
Abstract
This study examined sand fly diversity and the natural infection of sand flies by Leishmania in three localities in Rondônia State: Santo Antônio and São Vicente farm, which are anthropized environments, and the Jamari FLONA, which has not been anthropized. Sand flies were collected in canopies and at ground level using CDC light traps and BG-Sentinel CO2 traps. Female sand flies were grouped into pools that were subjected to DNA extraction and the samples were subjected to PCR to detect and identify Leishmania. A total of 6696 specimens were examined and 52 species were identified. Abundance was greater in canopies than at ground level (n = 5657 and n = 1039, respectively); however, fauna composition was more diverse at ground level. A total of 4870 specimens were collected using CDC light traps and 1826 specimens were collected using BG-Sentinel traps. The presence of Leishmania DNA was detected in 12 pools, and Leishmania species profiles were identified in three of these pools: one pool of Psychodopygus amazonensis and one pool of Psychodopygus chagasi infected with Le. (Vi.) braziliensis, and one pool of Psychodopygus davisi infected with Le. (Le.) amazonensis. Sand flies are sensitive to environmental modification and species diversity is greater in regions that have experienced less anthropization. Although the annual infection rate of cutaneous leishmaniasis is high in Rondônia State, the role that sand flies play in the leishmaniasis transmission cycle remains poorly understood. The high abundance of Ps. davisi and Trichophoromyia auraensis and the presence in these species of natural infection by Leishmania spp. suggest that these species are acting as vectors of tegumentary leishmaniasis in Rondônia.
Introduction
Sand flies (Diptera: Phlebotominae) are insects of medical importance that transmit several pathogens (Ready 2013). They are the primary vector for protozoans of the Leishmania genus (Maroli et al. 2012, Ready 2013). At present, ∼1000 sand fly species have been identified throughout world, 530 of these species occur only in the Americas (Shimabukuro et al. 2017), and 98 species are proven or putative vectors of leishmaniasis (Maroli et al. 2012). In Brazil, 12 species are proven vectors of leishmaniasis at the moment (Rangel and Lainson 2009).
Leishmaniasis is a disease with several clinic-pathological manifestations that impact public health (Akhoundi et al. 2017). Clinical aspects of the disease range from the benign cutaneous form (with the possibility of spontaneous recovery) to more severe forms (Silveira et al. 2004, Akhoundi et al. 2017). This range of manifestations is primarily a function of parasite virulence and host immune response (Shaw et al. 2007). There are two clinical forms of the disease: visceral leishmaniasis (VL) and tegumentary leishmaniasis (TL), which affects the skin or mucosa (Silveira et al. 2004, Akhoundi et al. 2017). The primary etiological agent of VL in Brazil is Leishmania (Leishmania) infantum, whereas TL is caused by a variety of species from the Leishmania genus (Gontijo and Carvalho 2003). In Brazil, TL is transmitted by a wide array of pathogens (Leishmania species), reservoirs (armadillos, sloths, monkeys, and rodents), and vectors (sand fly species) (Roque et al. 2014, Akhoundi et al. 2017). Several different transmission patterns are, therefore, possible and this makes it difficult to develop effective measures for combating the disease (Maroli et al. 2012, Ready 2013). From 1995 to 2014, there were, on average, 25,763 new cases registered annually with an average detection coefficient of 14.7 cases per 100,000 inhabitants (Brasil 2018). The north region had the highest detection coefficient of 55.53 cases per 100,000 inhabitants (Brasil 2018). Pará State had the highest number of cases in the region, followed by Amazonas and then Rondônia (Brasil 2018). In Rondônia, ∼1000 new cases of TL have been recorded every year since 2007, with an incidence rate of 71.54 cases per 100,000 inhabitants (Cantanhêde et al. 2015, 2018). In Porto Velho, the state capital of Rondônia, 1601 cases of TL were recorded between 2007 and 2017—the highest number of cases in the state with a coefficient of 33.96 cases per 100,000 inhabitants (Brasil 2018).
At least five species of Leishmania occur in Rondônia State: Le. (Le.) amazonensis, Le. (Viannia) braziliensis, Le. (Vi.) guyanensis, Le. (Vi.) lainsoni, Le. (Vi.) lindenbergi, and Le. (Vi.) shawi (Cantanhêde et al. 2015, 2018); of these, Le. (Vi.) braziliensis is the most prevalent among recorded cases. Sand fly species diversity impacts the diversity of Leishmania species that cause TL and the number of human cases; 131 sand fly species are known to occur in Rondônia (Resadore et al. 2017). Among the vectors of TL that are widely distributed in Brazil, the following have been recorded in Rondônia: Bichromomyia flaviscutellata, Bichromomyia olmeca nociva, Nyssomyia antunesi, Nyssomyia umbratilis, Nyssomyia whitmani, Psychodopygus davisi, Psychodopygus wellcomei, and Trichophoromyia ubiquitalis (Galardo et al. 2015, Ogawa et al. 2016, Resadore et al. 2017). Ps. davisi is a putative vector after the record of this species with Le. (Vi.) naiffi (Gil et al. 2003). Despite the presence of these vectors, few studies have been conducted to determine the role that these species play in the transmission cycle of this region.
Sand fly studies are important because they improve our understanding of leishmaniasis transmission dynamics. This study examined sand fly species composition in Rondônia State, with a focus on species that have been implicated in leishmaniasis transmission. Sand flies were collected at different stratification levels (canopy and ground) in three locations with differing phytophysiognomies. Sand fly fauna were assessed by comparing diversity, richness, abundance, and natural infection rate by Leishmania.
Materials and Methods
Study areas
This study was conducted in two rural areas and a FLONA (National Forest) in Rondônia State, western Amazon, Brazil. Sand flies were collected in the localities of Santo Antônio (08°53′7.10′′S, 64°0′55.90′′W) and São Vicente farm (09°34′55.50′′S, 64°51′57.50′′W), both in the municipality of Porto Velho, and in the Jamari FLONA (09°15′33.78′′S, 62°54′13.73′′O), in the municipality of Itapuã d'Oeste (Fig. 1). The localities of Santo Antônio and São Vicente farm are characterized by ombrophilous forest. These localities are home to a range of phytophysiognomies that have developed in response to anthropic activities related to the Jirau and Santo Antonio Hydroelectric Power Plants, respectively. The Jamari FLONA is an area dedicated to sustainable forest management. It is a predominantly open ombrophilous forest, but ∼10% of the tract is occupied by dense ombrophilous forest (IBGE 2012). Four collection points were selected in each locality and each point was divided into two strata.

Sand fly collection sites in the Porto Velho and Itapuã d'Oeste municipalities, Rondônia State, Brazil. South America
Sand fly collection and taxonomic identification
The collections were performed after the authorization obtained in Chico Mendes Institute, Ministry of Environment (SISBIO/ICMBio) with the license number 42353-2. Sand flies were collected during three expeditions conducted between August 2015 and February 2016. Collections were made from 5:00 PM to 8:00 AM, for 3 consecutive days, using four CDC light traps and four BG-Sentinel CO2 traps installed in the tree canopy (20 m) and at ground level (1.5 m). The array of traps was altered in the stratum each day in the localities: first day, canopy—four CDCs, ground—four BGs; second day, canopy—two CDCs and two BGs, ground—two CDCs and two BGs; and third day, canopy—four BGs and ground—four CDCs.
The males (whole insect) and females (head and last three segments of the body) were clarified in 10% potassium hydroxide and slide mounted with Berlese fluid. Species identification was based on the morphological characters of Galati (Galati 2016). The thorax and part of the abdomen of females were stored in 96% alcohol and maintained at −20°C preceding molecular analysis. This study adopted the generic abbreviations of Marcondes (2007).
DNA extraction and PCR for detection and identification of Leishmania
The females were divided into pools of 2 to 20 individuals, and pools were sorted according to collection area, vertical stratification (canopy or ground), and expedition. DNA extraction was performed using the DNeasy Blood and Tissue kit (Qiagen, Hilden, Germany); manufacturer's instructions were followed with the exception of a minor change: 2.5 μL of Proteinase K was used in the cell lysis process.
PCR was directed to a region of Leishmania kDNA using the primers 5′-GGG(G/T)AGGGGCGTTCT(G/C)CGAA-3′ and 5′-(G/C)(G/C)(G/C)(A/T)CTAT(A/T)TTACACCAACCCC-3′ (120 pb) (Oliveira et al. 2005). The reaction mixture comprised 1X Buffer, 1.5 mM of MgCl2, 0.8 μM of each primer, 0.2 mM of dNTPs, 1.25 U of Taq DNA Polymerase (Invitrogen, CA), and 3 μL of DNA in a final volume of 25 μL. The samples were amplified in an automatic thermocycler (Applied Biosystems®), with initial denaturation at 94°C for 5 min, followed by 30 cycles at 94°C for 30 s, 55°C for 30 s, and 72°C for 45 s, and a final extension at 72°C for 10 min. The amplified products were analyzed in 2% agarose gel stained with GelRed (Biotium, Fremont).
The positive samples from the first reaction were subjected to PCR directed to the hsp70 region using the primers Hsp70cF 5-GGACGAGATCGAGCGCATGGT-3′ and Hsp70cR 5′-TCCTTCGACGCCTCCTGGTTG-3′ (240 pb) (Da Graça et al. 2012). PCR was performed using 1X Master Mix 2X Go Taq (Promega, Madison), 1 μM of primer, and 5 μL of DNA in a final volume of 50 μL. The amplification of hsp70 was performed in an automatic thermocycler (Applied Biosystems) with initial denaturation at 94°C for 5 min, followed by 30 cycles at 94°C for 30 s, 61°C for 1 min, and 72°C for 3 min, and a final extension at 72°C for 8 min. The positive samples were subjected to PCR-restriction fragment length polymorphism (PCR-RFLP) using the HaeIII (ThermoScientific) and BstUI (ThermoScientific) enzymes. The reaction mixture comprised 1X Buffer R, 5 U of enzyme, and 5 μL of amplified material in a final volume of 15.5 μL, incubated at 37°C for 16 h. PCR-RFLP products were analyzed in 10% polyacrylamide gel stained with Buffer Dream (ThermoScientific).
Sequencing reaction
To confirm the presence of Leishmania, PCR products from pools containing hsp70 were subjected to a purification reaction using the GFX PCR kit DNA and Gel Band Purification (GE Healthcare) and sequenced with the Sanger RPT01-E platform (at the Research Center René Rachou—Fiocruz Minas) using a 3730 DNA Analyzer (Applied Biosystems). The sequences were analyzed using the software Phred, Phrap, and Consed, with the minimum value defined as Q = 30; the consensus sequence was generated and the nucleotides with a Q value <30 were considered as N. Identities, and similarities were analyzed using BLASTn, available at (
Data analysis
Interpolation curves (real data) and extrapolation curves (tendency of the data) were constructed to assess sampling effort and compare species diversity between vertical stratifications and between localities. These curves were based on Hill numbers expressed in q values on the axis of the abscissa (Chao et al. 2014). Permutational multivariate analysis of variance (PERMANOVA) (consisting of a multivariate nonparametric analysis based on permutations) was applied to assess the influence of localities, strata, and collection methods on sand fly species composition. Statistical analyses were performed on RStudio software using the programming language R, version 3.4.3. Minimum natural infection rates were obtained by multiplying the number of positive pools by 100 and dividing the result by the total number females in these pools.
Results
A total of 6696 sand flies were captured [2064 (30.85%) males and 4632 (69.15%) females] and 52 species from 13 genera were identified (Table 1). The genera with the highest richness and abundance were Psychodopygus (11 species, n = 3476 individuals, 51.91%), Nyssomyia (7 species, n = 2218, 33.12%), and Sciopemyia (3 species; n = 226, 3.37%). Lower richness and abundance were exhibited by the following genera: Lutzomyia (two species, n = 163 individuals, 2.43%), Trichophoromyia (two species, n = 150, 2.24%), Evandromyia (nine species, n = 118, 1.76%), Trichopygomyia (two species, n = 113, 1.69%), Psathyromyia (six species; n = 88, 1.31%), Pintomyia (one species, n = 72, 1.07%), Bichromomyia (two species; n = 37, 0.55%), Viannamyia (two species, n = 22, 0.33%), Micropygomyia (four species, n = 10, 0.15%), and Migonemyia (one species, n = 3, 0.04%). The most abundant species were Psychodopygus chagasi sensu lato (n = 2083 individuals, 31.10% of the total collected) followed by Nyssomyia yuilli yuilli (n = 740, 11.05%), Ps. davisi (n = 671, 10.02%), and Ny. antunesi (n = 627, 9.36%).
Sand Fly Species Collected with CDC and bg-Sentinel Traps in Jamari FLONA, Santo Antônio, and São Vicente Farm Localities, Rondônia State, Brazil, Between August 2015 and February 2016
Suspect species or incriminated vector of Leishmania.
The Jamari FLONA exhibited the highest richness (47 species) and the second highest number of individuals [n = 2.236 specimens (33.41% of the total collected) with 1648 females (73.64%) and 588 males (26.30%)]. São Vicente farm had the second highest richness (36 species) and the lowest number of individuals [n = 1231 (18.38%), with 805 females (65.39%) and 426 males (34.61%)]. Santo Antônio had the lowest richness (30 species), but the highest number of individuals [n = 3229 (48.21%), with 2180 females (67.51%) and 1049 males (32.49%)] (Table 1). Mean sample coverage for São Vicente Farm, Santo Antônio, and Jamari FLONA was estimated as 99.00%, 96.00%, and 94.50%, respectively, which indicates that sampling is nearly complete for all sites (Fig. 2A). The greatest value of the exponential Shanon index (q = 1) was observed in Jamari FLONA 16.30 (confidence interval [CI]: 16.07–17.11) followed by São Vicente Farm 11.50 (CI: 11.22–12.50) and Santo Antônio 6.45 (CI: 6.40–6.78). This implies that the species richness, when q = 1, is significantly greater in Jamari FLONA. Futhermore, when the inverse of the Simpson diversity index (q = 2) was computed, a similar result was found (Table 2). The PERMANOVA test indicated that significant differences exist between the localities studied (p < 0.05).

Sample coverage of sand fly richness observed between August 2015 and February 2016.
Diversity Estimates for Rarefied and Extrapolated Samples for São Vicente Farm, Santo Antônio, and Jamari FLONA Localities, Rondônia State, Brazil
CI, confidence interval.
Greater richness and abundance were found in canopies [48 species/n = 5657 individuals (84.49%)/3930 females (69.45%) and 1727 (30.53%) males] than at ground level [37 species/n = 1039 individuals (15.51%)/702 females (67.56%) and 337 males (32.44%)]. When CDC traps were installed in canopy, mean sample coverage estimated was 95.00%, when the traps were installed near the ground level, the value was 94.00%, which indicates that in booth strata, the sampling coverage was near complete. According to values of the inverse Simpson diversity index, the highest species richness was found at ground level 21.79 (CI: 21.32–22.96) than at canopy 10.8 (CI: 10.79–11.24). The same pattern was found when the Shannon diversity index was used. The difference in sand fly fauna composition between strata was statistically significant (p < 0.05).
CDC traps proved to be the most efficient collection method. Richness and abundance were greater among sand flies collected with CDC traps [50 species/n = 4870 individuals (72.72%)/3341 females (68.59%) and 1529 males (31.41%)] than collections made with BG-Sentinel traps [33 species/n = 1826 individuals (27.28%)/1291 females (70.66%) and 535 males (29.34%)]. The difference between sand fly fauna captured by each method was statistically significant (p < 0.05).
A total of 4328 female sand flies were grouped into 276 pools for the purpose of molecular detection, and 12 pools tested positive for mitochondrial DNA of Leishmania (Fig. 3). Eleven pools from the canopy tested positive; the infected species included Ps. chagasi s.l., Ps. Davisi, Ny. yuilli yuilli, Ny. umbratilis, and Psychodopygus amazonensis. Only one ground level pool tested positive and the infected species was Trichophoromyia auraensis. The general minimum infection rate was 0.28% (12 pools/4328 females). The species with the highest infection rate was Th. auraensis (8.33%), followed by Ps. amazonensis (4.35%), Ps. davisi (0.76%), Ny. yuilli yuilli (0.31%), Ny. umbratilis (0.29%), and Ps. chagasi s.l. (0.27%). The product generated by the digestion of enzymes HaeIII and BstUI (Fig. 4) was used to identify Leishmania species in three pools: Le. (Vi.) braziliensis in one pool of Ps. amazonensis and one pool of Ps. chagasi s.l.; and Le. (Le.) amazonensis in one pool of Ps. davisi. The Leishmania species were identified by sequencing hsp70 from two infected pools (Ps. davisi and Ps. amazonensis), and species identities were confirmed by comparing them with specimens deposited in GenBank: Le. (Le.) amazonensis (94% similarity) and Le. (Vi.) braziliensis (98% similarity) (deposited under GenBank codes MG029123.1 and HF586369.1, respectively).
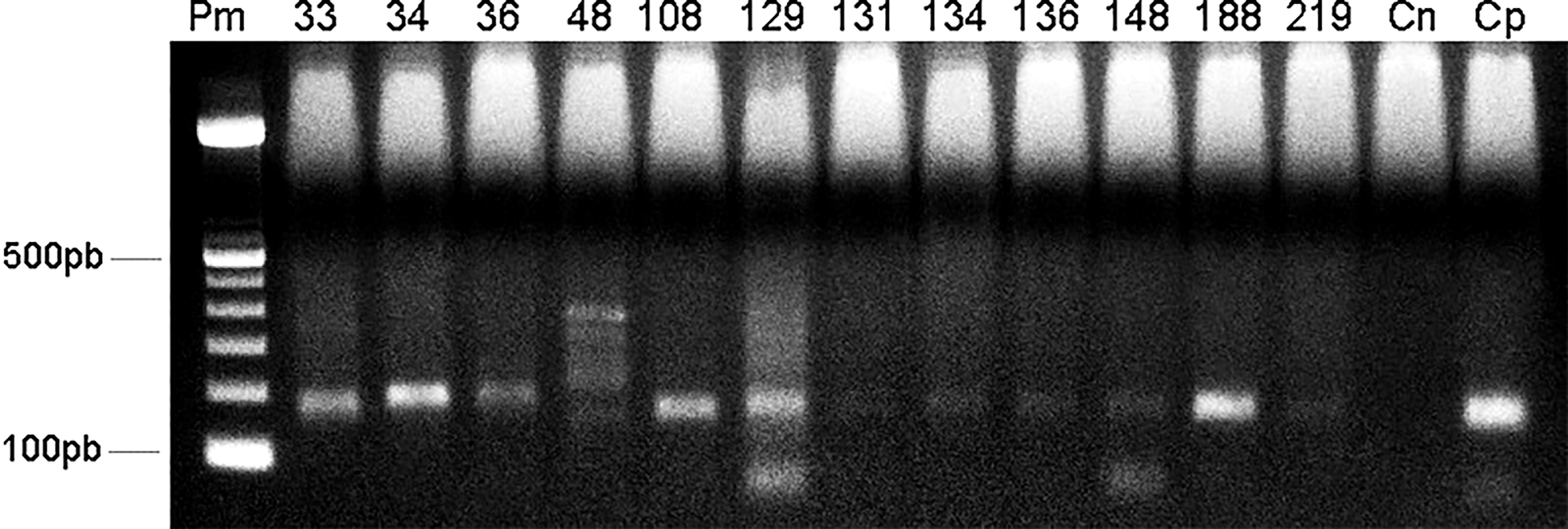
Positive pools for kDNA PCR in 2% agarosis gel stained with GelRed. Pm: molecular pattern; 33: Psychodopygus davisi; 34: Ps. davisi; 36: Nyssomyia umbratilis; 48: Trichophoromyia auraensis; 108: Nyssomyia yuilli yuilli; 129: Psychodopygus amazonensis; 131: Psychodopygus carrerai carrerai; 134: Psychodopygus chagasi s.l.; 136: Ps. chagasi s.l.; 148: Ps. davisi; 188: Ny. yuilli yuilli; 219: Ps. chagasi s.l.; Cn: negative control; Cp: positive control.
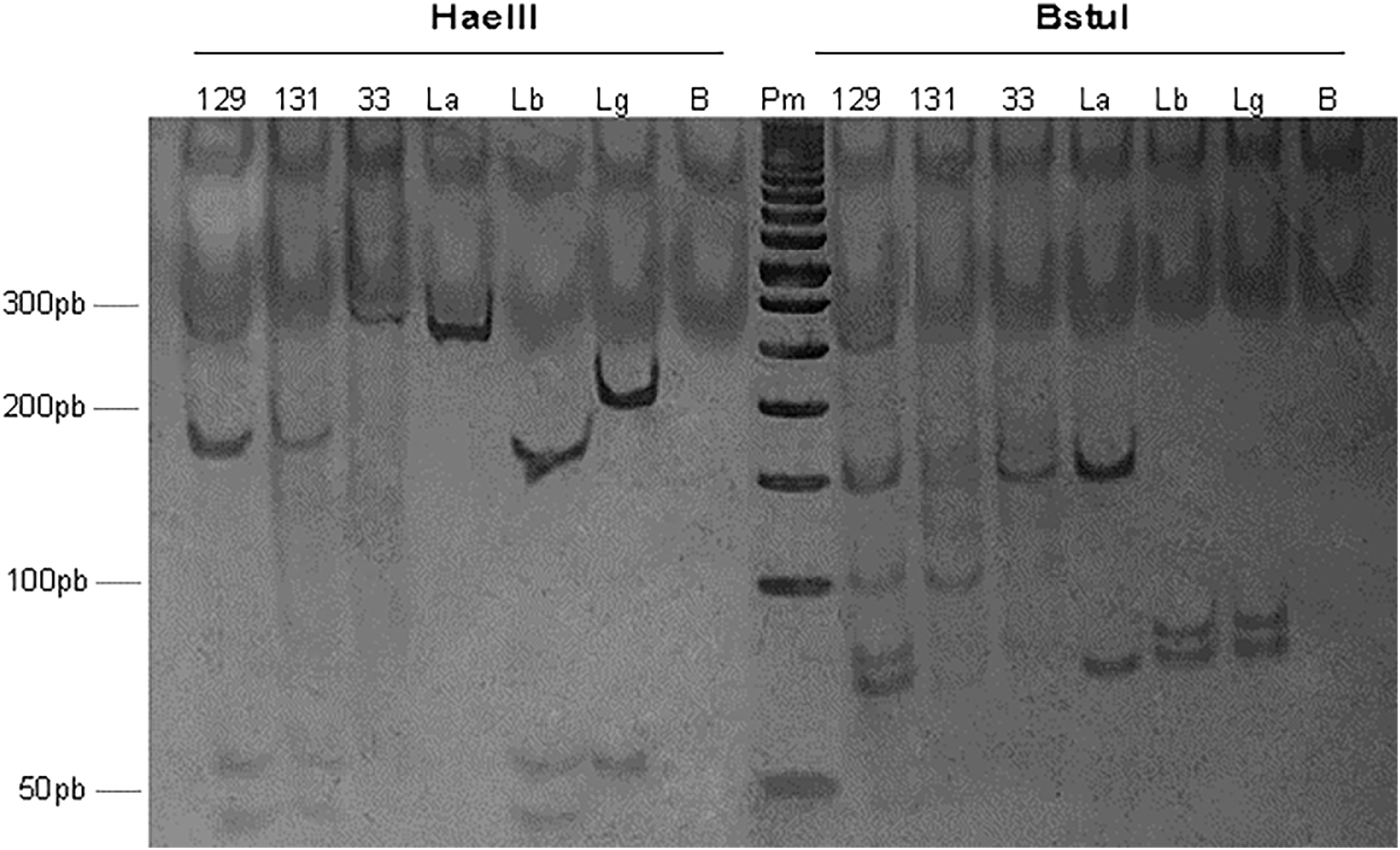
Enzyme digestion profiles of HaeIII and BstUI from pools 33, 129, and 131 in 12% polyacrylamide gel stained with silver nitrate. 129: Ps. amazonensis; 131: Ps. carrerai carrerai; 33: Ps. davisi; La: Le. (Le.) amazonensis; Lb: Le. (Vi.) braziliensis; Lg: Le. (Le.) guyanensis; B: negative control; Pm: molecular pattern.
Discussion
Currently, 131 sand fly species had been recorded in Rondônia Sate (47.50% of the species described in Brazil) (Galati 2016). A total of 52 species were identified in this study. Although Rondônia State is experiencing a substantial increase in disordered human occupation (Fearnside 2015) in the form of agricultural activities and the construction of settlements and hydroelectric power plants, sand fly species richness remains high and an increasing number of species continue to be found (Gil et al. 2003, 2009, Teles et al. 2013, Galardo et al. 2015, Resadore et al. 2017).
Of the areas examined in this study, the Jamari FLONA has been impacted by anthropization the least and it exhibited the greatest fauna richness (47 species), and the greatest diversity, confirmed by both Shannon and Simpson diversity indexes (Fig. 2), and it exhibited the second highest abundance of sand flies. This result corroborates the results of other studies that have shown that sand flies are sensitive to environmental changes since they consistently exhibit greater abundance and diversity in natural environments that have been well preserved. In areas with hydroelectric power plants that border the states of Amapa and Para, sand flies exhibited lower diversity in environments where anthropic activity was more invasive (Furtado et al. 2016). Conversely, sand fly richness and diversity have been observed to be greatest in forested areas where anthropic activity has had the least impact (Rosário et al. 2017). In general, in the Amazon, sand fly diversity is greatest in forested areas untouched by humans (Dias-Lima et al. 2002, Barbosa et al. 2008).
In this study, the most abundant species were Ps. chagasi s.l. and Ny. yuilli yuilli (42.15% of total collected), followed by Ps. davisi and Ny. antunesi (19.38%). Ps. chagasi s.l. has been observed as the most abundant species in other studies conducted in Amazonas State (Castellón et al. 2000); however, Ps. chagasi s.l. has been found in low abundance in studies conducted in Rondônia State: Biancardi et al. (1982), Gil et al. (2009), and Resadore et al. (2017) recorded its abundance at 0.74%, 0.37%, and 3.50%, respectively. Ny. yuilli yuilli was the second most abundant species in this study, yet it has been found in low abundance in the Amazon region; in Amazonas State, Castellon et al. (1994), Castellon et al. (2000), and Pereira-Júnior et al. (2015) recorded its abundance at 1.24%, 1.12%, and 4.57%, respectively; in Rondônia, Gil et al. (2009) recorded its abundance at 3.3%; and in Acre, its abundance has been recorded at 1.0% and 10.8% by Azevedo et al. (1993) and Teles et al. (2016), respectively. Ps. davisi was the third most abundant species in this study and it has been found in high abundance (between 21.07% and 39.63%) in studies performed in the states of Rondônia (Gil et al. 2003), Acre (Teles et al. 2016), and Pará (Souza et al. 2010). In this study, Ny. antunesi was most abundant in Santo Antônio, the area where anthropic activity has been most invasive, and Ny. antunesi abundance has been associated with peridomiciliary and intradomiciliary environments (Ramos et al. 2014, Pereira Júnior et al. 2015). Ny. antunesi has also been recorded in high abundance (between 16.92% and 25.47%) in other studies conducted in Rondônia (Biancardi et al. 1982, Teles et al. 2013, Resadore et al. 2017).
Higher abundance (5659 specimens) and greater richness (50 species) were observed in canopies (1039 specimens, 37 species), but the Shannon and Simpson indexes indicated that greater diversity was present at ground level. Other studies carried out in the Amazon region corroborate this finding: Castellon et al. (1994) identified 53 species in the canopy and 43 at ground level; Arias and Freitas (1982) identified 53 species in the canopy and 43 at ground level; and Arias and Freitas (1977) identified 21 species in the canopy and 17 at ground level. However, some studies have observed greater richness at ground level whereas others have observed relative parity between strata (Castellón et al. 2000, Feitosa and Castellón 2006, Furtado et al. 2016). Some species that were recorded only at ground level are species that have been incriminated as vectors or are thought to participate in the TL transmission cycle; these species include: Psychodopygus complexus, Ny. whitmani, Migonemyia migonei, and Psychodopygus paraensis (Silveira et al. 1991, Ready 2013). Feitosa and Castellon (2006) observed that some species are capable of changing their behavior by adapting their food preferences to available sources. In this context, female sand flies are found with greater abundance in stratum where blood sources are present; for example, Bi. flaviscutellata females feed on blood from ground dwelling rodents of the genera Oryzomys, Proechmys, and Rattus (Lainson and Shaw 1968, Caldart et al. 2017). Conversely, some sand fly species prefer blood sources found in the canopy; Ny. umbratilis and Ny. whitmani have been shown to feed on the blood of sloths (Bradypus) and anteaters (Tamandua) (Degener et al. 2014). It is noteworthy that most sand fly species change strata at night. This is probably due to their circadian cycle: sand flies hide in dark places during the day and become active at night (Campos et al. 2017).
This is the first study in the Amazon region to use BG-Sentinel CO2 traps. CDC light traps were more effective however, accounting for >70% of the specimens collected (n = 4871). The BG-Sentinel trap is widely used to capture mosquitoes of the genera Anopheles, Culex, and Aedes (Agha et al. 2017). Although CO2 is what attracts hematophagous insects to their bloodmeal sources (Gillies 1980), this study found that CO2 alone attracted sand flies less effectively than light, but conclusions about these two methods compared are imprecise. In the northern region of Africa, some studies report a variable fauna in relation to the capture of sand flies with CDC and BG-Sentinel traps. Hoel et al. (2010) observed more sand flies attracted to BG Sentinel. However, Obenauer et al. (2012) captured more sand flies in CDC than in BG Sentinel. Hence, more studies using these two methods of capture are necessary.
The natural infection rate of sand flies by Leishmania was 0.28%. This rate is highest in the study performed by Resadore et al. (2017) that recorded an infection rate of 0.23% in Porto Velho, Rondônia state. When compared with other studies conducted in the Amazon region, the rate found in our study is lower: in Acre, Teles et al. (2016) recorded 0.99%; in Amazonas, Pereira-Júnior et al. (2015) recorded 0.83%; and in Columbia, Trujillo et al. (2013) recorded 7.5%. According to Paiva et al. (2007), variability among recorded infection rates may be the result of differences between the various methodologies used for detection and identification of Leishmania.
In this study, Th. auraensis had the highest infection rate (8.33%). In Peru, Valdivia et al. (2012) identified Le. (Vi.) braziliensis and Le. (Vi.) lainsoni in samples of Th. auraensis with an infection rate of 0.6%; in Acre State, Teles et al. (2016) identified Le. (Vi.) braziliensis in Th. auraensis. In this study, Ps. amazonensis had the second highest infection rate (4.35%), and Le. (Vi.) braziliensis was identified in one pool of Ps. amazonensis, which makes this the first record of Ps. amazonensis naturally infected by Leishmania in Rondônia. Ps. davisi had the third highest infection rate (0.76%), and Le. (Vi.) amazonensis was identified in one pool of Ps. davisi. The presence of infection in Ps. davisi corroborates previous findings in Rondônia and this demonstrates Ps. davisi's importance as a probable TL vector within the state, especially since Ps. davisi has been found in high abundance throughout the Amazon region (Gil et al. 2003, Galardo et al. 2015, Teles et al. 2016). DNA of Le. (Vi.) braziliensis was detected in Ps. chagasi s.l., with an infection rate of 0.27%; tests using monoclonal antibodies uncovered similar infection rates in Para (Ryan et al. 1987). Some studies in the Amazon region have recorded sand flies naturally infected by Le. (Vi.) braziliensis, particularly sand flies of the genus Psychodopygus (Rangel et al. 1985, Grimaldi et al. 1991).
Although the annual infection rate of TL is high in Rondônia, the transmission dynamics of TL remain poorly understood; this is largely due to our ignorance of the participating vectors. Arias et al. (1985) found Ny. yuilli yuilli infected with Leishmania but did not identify the relevant strata (although it is known that this species has a preference for sloths). Gil et al. (2003) recorded Ps. davisi in high abundance and found specimens naturally infected with flagellated forms of Leishmania, identified as Leishmania naiffi. Ogawa et al. (2016) identified Leishmania DNA in pools of Th. auraensis, Nyssomyia Shawi, and Ny. antunesi. Resadore et al. (2017) identified Leishmania DNA in Th. ubiquitalis, and Th. ubiquitalis was the second most abundant species in that study, which suggests that this species is a probable vector of Leishmania in the municipality of Porto Velho. All the species mentioned in this study were found in abundance except for Ny. shawi. High abundance and the presence of Leishmania DNA in species such as Ps. davisi and Th. auraensis indicate that these species may be acting as vectors in the TL cycle in Rondônia State.
Footnotes
Acknowledgments
We thank Marlon F. Simplício, Bruno R. de Souza, Anne C.A. Meirelles and Lucas R. da Silva for their collection support. This research was performed using grant by the Conselho Nacional de Desenvolvimento Científico e Tecnológico (MCTI/CNPq No. 14/2013, process numbers 477490/2013-1 and 400238/2015-2).
Author Disclosure Statement
No conflicting financial interests exist.