
Abstract
Ticks are blood-sucking ectoparasites that transmit zoonotic pathogens to humans and animals. Ticks harbor not only pathogenic microorganisms but also endosymbionts. Although some tick endosymbionts are known to be essential for the survival of ticks, their roles in ticks remain poorly understood. The main aim of this study was to isolate and characterize tick-borne microorganisms from field-collected ticks using two arthropod cell lines derived from Ixodes scapularis embryos (ISE6) and Aedes albopictus larvae (C6/36). A total of 170 tick homogenates originating from 15 different tick species collected in Japan were inoculated into each cell line. Bacterial growth was confirmed by PCR amplification of 16S ribosomal DNA (rDNA) of eubacteria. During the 8-week observation period, bacterial isolation was confirmed in 14 and 4 samples using ISE6 and C6/36 cells, respectively. The sequencing analysis of the 16S rDNA PCR products indicated that they were previously known tick-borne pathogens/endosymbionts in three different genera: Rickettsia, Rickettsiella, and Spiroplasma. These included four previously validated rickettsial species namely Rickettsia asiatica (n = 2), Rickettsia helvetica (n = 3), Rickettsia monacensis (n = 2), and Rickettsia tamurae (n = 3) and one uncharacterized genotype Rickettsia sp. LON (n = 2). Four isolates of Spiroplasma had the highest similarity with previously reported Spiroplasma isolates: Spiroplasma ixodetis obtained from ticks in North America and Spiroplasma sp. Bratislava 1 obtained from Ixodes ricinus in Europe, while two isolates of Rickettsiella showed 100% identity with Rickettsiella sp. detected from Ixodes uriae at Grimsey Island in Iceland. To the best of our knowledge, this is the first report on successful isolation of Rickettsiella from ticks. The isolates obtained in this study can be further analyzed to evaluate their pathogenic potential in animals and their roles as symbionts in ticks.
Introduction
Ticks are important vectors among blood-sucking ectoparasites that transmit various zoonotic pathogens to humans and animals through their bite. Ticks harbor not only pathogenic microorganisms of veterinary and medical importance (Jongejan and Uilenberg 2004) but also several endosymbionts of the genera Coxiella, Francisella, and Rickettsia (Paddock et al. 2004, Ahantarig et al. 2013). The recent development of deep sequencing technologies has enabled high-throughput screening of pathogens and symbionts in ticks and expanded our knowledge on the diversity of microorganisms harbored by ticks (Nakao et al. 2013a, Qiu et al. 2014, Kurilshikov et al. 2015). Although these studies have led to the discovery of several previously unexpected or poorly characterized microorganisms, it is challenging to evaluate their roles in ticks and their pathogenic potential to animals solely based on their partial genome sequences.
In Japan, several tick-borne human diseases have been recognized. Until the recent emergence of severe fever with thrombocytopenia syndrome (Takahashi et al. 2014) and the reemergence of tick-borne encephalitis (Yoshii et al. 2017), most cases of tick-borne human diseases have been associated with bacterial infections. In particular, Japanese spotted fever caused by Rickettsia japonica is the most common tick-borne human diseases with hundreds of cases reported annually (National Institute of Infectious Diseases 2017). Several other rickettsioses caused by Rickettsia heilongjiangensis, Rickettsia helvetica, and Rickettsia tamurae have also been reported to date (Noji et al. 2005, Ando et al. 2010, Imaoka et al. 2011). The etiological agents of these rickettsial diseases were isolated from patients and ticks primarily using L929 mouse fibroblast cells (Uchida et al. 1992, Fournier et al. 2002, Fujita et al. 2006, Mahara 2006, Ando et al. 2010, Andoh et al. 2014). However, none of these studies has used arthropod cells for isolation of tick-borne pathogens in Japan.
At present, tick cell lines are indispensable tools to study the interaction between ticks and tick-borne microorganisms, including pathogens and symbionts in vitro (Bell-Sakyi et al. 2007, 2012). The cells have also been successfully used for isolating and propagating a number of tick-borne microorganisms (Bell-Sakyi et al. 2007, 2015). For example, the previously unculturable Borrelia lonestari was isolated and propagated for the first time in a tick cell line (Varela et al. 2004). Similarly, other tick-borne pathogens from genera such as Anaplasma and Ehrlichia have been successfully cultivated and maintained in tick cell lines (Munderloh et al. 2003, Zweygarth et al. 2013).
The main aim of this study was to isolate and characterize tick-borne microorganisms from field-collected ticks using two arthropod cell lines derived from Ixodes scapularis embryo (ISE6) and Aedes albopictus larvae (C6/36). Our approach led to the isolation of four previously validated rickettsial species, one uncharacterized rickettsial genotype and two tick endosymbionts, Rickettsiella and Spiroplasma.
Materials and Methods
Tick samples
This study employed unfed ticks collected in 11 different prefectures by a flagging method between 2013 and 2015 (Table 1). Tick species were identified morphologically under a stereomicroscope using standard keys (Nakao et al. 1992, Yamaguti et al. 1971). The samples included 15 different tick species: Amblyomma testudinarium (n = 10), Dermacentor taiwanensis (n = 3), Haemaphysalis concinna (n = 3), Haemaphysalis flava (n = 8), Haemaphysalis formosensis (n = 15), Haemaphysalis hystricis (n = 24), Haemaphysalis japonica (n = 9), Haemaphysalis kitaokai (n = 3), Haemaphysalis longicornis (n = 18), Haemaphysalis megaspinosa (n = 27), Ixodes monospinosus (n = 7), Ixodes nipponensis (n = 2), Ixodes ovatus (n = 9), Ixodes pavlovskyi (n = 2), and Ixodes persulcatus (n = 30). After identifying tick species, ticks were washed with 70% ethanol and sterile phosphate-buffered saline (PBS), and then homogenized in 100 μL of high-glucose Dulbecco's modified Eagle's medium (DMEM Gibco, Life Technologies, Carlsbad, CA) using a Micro Smash™ MS100R (TOMY, Tokyo, Japan). Half of the homogenate was subjected to DNA extraction using a blackPREP Tick DNA/RNA Kit (Analytikjena, Germany), while the other half was kept at −80°C and used for bacterial isolation. A total of 170 tick homogenates, including 158 from single unfed ticks (adult or nymph) and 12 from nymphal pools (5–24 nymphs/pool), were included in this study.
Collection Details of Ticks Used for Bacterial Isolation Using Arthropod Cells
−, this tick species was not collected in the sample year.
Nymphal pool samples were prepared from a pool of 5 to 24 nymphs.
The number in parentheses indicates the number of samples positive for rickettsiae by gtlA real-time PCR.
Maintenance of cell lines
ISE6 cells, originally reported by Kurtti et al. (1996) and received from the CEH Institute of Virology and Environmental Microbiology (Oxford, United Kingdom), were grown in L-15B medium supplemented with 10% fetal bovine serum, and 5% tryptose phosphate broth (Sigma-Aldrich, St. Louis, MO) at 32°C as described previously (Munderloh and Kurtti 1989), except that 0.1% bovine lipoprotein concentrate was not included in the culture medium. C6/36 cells, purchased from the American Type Culture Collection (No. CRL-1660), were grown in minimum essential medium (MEM; Gibco) supplemented with 10% fetal bovine serum, 2% MEM Non-essential amino acids (Gibco), 1% sodium pyruvate 100 mM (Gibco), and 1% L-glutamine (Gibco) at 28°C in a humidified atmosphere of 5% CO2 in air.
Co-culture with tick homogenates
ISE6 and C6/36 cells were seeded in 24-well culture plates and incubated overnight. On the following day, 5 μL of each tick homogenate was inoculated into separate wells of both cell lines. The culture medium was changed every 3 days for C6/36 cells and once a week for ISE6 cells. At 2 weeks postinoculation (pi), 100 μL of culture suspension was passaged into new wells containing uninfected cells. Second and third passages were conducted in the same way as first at 4 and 6 weeks pi, respectively. At 8 weeks pi, the experiment was terminated. All the bacterial isolates obtained in this study were preserved at −80°C for downstream analysis. Cell morphology was observed daily under an inverted microscope to detect cytopathic effects presumably caused by bacterial infections. When contamination of fungi or environmental bacteria was observed, the contaminated wells were sterilized with 10% hypochlorous acid for more than 10 min to prevent the spread of contamination to the neighboring wells.
Polymerase chain reaction
When the cells showed sign of bacterial infection, DNA was extracted from 100 μL of parent culture suspension and/or first subculture using a Wizard Genomic DNA Purification Kit (Promega, Madison, WI) following the manufacturer's instructions. In addition, cell suspensions from all wells at 4 and 8 weeks pi (from first and third subcultures, respectively) were subjected to DNA extraction to detect possible bacterial infection, whether or not morphological changes were seen in cells. PCR was conducted using the primers fD1 and Rp2 to amplify eubacterial 16S ribosomal DNA (rDNA) (Weisburg et al. 1991). To characterize rickettsial isolates, three additional genes were amplified: citrate synthase gene (gltA) (Gaowa et al. 2013), 190-kDa outer membrane gene (ompA) (Roux et al. 1996), and 120-kDa outermembrane protein gene (ompB) (Roux and Raoult 2000). All PCRs were conducted in a 25 μL reaction mixture containing 2.5 μL of 10 × KOD Plus Neo PCR Buffer, 0.5 μL of a high-fidelity KOD-Plus-Neo DNA polymerase (Toyobo), 200 nM of each primer, and 1.0 μL of template DNA. PCR conditions were as follows: 40 cycles of denaturation (94°C, 15 s), annealing (55°C for gltA, ompA, and 16S rDNA and 48°C for ompB, 30 s), and extension (68°C, 30 s for gltA, ompA, and ompB and 90 s for 16S rDNA). For some samples, we conducted TA-cloning using the pGEM-T vector (Promega) as described previously (Nakao et al. 2013b). The experimental procedures were approved by the Hokkaido University Safety Committee on Genetic Recombination Experiments (No. 2017-046). All the primer information is available in Table 2. All negative control containing sterile water instead of template DNA was included in all PCR assays.
Oligonucleotide Primers and Probe Used in Real-Time and Conventional Polymerase Chain Reactions and Sequencing
Real-time PCR
Real-time PCR to detect the gltA gene of spotted fever group and typhus group rickettsiae was conducted using the primers and probe shown in Table 2. Reaction mixtures were prepared using THUNDERBIRD Probe qPCR Mix (Toyobo, Osaka, Japan) and the reactions were carried out in a C1000 Thermal Cycler with a CFX96 Real-Time PCR Detection System (BioRad Laboratories, Hercules, CA) as described previously (Nakao et al. 2013b).
Sequencing and data analysis
All amplified PCR products were purified using a NucleoSpin Gel and PCR Clean-Up Kit (Takara Bio Inc., Otsu, Japan) and sequenced using the BigDye Terminator version 3.1 Cycle Sequencing Kit (Applied Biosystems, Foster City, CA). To sequence 16S rDNA PCR products, sequencing primers were newly designed in this study (Table 2). The purified sequencing products were analyzed on an ABI Prism 3130xl Genetic Analyzer Kit (Applied Biosystems) according to the manufacturers' instructions. The sequences from this study were submitted to the DNA Data Bank of Japan (
Results
Real-time PCR
Among 170 tick homogenates examined, 114 tested positive for Rickettsia infections by gltA real-time PCR (Table 1). We used both negative and Rickettsia-positive samples for the cell culture isolation experiments.
Isolation results
During the 8-week observation period, we confirmed bacterial isolation in 14 and 4 samples using ISE6 and C6/36 cells, respectively (Table 3). Ten isolates (nine from ISE6 and one from C6/36) were obtained in the parent cultures, while eight isolates (five from ISE6 and three from C6/36) were obtained in the first subcultures (Fig. 1). The sequencing analysis of the 16S rDNA PCR products indicated that they were previously known tick-borne bacteria in three different genera; Rickettsia, Rickettsiella, and Spiroplasma (Table 4). Although a further 4 and 11 samples in ISE6 and C6/36 cells, respectively, also showed bacterial growth in the well, sequencing analysis indicated the growth of environmental bacteria such as Bacillus spp., Pseudomonas spp., and Mycobacterium spp. (data not shown). Fungal infections developed in 75 ISE6 and 57 C6/36 wells; the remaining 77 and 98 wells, respectively, did not yield any isolates (Table 3). Although four rickettsial and three spiroplasmal isolates did not show any cytopathic effects in the infected cells, their infections were detected by PCR conducted at 28 days pi (Fig. 1).
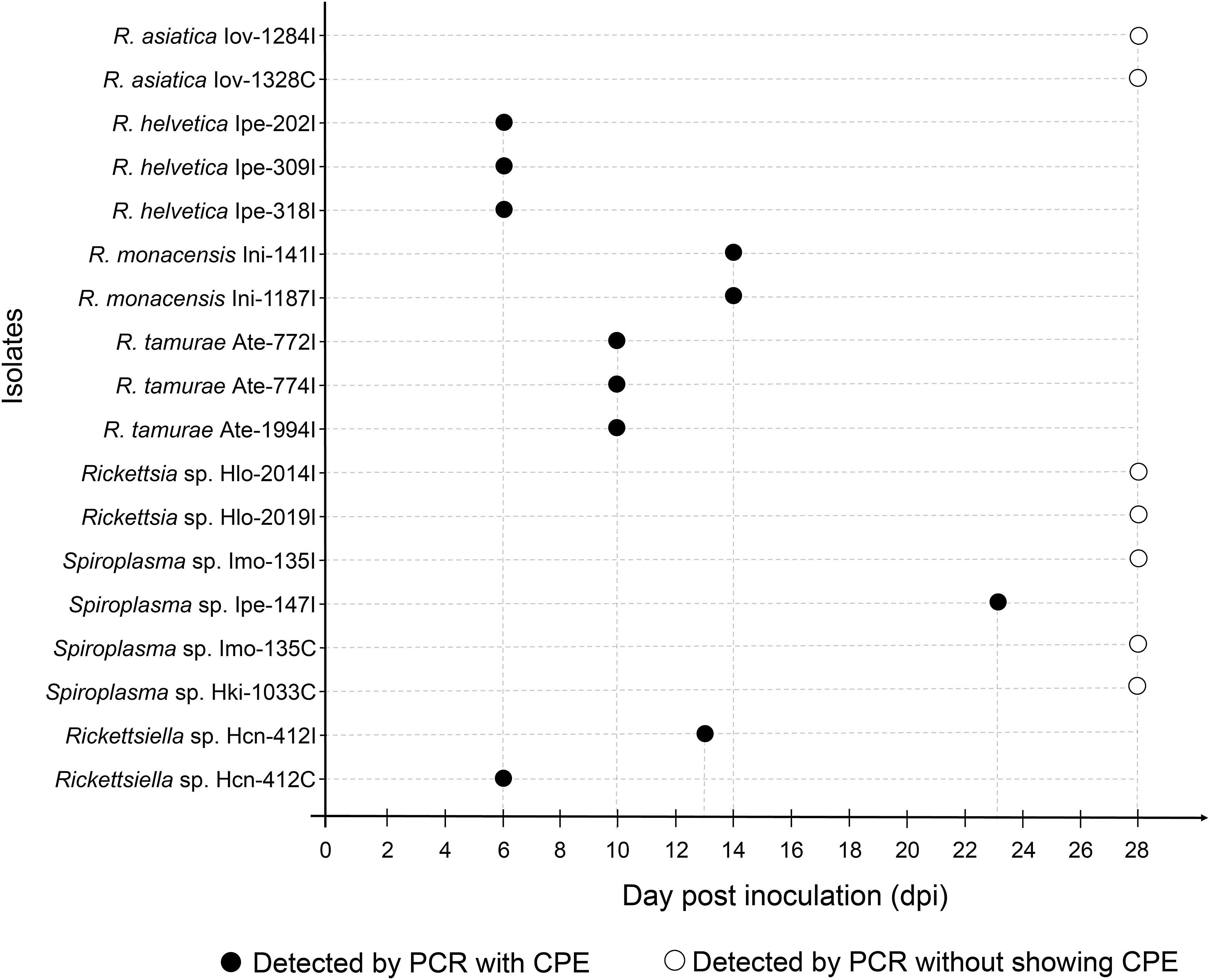
The time of onset of cytopathic effect observed in each bacterial isolate in ISE6 and C6/36 cells.
Summary of Bacterial Isolation from Homogenates of Ticks Collected in Japan Using Tick (Ise6) and Mosquito (C6/36) Cell Lines
Details of Ticks from Which Bacterial Isolates Were Obtained and the Results of Real-Time and Conventional Polymerase Chain Reactions Amplifying Bacterial Isolates
Screened by gltA real-time PCR using DNA extracted from tick homogenates.
PCR using DNA extracted from ISE6 and C6/36 cells.
+, positive; −, negative; NA, not applicable; dpi, day postinoculation.
Rickettsia isolation
Twelve isolates of Rickettsia (11 in ISE6 and 1 in C6/36) were obtained from different tick homogenates (Table 3). The amplification of gltA and ompB genes was successful in all rickettsial isolates, while the ompA gene was amplified only from seven isolates (Table 4). Based on the phylogenetic analysis of each rickettsial gene, the 12 isolates were identified as 4 previously validated species, Rickettsia asiatica (n = 2), R. helvetica (n = 3), Rickettsia monacensis (n = 2), R. tamurae (n = 3), and Rickettsia sp. LON, an unnamed rickettsial agent previously isolated from H. longicornis in Japan (Fujita 2008) (n = 2) (Fig. 2 and Supplementary Table S1). There was complete correspondence between rickettsial species/genotype and tick species of origin: R. asiatica, R. helvetica, R. monacensis, R. tamurae, and Rickettsia sp. LON was isolated from I. ovatus, I. persulcatus, I. nipponensis, A. testudinarium, and H. longicornis, respectively. A cytopathic effect was observed in R. helvetica-, R. monacensis-, and R. tamurae-infected cells at 6, 14, and 10 days pi, while there was no obvious morphological damage observed in R. asiatica- and Rickettsia sp. LON-infected cells (Fig. 1).

Phylogenetic tree of
Rickettsiella isolation
Rickettsiella was isolated from a homogenate of H. concinna collected in Hokkaido using both ISE6 and C6/36 cells (Table 1). A cytopathic effect was observed at 6 and 13 days pi in ISE6 and C6/36 cells, respectively (Fig. 1). The sequences of 16S rDNA of Rickettsiella obtained from two cell lines were identical and showed 100% identity with Rickettsiella sp. detected in Ixodes uriae from Grimsey Island in Iceland (GenBank No. KT697673). A phylogenetic analysis showed that our isolates formed a cluster with Rickettsiella spp. detected in pea aphids (Fig. 3 and Supplementary Table S1).

Phylogenetic tree of 16S rDNA of Rickettsiella endosymbionts isolated from ticks in Japan using maximum likelihood method. The sequences obtained in this study are shown in bold. GenBank accession numbers are given after species name.
Spiroplasma isolation
Four Spiroplasma isolates were obtained (two each from ISE6 and C6/36) (Table 3). Since two isolates were obtained from the same tick homogenate using different cell lines, four isolates originated from three tick homogenates: I. monospinosus, I. persulcatus, and H. kitaokai collected from Yamagata, Hokkaido, and Fukushima prefectures, respectively (Table 1). Only one isolate (Ipe_147I) showed a cytopathic effect in ISE6 cells at 23 days pi (Fig. 1). Among four isolates, two sequence types of 16S rDNA had two base pair (bp) differences in their sequences. One sequence type was obtained from two isolates (Imo_135I and Imo_135C) from I. monospinosus and one isolate (Hki_1033C) from H. kitaokai, while the other was from I. persulcatus (Ipe_147I). Both sequences showed the highest identities (1443/1444 bp and 1441/1444 bp) with 16S rDNA from Spiroplasma sp. detected from Fannia manicata (little housefly) (GenBank No. AY569829). In a phylogenetic analysis, our isolates were clustered together with 16S rDNA of two tick-derived Spiroplasma isolates: Spiroplasma ixodetis (GenBank No. NR104852) obtained from Ixodes pacificus and Spiroplasma sp. Bratislava 1 (GenBank No. KP967685) obtained from Ixodes ricinus, and several Spiroplasma spp. detected from various arthropods (Fig. 4 and Supplementary Table S1).
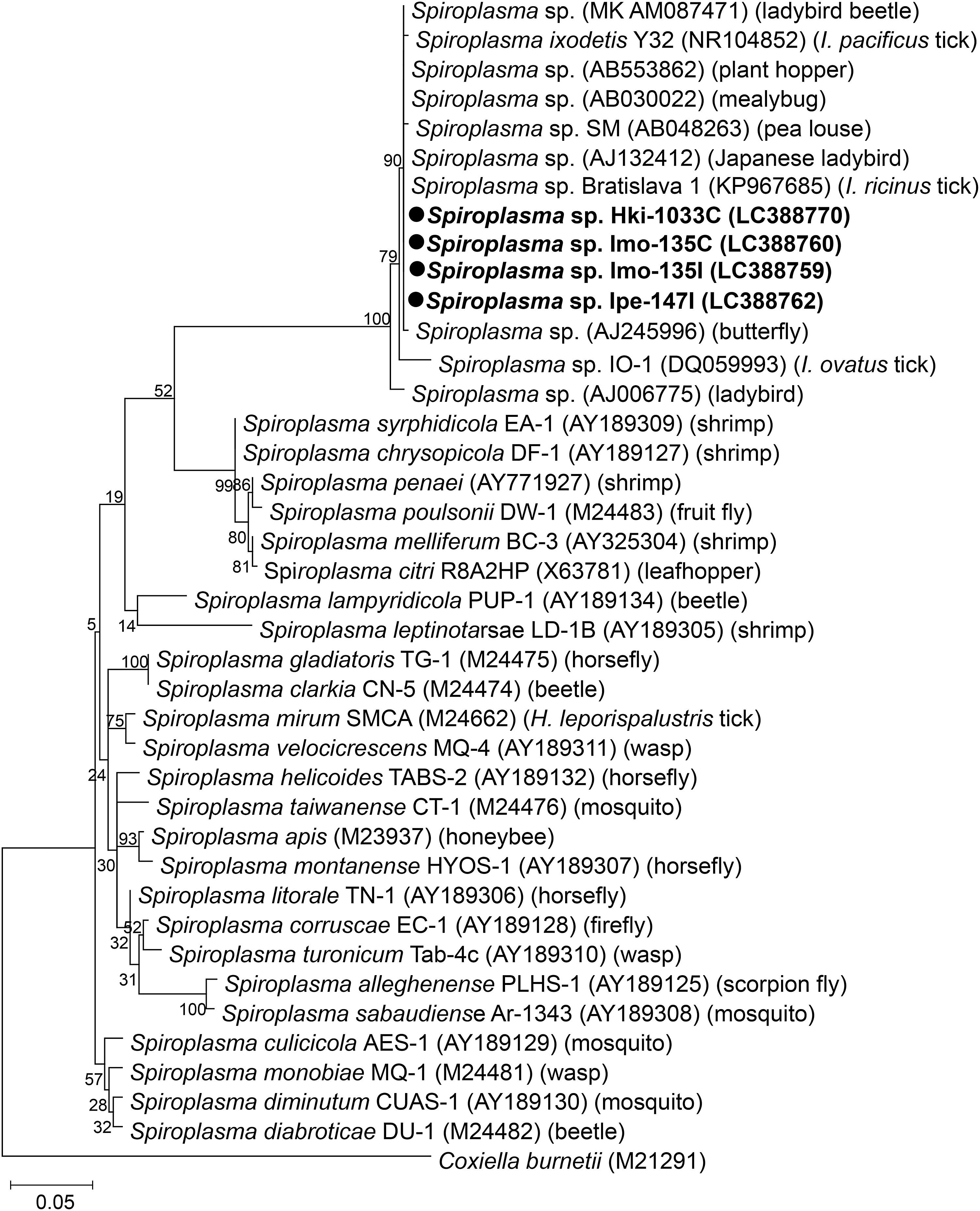
Phylogenetic tree of 16S rDNA of Spiroplasma endosymbionts isolated from ticks in Japan using maximum likelihood method. The sequences obtained in this study are shown in bold. GenBank accession numbers are given after species name.
Discussion
We achieved the first isolation and propagation of R. monacensis from Japan using ISE6 cells. This rickettsial agent has not been officially reported in Japan; in a recent nationwide survey, we found sequences of R. monacensis in I. nipponensis with high infection rates (Thu et al. 2019). R. monacensis was first isolated from I. ricinus collected in Germany using ISE6 cells (Simser et al. 2002), where the authors detected a cytopathic effect after the third passage (5 months). In our experiment, we observed a cytopathic effect as early as 14 days after the inoculation in both ISE6 and C6/36 cells (Fig. 1). Although the reason for this difference is not clear, it may suggest that phenotypes under in vitro culture conditions differ between R. monacensis strains. R. monacensis has been associated with a human rickettsiosis presenting Mediterranean spotted fever-like symptoms (Jado et al. 2007, Madeddu et al. 2012, Kim et al. 2017). This study reconfirmed the presence of this rickettsial agent in Japan and highlighted the necessity for further investigation of the clinical cases it may cause.
In addition to R. monacensis, we isolated three previously validated rickettsial species, R. asiatica, R. helvetica, and R. tamurae, and one uncharacterized genotype Rickettsia sp. LON, using ISE6 cells, among which only R. asiatica was also isolated in C6/36 cells (Table 4). Although the C6/36 cells inoculated with the tick homogenates from which Rickettsia were isolated using ISE6 cells were tested for rickettsial infections by PCR at 8 weeks pi, there were no positive amplicons. However, when the isolates of R. monacensis and R. helvetica obtained from ISE6 culture were inoculated into C6/36 cells in our preliminary experiments, their persistent growth in the cells was observed (data not shown). In fact, R. monacensis and R. helvetica were isolated from I. ricinus in Portugal using C6/36 cells in a previous study (Milhano et al. 2010). These results may indicate that lower success rates of rickettsial isolation using C6/36 cells are partly attributed to low bacterial burden in the inocula. It is also possible that propagation in ISE6 cells might help the rickettsial isolates adapt to in vitro culture conditions using C6/36 cells.
There are only a few reports on the in vitro propagation of Rickettsiella. For example, Rickettsiella grylli isolated from the variegated grasshopper, Zonocerus variegatus, was cultured in several cell lines derived from different arthropods (Henry et al. 1986). To the best of our knowledge, this is the first report of the successful isolation of Rickettsiella spp. from ticks using ISE6 cells and C6/36 cells. Bacteria within the genus Rickettsiella are known to be symbionts of many arthropods (Tsuchida et al. 2010, Leclerque et al. 2011, Iasur-Kruh et al. 2013, Łukasik et al. 2013). In ticks, Rickettsiella species have been reported in the genera Ixodes and Ornithodoros (Kurtti et al. 2002, Vilcins et al. 2009, Duron et al. 2015, 2016). A recent metagenomic approach based on 16S rDNA amplicons also showed the presence of Rickettsiella in the genus Haemaphysalis (Khoo et al. 2016). The sequence analysis of 16S rDNA revealed that the Rickettsiella sp. isolated from H. concinna showed 100% identity with a Rickettsiella endosymbiont detected in I. uriae collected from a seabird in Iceland (Fig. 3). These facts may support the hypothesis that Rickettsiella has a wide geographical distribution in ticks and is maintained by horizontal transfer between arthropod species as previously suggested (Duron et al. 2016). However, a more detailed analysis such as whole genome comparison between Rickettsiella spp. found in different arthropod hosts is essential to prove this hypothesis. The role of Rickettsiella in ticks is totally unknown; however, the lines of evidence from other arthropods indicated an effect on the survival of their host arthropods (Tsuchida et al. 2010, Łukasik et al. 2013). The isolate obtained in this study might be useful to further investigate potential roles of Rickettsiella in ticks.
We obtained four spiroplasmal isolates from three tick species: I. monospinosus, I. persulcatus, and H. kitaokai. The isolates obtained from I. monospinosus and H. kitaokai had completely identical 16S rDNA sequences and all spiroplasmal isolates in this study made one clade with previously reported Spiroplasma species, which were detected in a variety of arthropods, including ticks, ladybirds, plant hoppers, and mealybugs (Fig. 4). These findings may suggest that Spiroplasma is maintained by horizontal transfer between different arthropod species as suggested for Rickettsiella. This hypothesis should be explored in future studies. Most members of Spiroplasma are symbionts in arthropods and some of them are known to be beneficial to their hosts, for example, by protecting from fungal or parasitic infections (Łukasik et al. 2013, Xie et al. 2014, Yadav et al. 2018). In some arthropods, Spiroplasma species are pathogenic and cause gender ratio distortions known as a male-killing effect in Drosophila (Harumoto et al. 2014). However, a recent study conducted on a nidicolous tick Ixodes arboricola did not find any association between female-biased sex ratios and infections of six maternally inherited bacteria, including Spiroplasma (Van Oosten et al. 2018). Moreover, Spiroplasma mirum, an isolate obtained from the rabbit tick Haemaphysalis leporispalustris in the USA (Tully et al. 1982), was shown to have potentially pathogenic properties; for example, S. mirum was virulent for chick embryos and induced cataracts or lethal brain infections when introduced intracerebrally into experimental animals such as suckling rats and rabbits (Tully et al. 1977). Collectively, further biological characterization of Spiroplasma detected in this study is necessary to understand their roles in ticks and potential risks for human and animal health.
In this study, a number of wells were contaminated with bacterial or fungal infections (79 wells of ISE6 culture and 78 wells of C6/36 culture) (Table 3), despite the fact that tick surface was washed with 70% ethanol and sterile PBS. This result highlights the necessity of using additional chemicals to sterilize the tick surface, especially those effective against fungal infections. Another possible option might be to use only internal organs for bacterial isolation by dissecting ticks.
Conclusion
In conclusion, this study employed two arthropod cell lines to isolate pathogenic and symbiotic microorganisms from questing ticks collected in the field. Although the technique needs to be improved to reduce contaminations by fungal infections, the use of arthropod cell lines seems promising to expand our knowledge on microorganisms in ticks. The isolates obtained in this study are useful materials to further analyze their pathogenic potential in vertebrate animals and their roles as symbionts in ticks.
Footnotes
Acknowledgments
We would like to thank Dr. Ulrike Munderloh from the University of Minnesota and Dr. Kentaro Yoshii from Hokkaido University for providing ISE6 cells, and Dr. Yasuko Orba from Hokkaido University for providing C6/36 cells during the experiments. This work was financially supported by JSPS KAKENHI Grant-in-Aid for Young Scientists (B) (25850195 and 16K19112) and (A) (15H05633) and for Scientific Research on Innovative Areas (16H06429, 16K21723 and 16H06431).
Author Disclosure Statement
The authors declare no conflict of interests.
Supplementary Material
Supplementary Table1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.