the farm were offered to the multivariable model as the dichotomous variables.
Secondly, logistic regression models for finding at least one seropositive sheep in a flock were used to evaluate the potential risk factors separately and together.
Results
Descriptive data
The age of the sheep included in this study ranged from 3 to 159 months (0.27–13.22 years), the median age was 51 months (4.24 years), and the mean age was 55 months (4.54 years). The majority of the sheep were adults (age ≥12 months) (Table 1). Six breeds were represented by at least 10 sheep: the Estonian Blackhead (33% of sheep in the sample), crossbreds (32%), the Estonian Whitehead (16%), the Oxford Down (13%), the Suffolk (3%), and the Texel (2%); other breeds (1%) were the Icelandic, the Dala, the Dorset, and the Ouessant (Table 1).
Animal-Level Toxoplasma gondii Seroprevalence in Sheep in Estonia, and Proportion of Sheep Flocks with At Least One Toxoplasma gondii Seropositive Sheep
Age was unknown for 52 sheep and breed for 44 sheep.
The Icelandic, the Dala, the Dorset, the Ouessant.
No sheep from Läänemaa County, Põlvamaa County, and Pärnumaa County.
Flock size ranged from 2 to 3530 sheep (median 134; mean 234), and 1 to 65 sheep (median 14; mean 17) were tested per flock. The farms were located in different parts of the country: 52 farms (760 sheep) in the mainland and 70 farms (839 sheep) on the islands; 12 out of the 15 counties were represented (Table 1).
Seroprevalence
Of the 1599 sheep, 667 tested T. gondii seropositive, yielding an apparent seroprevalence estimate of 41.71% (95% CI 39.31–44.14) (Table 1). When adjusting for clustering (sampling weights), the animal-level seroprevalence estimate among 1598 sheep from 121 flocks (one sheep omitted because it was the only sheep sampled from the flock) was 30.38% (95% CI 20.66–39.90).
At least one seropositive sheep was detected in 108 of the 122 flocks (88.53%; 95% CI 81.92–93.32) (Table 1). A total of 87 (75.00%; 96% CI 66.53–82.24) of the 116 flocks from where at least 5 sheep were sampled had at least 3 seropositive sheep. The median proportion of seropositive sheep within these 116 flocks was 35.71%, the mean was 44.79%, and the proportion ranged from 0.00% to 100.00% (i.e., with this sampling, no seropositive sheep was found in some of these flocks, whereas all tested sheep were seropositive in some flocks).
Risk factors
The seroprevalence was higher (p < 0.001) in adult sheep than in lambs (Table 1). The youngest seropositive sheep was 11 months (0.9 years) old, and the oldest seropositive sheep was the oldest in the sample, >13 years old. The seroprevalence by age is shown in Fig. 1. Based on the univariable random-effects logistic regression analysis, adult sheep had 13.52 (95% CI 3.23–56.62) times higher odds to be T. gondii seropositive than lambs. A univariable model using age as a continuous variable suggested that odds to test seropositive increased with 22% (95% CI 15–30) for each additional year of age. In the final multivariable random-effects logistic regression model (Table 2), adult age was a significant risk factor for animal-level T. gondii seropositivity, and age acted as a confounder.
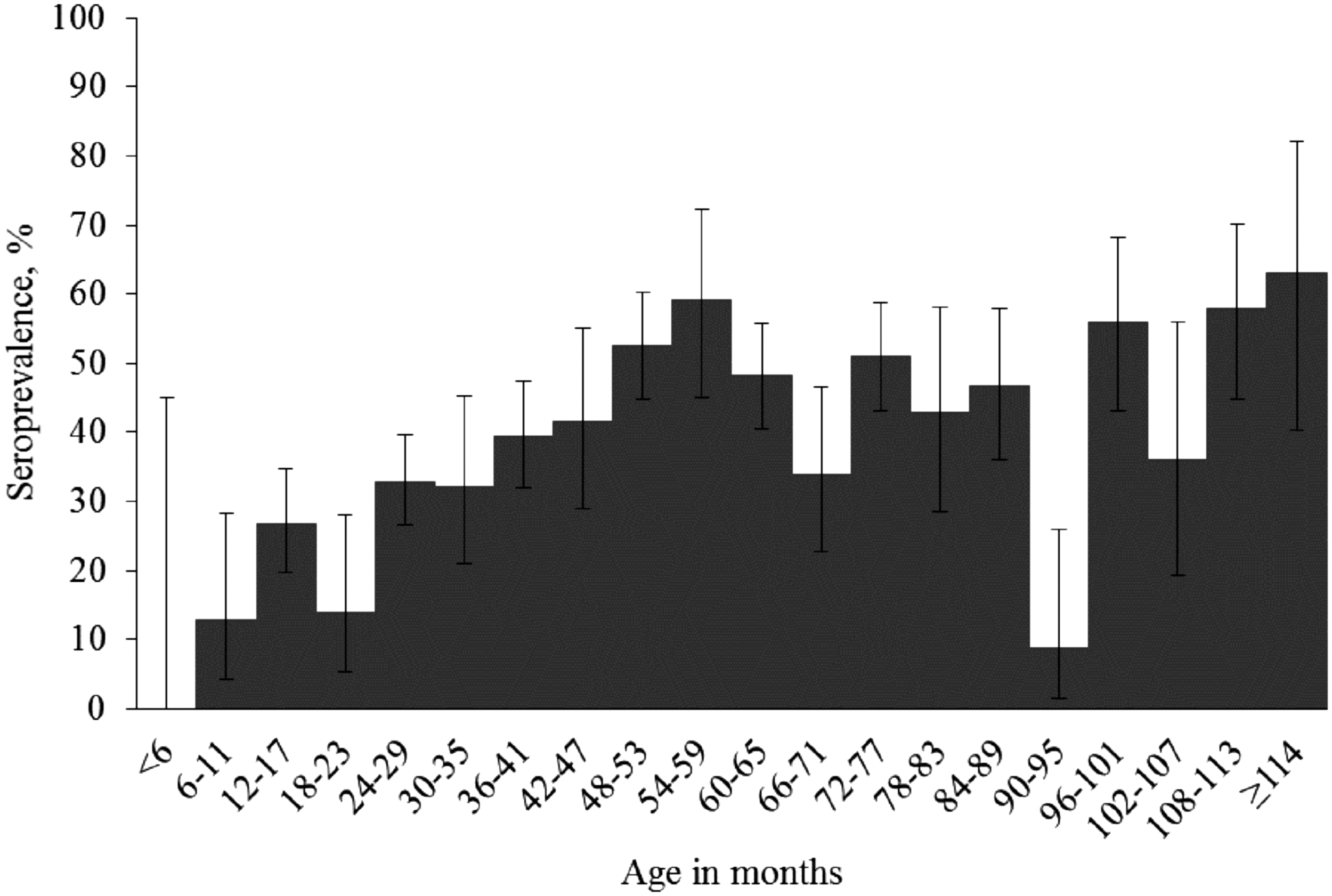
Toxoplasma gondii seroprevalence in sheep in Estonia, by age in months, with 95% confidence intervals.
Results of Multivariable Random-Effects Logistic Regression Model for Animal-Level Toxoplasma gondii Seropositivity, Estimated for 1555 Sheep in Estonia
The Icelandic, the Dala, the Dorset, the Ouessant.
The seroprevalence of T. gondii by sheep breed is given in Table 1. The seroprevalence was the highest (47.20%) in the Estonian Whitehead sheep and significantly lower (37.38%, p < 0.01) in sheep of the other local breed, the Estonian Blackhead sheep. In the final multivariable model (Table 2), which used the Estonian Blackhead breed as the reference breed, being of the Texel breed appeard as a protective factor (OR 0.34).
The seroprevalence was higher (p < 0.01) in sheep from small flocks than in sheep from large flocks (Table 1), but flock size did not appear as a significant factor for animal-level T. gondii seropositivity nor for flock-level T. gondii positivity in the univariable analyses (p = 0.360 and p = 0.988, respectively) and it was removed from the multivariable models as statistically nonsignificant.
The animal-level seroprevalence and the proportion of flocks with at least one seropositive sheep are given by counties in Table 1. Based on the univariable analysis, being from any of the counties was not a significant factor for animal-level T. gondii seropositivity nor for flock-level T. gondii positivity, when comparing with the reference county, Harjumaa County.
The seroprevalence was higher (p < 0.001) in sheep raised on the islands than in sheep raised on the mainland (Table 1). Being from a farm located on the islands was a significant risk factor for animal-level seropositivity in the univariable analysis (OR 2.50; 95% CI 1.29–4.84) and in the final multivariable model (Table 2). Location on the islands was not a significant factor for flock-level T. gondii positivity based on the univariable analysis (p = 0.981).
Discussion
The results of this study demonstrated that exposure to T. gondii was widespread in sheep that were raised in Estonia during 2012–2013: the seroprevalence estimate is high. Notably, the seroprevalence estimate (41.71%; 95% CI 39.31–44.14) was 1.7 times higher (p < 0.001, chi-square) than an estimate reported from Finland (24.6%; 95% CI 22.7–26.5), which was obtained with the same method and cutoff for seropositivity and from a similar sample of sheep (Jokelainen et al. 2010). Similarly, the seroprevalence in domestic cats, a key host species for the epidemiology of T. gondii, and the seroprevalence in free-ranging moose (A. alces), an indicator species of environmental oocyst contamination, were higher in Estonia than in Finland (Jokelainen et al. 2010, 2012, Must et al. 2015, Remes et al. 2018). These data together show that the epidemiological situation differs in the two countries.
For sheep, lower T. gondii seroprevalence estimates than our estimate from Estonia have been reported also from other Nordic and Baltic countries, but mostly using enzyme-linked immunosorbent assays (ELISA), making the results not directly comparable. Ignoring methodological differences, in contrast to our result, all other seroprevalence estimates from this region were <25%: 24.6% (95% CI 22.7–26.5) in Finland (p < 0.001, chi-square) (Jokelainen et al. 2010), 18.8% (95% CI 16.0–21.76) in Sweden (p < 0.001) (Lunden et al. 1992), 17.2% (95% CI 15.0–19.6) in Latvia (p < 0.001) (Deksne et al. 2017), and 16.2% (95% CI 14.65–17.93) in Norway (p < 0.001, chi-square) (Skjerve et al. 1998). Higher T. gondii seroprevalence estimates than our estimate from Estonia were reported for sheep from central and southern European countries. The range of seroprevalence estimates is wide: 87.4% (95% CI 82.0–93.4) in Belgium (p < 0.001) (Verhelst et al. 2014), 84.5% (95% CI 81.4–87.7) in Serbia (p < 0.001) (Klun et al. 2006), 74.0% (95% CI 72.54–75.43) in Great Britain (p < 0.001, chi-square) (Hutchinson et al. 2011), 53.7% (95% CI 49.13–58.25) in Greece (p < 0.001) (Diakoua et al. 2013), and 49.3% (95% CI 44.9–53.7) in Spain (p = 0.004) (García-Bocanegra et al. 2013). The Serbian study used the same method as we used in our study, but a lower cutoff titer for seropositivity (titer of 25), and the studies from Belgium, Greece, and Spain used ELISA. Taken together, these results suggest that exposure to T. gondii among sheep was more common in Estonia than in other Baltic and Nordic countries, but less common in Estonia than in the central and southern European countries. The reasons for these differences, in particular for the higher seroprevalence in Estonia than in other countries in the region, remain unknown but could include, for example, different management of sheep, differences in feline populations, and different local environmental factors; a multicountry epidemiological study using a detailed questionnaire could be used to investigate these aspects.
Almost 90% (95% CI 81.92–93.32) of the flocks included in our study had at least one seropositive sheep. Proportions of sheep flocks defined positive that did not significantly differ from our estimate from Estonia have been reported from northern Italy (87.5%; 95% CI 74.3–100.0; p = 0.853) (Gazzonis et al. 2015), southern Spain (84.7%; 95% CI 75.0–91.7; p = 0.453) (García-Bocanegra et al. 2013), and Sweden (77.8%; 95% CI 65.3–87.4; p = 0.076) (Lunden et al. 1992). Higher proportions have been reported from Greece (100.0%; 95% CI 94.2–100.0; p = 0.007) (Diakoua et al. 2013) and Belgium (96.2%; p = 0.010; 95% CI 92.9–98.2) (Verhelst et al. 2014), whereas lower proportions have been reported from Campania region in Italy (77.8%; 95% CI 69.6–84.6; p = 0.028) (Fusco et al. 2007), Finland (76.3%; 95% CI 67.1–84.0; p = 0.018) (Jokelainen et al. 2010), Norway (44.3%; 95% CI 37.5–51.4; p < 0.001) (Skjerve et al. 1998), and Latvia (44.0%; 95% CI 33.7–54.8; p < 0.001) (Deksne et al. 2017). Overall, the flock-level estimates indicate that the parasite is present in a substantial proportion of sheep flocks, and this was now also demonstrated in Estonia. This study was not primarily designed for estimating flock-level or within-flock prevalences, thus the flocks with no seropositive sheep detected should not be considered free of T. gondii.
The sample size was high, and the sample was suitable for estimating the animal-level seroprevalence, but suboptimal for flock-level estimate and for evaluating flock-level risk factors. From some farms, only a single or few sheep were sampled, but in contrast, the range of flock size was wide: the sample included also very small flocks. Overall, the sample, which was 3.5% of the target population of adult sheep and where 6.4% of all registered flocks were represented (ARIB 2012), could be considered representative of sheep raised in Estonia. Regression model including random factor (“farm”) is generally considered to be sufficient to control clustering effect due to nonhomogeneous sampling and to avoid biased estimates of associations. However, sampling bias could affect the results, and, for example, the results by breed should be interpreted with caution.
The serology method we used for analyzing the sera has been widely used for sheep (Dubey et al. 2008, Halos et al. 2010, Jokelainen et al. 2010, Lopes et al. 2013a). Serology is an indirect method; it detects the specific humoral immunological reaction of the host against the parasite, not the presence of viable parasites. The cutoff for seropositivity we used in this study can be considered high, and thus the seroprevalence estimate is conservative. Moreover, using only a single dilution, some false negatives are possible due to undetected prozone phenomenon, which is caused by excess of antibodies for the test. We chose to use one method and a single dilution because the expected benefit of using another method or several dilutions was considered low compared with the costs. Another method in series could have been used to confirm the seropositivity status of the sheep, and using two serological methods in parallel would have increased sensitivity to detect seropositive animals. We did not expect cross-reactions to affect the results, but it should be emphasized that the prevalence of parasites that could potentially cause cross-reactions in serology (e.g., Neospora caninum) is unknown in sheep in Estonia.
One of the major strengths of our study was that we had detailed information on the age of the investigated sheep (Fig. 1). T. gondii seroprevalence increased steeply with age in the first years of the life of the sheep (Fig. 1), indicating that there was substantial infection pressure. As expected, being an adult sheep was a risk factor for T. gondii seropositivity, also in the final model wherein the breed of the sheep and the location of the farm were included (Table 2). Older age has been identified as a risk factor for T. gondii seropositivity also in other studies (Dumétre et al. 2006, Halos et al. 2010, Lopes et al. 2013a, Tegegne et al. 2016, Deksne et al. 2017). These results indicate that most ovine T. gondii infections are acquired postnatally. Our results (Fig. 1) further exemplify why it is important that seroepidemiological T. gondii studies report the age range of the animals sampled and include age of the individuals in the analyses. The major sampling-related issue in this study was the low number of samples from lambs. In contrast, focus was on adult sheep, the age range was wide, and the results are comparable with those from Finland: the sheep in the Finnish study were all >1 year of age (Jokelainen et al. 2010).
Differences in T. gondii seroprevalence by sheep breed have been reported (Deksne et al. 2017), and a possibility that the pathogenesis could vary by sheep breed has been previously discussed (Williams et al. 2005). We have observed and investigated differences in T. gondii seroprevalence by breed in other host species (Jokelainen et al. 2017, Must et al. 2017). In this study, T. gondii seroprevalence was estimated for the sheep breeds represented in the sample. Some of the breeds appeared as a risk factor or a protective factor for testing seropositive, also when the age of sheep and the location of the farm were included in the model and “farm” was used as a random factor. This appeared even more pronounced when the reference breed was changed to Texel breed (Supplementary Data S1, Supplementary Tables S1, S2, S3, S4, S5, S6, S7, and Supplementary Data S2, Supplementary Tables S8, S9, S10, S11, S12, S13, S14) but the number of sheep of Texel breed included was limited. It needs to be emphasized that this study did not aim to investigate differences in seroprevalence by breed. Further research on the topic should include higher number of samples per breed as well as information on relevant epidemiological factors, such as farm management, collected using a pathogen-specific questionnaire.
In this study, sheep from small flocks had higher apparent animal-level T. gondii seroprevalence than sheep from large flocks (Table 1), but in the further analyses, flock size was not a significant factor for animal-level T. gondii seropositivity nor for finding at least one seropositive sheep in the flock. In the neighboring country Latvia, sheep from large flocks had higher odds to test seropositive than sheep from small flocks (Deksne et al. 2017), whereas in Italy, adjusted for the number of sheep tested, the number of seropositive sheep in a flock decreased with increasing flock size (Cenci-Goga et al. 2013). The latter result was discussed to be due to more intensive management of larger flocks. A study from Greece, in contrast, showed that sheep kept under conditions defined as intensive or semi-intensive had higher odds of being T. gondii seropositive than sheep kept under conditions defined as extensive (Tzanidakis et al. 2012).
Farm location by county was not a significant factor for animal-level T. gondii seropositivity nor for flock-level T. gondii positivity. This indicates that T. gondii is present and widespread in the agricultural environment across the country. This result is in line with the results from cattle and wild boars (Jokelainen et al. 2015, 2017).
Sheep raised on the two large Estonian islands had higher odds to be T. gondii seropositive than sheep from the mainland. Similarly, in Finland, the highest seroprevalence was reported from the Åland islands (Jokelainen et al. 2010). These observations might be linked to different management practices, differences in feline populations, or to different local environmental or climatic conditions on the islands than on the mainland. Estonia is a small country that has temperate climate; the islands and the coastal areas in the west are warmer during winters than the mainland. That is, the environmental and climatic conditions on the islands might be more suitable for survival of T. gondii oocysts. Unfortunately, details about the management of the farms and feline populations were unavailable, and we were, therefore, unable to investigate these aspects in this study. Future studies should include information on relevant epidemiological factors, such as farm management, collected using a pathogen-specific questionnaire.
Our study design allowed a large sample size and was economical and ethically sound but, being an observational study based on a convenience sample from unrelated monitoring purpose and data that were available from the official registers, we were unable to evaluate some potential risk factors. The lack of specific risk factor questionnaire limited the epidemiological conclusions we could make. We used “farm” as a random factor in the models, that is, accounted statistically for factors that we could not directly measure. It would have been important to evaluate, for example, details of the management of the farms as well as presence of cats on the farms and, in particular, their access to feed, water, or pastures, as potential risk factors, but this was unfortunately not possible in this study. What is important to emphasize is that the sample included a good number of separate farms of different sizes. Different management practices were thus included, and the overall estimate can be considered representative of the overall management of sheep in the country. Another aspect that should be emphasized is that the samples for this study were collected during 2012–2013, and the epidemiological situation may have changed since then.
As herbivores, sheep likely acquire T. gondii by the fecal–oral route: by ingesting sporulated oocysts, which have been shed in unsporulated form in feces of infected felids. Seropositive sheep can thus be seen as indicators of contamination of their environment, feed, or water with T. gondii oocysts. However, other routes of infection are also possible and include transplacental, venereal, and lactogenic transmission (Lopes et al. 2013b, da Silva et al. 2015, Luptakova et al. 2015, Klauck et al. 2016).
After T. gondii infection, both specific antibodies and tissue cysts are presumed to persist for life in sheep (Dubey 2009, Opsteegh et al. 2016). T. gondii tissue cysts have been found in several edible parts of infected sheep, and consumption of undercooked mutton is considered a risk factor for human T. gondii infection (Spósito Filha et al. 1992, Cook et al. 2000, Belbacha et al. 2004, Dubey 2009, Opsteegh et al. 2016). Moreover, eating raw or undercooked meat of animals infected with T. gondii poses a risk also for other host species, including domestic cats that can themselves become clinically ill from the infection and contribute to the environmental reservoir of the parasite (Jokelainen et al. 2012, Must et al. 2015, 2017). The results of this study are relevant from the food safety, pet feed safety, veterinary, public health, and One Health point of views.
Conclusions
Exposure of sheep to T. gondii was demonstrated to have been widespread in Estonia. The infections were likely acquired as the seroprevalence increased with age, and the infection pressure appeared high. Seropositivity was associated with farm location on the islands and being of some breeds, which may reflect farm management and inherent host characteristics, respectively. Assuming that the seropositivity indicates presence of infectious T. gondii parasites in the tissues of the sheep, undercooked meat and unpasteurized milk from sheep from Estonia should be considered possible sources of T. gondii infections to other hosts, including humans.
Footnotes
Acknowledgments
We thank the Estonian Veterinary and Food Laboratory and Estonian Sheep Breeding Society for their collaboration, and the Estonian Agricultural Registers and Information Board for providing data. We thank Pille Paats for laboratory excellence and Kädi Neare for data management and contributing to the preparation of the data set. The study was supported by the health promotion research program TerVe 3.2.1002.11-0002 EKZE_SS from the Estonian Research Council, by project funding M14143VLVP from the Strategic Development Fund of the Estonian University of Life Sciences and 8P160014VLVP from the Base Funding of Estonian University of Life Sciences, and by the Niemi Foundation.
Author Disclosure Statement
No conflicting financial interests exist.
Supplementary Material
Supplementary Data S1
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Table S6
Supplementary Table S7
Supplementary Data S2
Supplementary Table S8
Supplementary Table S9
Supplementary Table S10
Supplementary Table S11
Supplementary Table S12
Supplementary Table S13
Supplementary Table S14
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.