
Abstract
Arthropod vector feeding preferences are defined as an overutilization of a particular host species given its abundance in relationship to other species in the community. Numerous methods exist to quantify vector feeding preferences; however, controlled host choice experiments are generally an underutilized approach. In this report, we present results from controlled vector host choice experiments using Culex quinquefasciatus Say (Diptera: Culicidae) mosquitoes and wild avian hosts identified as important contributors to West Nile virus (WNv) transmission in Atlanta, Georgia, United States. In each experiment, we allowed lab-reared F 1 Cx. quinquefasciatus to feed freely overnight on two avian individuals of a different species (i.e., northern cardinals, American robins, blue jays, brown thrashers, and gray catbirds). We then estimated WNv transmission potential using vectorial capacity and R 0. We found that mosquito blood feeding success was extremely variable among experimental replicates and that patterns of host choice only occasionally aggregated to a particular bird species. Vectorial capacity was highest for American robins and blue jays due to these species' higher reservoir competence for WNv and greater probabilities of mosquito selection of these species. Despite species-specific differences in vectorial capacity, total community capacity was similar among species pairs. R 0 estimates were qualitatively similar to capacity, and R 0 was below and above unity across species pairs. Our results provide empirical evidence that C. quinquefasciatus is an opportunistic blood feeder and highlight how variability in vector–host contact rates as well as host community composition can influence the likelihood of WNv transmission in avian communities.
Introduction
In recent decades, mosquito blood-feeding ecology has become an integral component of the study of mosquito-borne diseases. This is because field evidence has revealed extreme variability in mosquito bloodmeal choice, which theoretical models indicate can significantly modulate the incidence of disease in host communities (Woolhouse et al. 1997). Improvements in molecular methods and cost reductions have also influenced the growth of mosquito bloodmeal analyses as important investigative tools. Arguably the most common method for quantifying mosquito host choice is molecular identification of bloodmeals from passive or active collection of mosquitoes in the field (Silver 2008). When paired with estimates of host abundance at the time and place near when feeding occurs, bloodmeal analyses of field collections can identify patterns of host species under- and overutilization (Hess et al. 1968); an overutilization of a particular host species as a bloodmeal source may indicate a possible mosquito host preference. Although this is a common and well-accepted methodological framework, identifying mosquito host preferences from field-captured mosquitoes does have its limitations: suitable hosts are heterogeneously available in space and time (Kelly and Thompson 2000), and mosquito host selection patterns are influenced by the scale at which they are quantified (Chaves et al. 2010). Mosquito feeding success is also influenced by mosquito density (Edman et al. 1972) and host defensive behaviors (Edman and Kale 1971), which are difficult to quantify in passive field studies.
Since the invasion of West Nile virus (WNv) into North America, WNv has become a model system for understanding the impact of heterogeneous vector–host contacts on pathogen transmission among a broad range of potential host species. WNv is a zoonotic arbovirus primarily transmitted by Culex pipiens complex mosquitoes (Culex pipiens pipiens Linnaeus in northern latitudes and Culex quinquefasciatus Say in southern latitudes) among birds (Andreadis 2012). Patterns of C. pipiens complex host selection vary among the complex's members as well as across the United States, with C. pipiens pipiens considered a more ornithophilic vector than C. quinquefasciatus (Molaei et al. 2012). Previous research has shown that C. pipiens pipiens host choice is influenced by host availability, genetic heritage, and intrinsic vector feeding preferences (Kilpatrick et al. 2006, Hamer et al. 2009, Huang et al. 2009). Experimental host attraction studies suggest that C. pipiens pipiens individuals are more likely to feed on American robins (Turdus migratorius) than house sparrows (Passer domesticus) and European starlings (Sturnus vulgaris) (Simpson et al. 2009). Additionally, field and theoretical studies from the eastern and midwestern regions of the United States suggest that American robins are an overutilized bloodmeal host of C. pipiens pipiens in relation to other available hosts (Kilpatrick et al. 2006, Hamer et al. 2011, Simpson et al. 2012). Because American robins are considered reservoir competent for WNv (Komar et al. 2003), overutilization of this species may explain the timing and intensity of WNv epizootics and epidemics throughout the region (Kilpatrick 2011).
Throughout the southern portion of the United States, C. quinquefasciatus feeds on both birds and mammals (Farajollahi et al. 2011). In the southeast United States, C. quinquefasciatus feeds on a variety of peridomestic avian species such as northern mockingbirds (Mimus polyglottos), northern cardinals (Cardinalis cardinalis), American robins (Turdus migratorius), common grackles (Quiscalus quiscula), and gray catbirds (Dumetella carolinensis); the subspecies also occasionally feeds on mammals (Savage et al. 2007, Levine et al. 2016). C. quinquefasciatus feeding preferences may also change during a transmission season. In Atlanta, Georgia, American robins were identified as an overutilized bloodmeal host of C. quinquefasciatus during early summer, while northern cardinals were primary bloodmeal hosts during late summer (Levine et al. 2016). A similar seasonal shift in C. quinquefasciatus feeding patterns was recently identified in College Station, Texas, although from northern mockingbirds to northern cardinals (Komar et al. 2018). Because both northern cardinals and northern mockingbirds are considered moderately competent reservoir hosts for WNv (Komar et al. 2003, 2005), preferences for and/or shifts to these two hosts by C. quinquefasciatus could partially explain the lower incidence of WNv spillover in human populations in certain regions of the southern United States (Levine et al. 2016).
As a complement to the field study of WNv by Levine et al. (2016), we designed a controlled blood feeding experiment to quantify C. quinquefasciatus feeding success and host selection for five regionally important hosts of WNv in the southeast United States: American robins, blue jays (Cyanocitta cristata), brown thrashers (Toxostoma rufum), gray catbirds, and northern cardinals. By employing a direct host choice experimental design, we controlled for some of the limitations of bloodmeal field analyses and provide a complimentary approach to understanding the complexities of C. quinquefasciatus host choice. Our null hypothesis was that C. quinquefasciatus would feed opportunistically among available hosts and there would be no detectable preference for any particular species. We then utilized our experimentally derived host choice results to estimate WNv transmission potential and intensity in our experimental host combinations using two metrics of transmission, vectorial capacity (C) and the basic reproductive number of the pathogen, R 0.
Materials and Methods
All procedures were approved by Emory University's Institute for Animal Care and Use (DAR-2002351). Bird collection permissions were granted by local and federal bird collection permits (GA DNR 29-WJH-14-90 and USGS 23673).
Study design
An aviary was constructed with ½″ PVC piping and mosquito netting glued to the pipes, measuring 1.75 × 0.75 × 0.75 meters (Fig. 1). The interior of the aviary allowed two 0.33 × 0.28 × 0.41-meter metal birdcages (Prevue Pet Products) with 0.013 meters of wire spacing to be inserted within so that birds remained confined, but could actively defend themselves from mosquito attacks. One side of the aviary's netting was held to the structure with detachable adhesives so that the aviary could be opened to add/remove the birdcages. A cloth stockinette was also attached to one side of the cage so that a mosquito-proof seal could be maintained when adding/removing birds and mosquitoes. The aviary was placed lengthwise on top of a plastic folding table and kept underneath an outdoor pavilion for all experiments.

Experimental set up.
Culex quinquefasciatus mosquitoes were reared from field-collected egg rafts obtained in Atlanta, GA, between July and September 2010, 2011, and 2013 using pans baited with a 1 gallon mixture of water, hay, and dog food (Chaves et al. 2009). Larvae emerging from eggs were reared at 30°C and fed 5 g of yeast (Fleishmann's; ACH Food Companies, INC., Memphis, TN) or 5 g of crushed Koi food pellets (Tetra Pond; Tetra Holding, INC., Blacksburg, VA) daily. Pupae were removed to an emergence chamber and adults were provided a 10% sugar/water solution. Female mosquitoes used in host choice assays were at least 3 days postemergence and were deprived of the sugar solution ∼8 h before each experiment.
Each experiment consisted of pairing a northern cardinal with an American robin, blue jay, brown thrasher, gray catbird, or another northern cardinal. Collections attempted to collect two individuals of a different species. However, because northern cardinals were the most commonly captured species, and often the only captured species during a collection event, we had to limit our experimental combinations to the inclusion of at least one northern cardinal and one of the species listed above (experimental group) or another northern cardinal (control group). Birds were captured with 35-mm mesh polyester mist nets in a study site near Emory University's campus in Atlanta, GA. Captured birds were aged, sexed, weighed, and banded when banding equipment was available. Birds used for experiments were housed on-site in metal birdcages with water, a wooden perch, and either 100 grub worms or 50 g birdseed, depending on each species' diet. Cages were placed in a dark and ventilated location until the experiment was conducted (the evening of the mist-netting day).
Thirty minutes after official sunset time, the metal cages containing the birds were placed on the floor of the aviary, spaced ∼0.5 meters apart (Fig. 1). After 5 min of acclimation, a cup containing 30 female mosquitoes was inserted into the enclosure using the stockinette entrance and mosquitoes were released at the release point in Fig. 1. Mosquitoes and birds were left in the aviary overnight, and the experiment ended at ∼6:30 AM the following morning. Birds were first released from their cages and aviary using the stockinette entrance and mosquitoes were then aspirated out of the enclosure with a Prokopack mosquito aspirator (Vazquez-Prokopec et al. 2009).
Bloodmeal analysis
Abdomens of all blood-fed mosquitoes were dissected and DNA was purified using a Qiagen DNeasy Blood and Tissue Kit (Qiagen, INC., Valencia, CA) following the manufacturer recommendations. A fragment of the 16s ribosomal gene was amplified by direct PCR (Killmaster et al. 2011), and products were run on a 1.5% ethidium bromide-stained gel and visualized under UV light. Successful PCR products were purified using a multiscreen plate purification kit (Millipore, Burlington, MA). DNA was then sequenced in forward and reverse directions with an ABI 3500 (Applied Biosystems, Foster City, CA) automated sequencer. Sequences were assembled and reconciled using DNA Star Seqman Pro (DNASTAR, INC., Madison, WI). A multisequence alignment with sample and control sequences was performed using BioEdit (Ibis Biosciences, Carlsbad, CA) and MEGA5 (Tamura et al. 2011).
The control reference sequences for each bird species were generated from blood clots from field-collected birds, which were blasted with 100% homology to sequences available in GenBank (except for brown thrasher, which was not previously available and which we submitted to GenBank [MH031275—this work]). Sequence comparisons were performed in a 500-bp fragment of the 16S ribosomal gene using 76 variable sites that unmistakably discriminated all the bird species assayed in this work. Specifically, 38 sites differentiated northern cardinals from American robins, 44 sites differentiated from blue jays, 36 sites from brown thrashers, and 41 sites from gray catbirds. A >98% base-pair match to one of the control sequences was used to determine which species the mosquitoes had fed on.
Data analyses
We utilized binomial, generalized, linear mixed-effect models (GLMMs) to compare C. quinquefasciatus feeding success and host choice among replicates. Because northern cardinals were used in all experiments, the reported outcomes for mosquito feeding success are attributed to the effect of the noncardinal hosts. For comparisons of host choice, cardinal–cardinal experiments were dropped from the analysis because northern cardinal was the only species available and our molecular methods could not distinguish individuals of the same species.
Comparisons were made at the level of each individual mosquito to minimize the effect of limited replication and to increase power in detecting differences in host choice (Chaves 2010). For comparisons of feeding success (i.e., a mosquito fed on any species or it did not), the average recorded temperature and humidity for the duration of each experiment and the noncardinal host were modeled as fixed effects with cardinal–cardinal experiments as the reference group and replicate as a random effect. Because we could not conduct a full factorial design of host species combinations, models comparing host selection (i.e., whether a mosquito fed on the blood of a northern cardinal or it did not) used only temperature and humidity as fixed effects and replicate as a random effect. Temperature and humidity were centered to the average value in the data set to improve model convergence. All models were compared using AIC, and models were considered similar when ΔAIC <2. Odds ratios for feeding success and host choice were calculated from the best-fitting GLMM. All GLMMs were implemented using the lme4 package for R (Bates et al. 2015), odds ratio plots were generated using the sjPlot package for R (Ludecke 2018), and all other statistical analyses were performed in R (R Development Core Team 2008).
Vectorial capacity and R 0
We used our predicted feeding success and host choice results to assess the impact of host choice on two measures of WNv transmission intensity for each host species combination: vectorial capacity (C) and the basic reproductive number of the pathogen, R 0. The two measures are inter-related, although each quantifies different aspects of mosquito-borne pathogen transmission. Vectorial capacity is often referred to as the daily reproductive rate of the pathogen and is the entomological component of R 0 in its simplest derivation (Smith and McKenzie 2004). We define vectorial capacity as the expected number of infected mosquitoes (or bites) from a susceptible population feeding on a single infected host per unit area and time. The utility of C is its ability to measure (or provide an estimate of) the transmission potential of a vector population in the absence of empirical evidence for transmission and without any information on infections in host(s) (Garrett-Jones and Shidrawi 1969). An additional attribute of C is that the model assumes independence of transmission between species, and total transmission C is the sum of each species' estimate (Smith and McKenzie 2004). R 0, which is the expected number of secondary infections arising from the introduction of a single infected individual into a fully susceptible population, is a more robust measure of transmission potential with widely accepted and established transmission thresholds: R 0 estimates above unity provide a theoretical indication that transmission pathways can be sustained, which is a signal of epidemic potential.
The formula for C
total is as follows:
where m = mosquito abundance in relation to species i, a = mosquito feeding rate on species i, P = mosquito nightly survival, EIP = WNv extrinsic incubation period, and h = probability that species i transmits WNv to a biting mosquito.
We calculated R
0 using the formula presented by Simpson et al. (2012). This derivation of R
0 was developed to investigate the impact of feeding preferences on WNv transmission and incorporates a feeding index of a preferred host in the equation. The formula is as follows:
where β v = vector-to-host transmission rate, β p = preferred host-to-vector transmission rate, β a = alternative host-to-vector transmission rate, ν = vector biting rate, N v = number of vectors, N a = number of alternative hosts, N p = number of preferred hosts, d v = vector death rate, d p = preferred host death rate, d a = alternative host death rate, α v = feeding index of the preferred host, γ a = recovery rate of alternative host, and γ p = recovery rate of preferred host.
For estimates of C, the biting rate term was predicted using the best host choice model under AIC and the bootMer function in the lme4 package (Bates et al. 2015). The other parameters of the equation were fixed: 14 mosquitoes per bird to reflect the average number of mosquitoes released in each experiment; survival probability of 0.9, which is similar to quantities used by Simpson et al. (2012); and the EIP was fixed at 14 days, which reflects previously published estimates for C. pipiens complex mosquitoes (Kilpatrick et al. 2008). The probabilities of mosquitoes acquiring infections from hosts (hi ) were based on published estimates by Komar et al. (2003, 2005). For estimates of R 0, the biting rate (ν) was estimated using the overall feeding success of mosquitoes per replicate (predicted from best-fitting feeding success model), while the feeding index was estimated as the ratio of noncardinal to cardinal host choice estimates from our host selection GLMMs. Estimates of the recovery rate of WNv in our experimental hosts were based on published reports of the WNv infectious period by Komar et al. (2003, 2005). Additionally, host death rates were assumed negligible. Because host competence and the infectious period of WNv for brown thrashers and gray catbirds have not been quantified, we used the value published for the closely related species, northern mockingbird.
Results
Thirty-four birds were captured and utilized for 17 replicates: 3 cardinal–robin, 2 cardinal–jay, 5 cardinal–thrasher, 3 cardinal–catbird, and 4 cardinal–cardinal combinations (Table 1). A total of 460 C. quinquefasciatus mosquitoes were used for all experiments with an average of 27 mosquitoes released per replicate; 387 (84.1%) were recaptured and 219 (56.6%) were fully engorged (considered a sign of a successful bloodmeal). No partial bloodmeals were observed. Bloodmeals from 154 mosquitoes were successfully sequenced and identified to a particular bird species. PCR products from 8 blood-fed mosquitoes did not provide a reliable sequence and 57 bloodmeals from cardinal–cardinal trials were not sequenced because northern cardinal was the only possible species to feed from. No mixed species bloodmeals were detected by PCR.
Culex quinquefasciatus Blood Feeding Success for Each Experimental Host Combination
Odds ratios were calculated using binomial, generalized, linear mixed-effect models for overall mosquito blood feeding success, and the reference group is listed within the table.
CI, confidence interval.
Feeding success ranged between 16.7% and 96% across replicates (Table 1). Two candidate models for feeding success were identified. The first model (Model 1) included humidity and the noncardinal species as fixed effects and replicate as a random effect. Under this model, humidity improved model fit, but was not a significant predictor of feeding success (F = 1.13, p > 0.05). Additionally, in Model 1, there was no statistically significant difference in feeding success between cardinal–robin, cardinal–jay, or cardinal–catbird combinations (Table 1). However, there was a significant difference in feeding success when the noncardinal host was a brown thrasher (odds ratio 0.22; 95% confidence interval 0.05–0.35) (Table 1). The alternate feeding success model by AIC included only replicate as a random intercept. Under this model, feeding success varied considerably among replicates, and the probability of successful feeding was significantly different from 0.5 for numerous replicates (Table 2). Model comparisons for host choice identified humidity as a fixed effect and replicate as a random effect for the best model. Under this model, humidity had a small but significant effect on host choice (F = 4.88, p = 0.03). The model additionally predicted a slight avoidance of cardinals, but only when the species pair was with a blue jay; all other estimates of host choice or avoidance did not differ from 0.5 (Table 2).
Random Intercept Effect Terms from the Alternate Feeding Success Model and the Host Choice Model
Feeding success was modeled using a binomial, generalized, linear mixed-effect model (GLMM) with only experimental replicate as a random intercept term. Host choice was modeled as a binomial error GLMM with humidity as a fixed effect and experimental replicate as a random intercept effect. Separate models predicted host choice on a cardinal and a noncardinal.
Estimates of C were largely driven by each species' WNv competence index rather than our model predictions of host choice. C robin and C jay estimates tended to be higher than C cardinal estimates and few confidence limits overlapped (Fig. 2A). Capacity estimates from cardinal–thrasher and cardinal–catbird combinations were more variable and only occasionally were significant differences observed by comparisons of confidence intervals (Fig. 2A). Despite sometimes large differences in C i estimates within species combinations, C total estimates were similar for cardinal–robin, cardinal–thrasher, cardinal–catbird, and cardinal–cardinal combinations (Fig. 2B). R 0 estimates were above unity for all cardinal-robin, cardinal-jay, and cardinal-cardinal replicates while unity was included in the 95% confidence interval for cardinal-thrasher and cardinal-catbird replicates when using predicted feeding success in Model 1 (Fig. 2C). R0 estimates were qualitatively similar when using the alternate feeding success model with the exception that estimates were more variable among species-pairs (Fig. 2D).
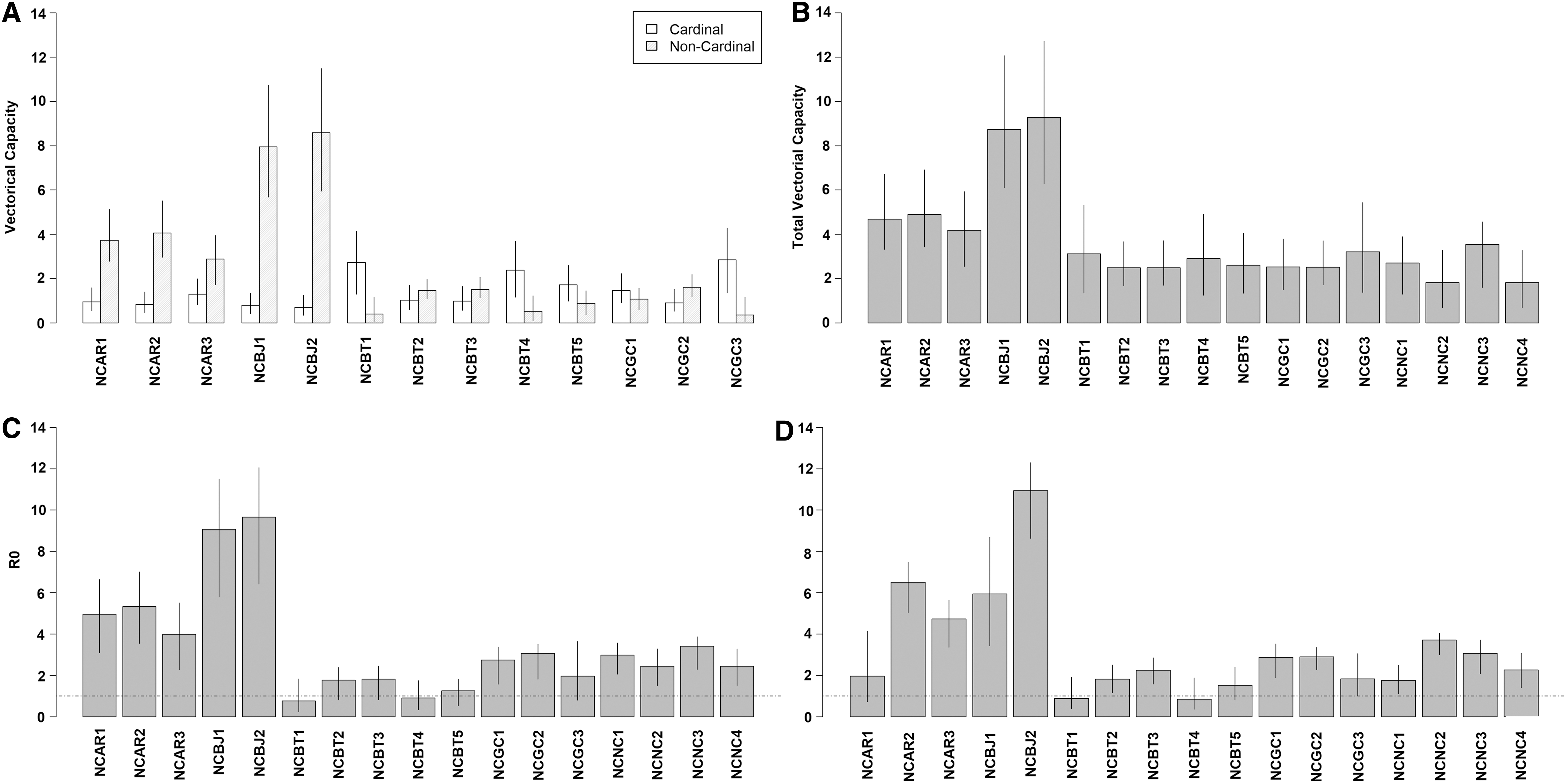
Vectorial capacity
Discussion
Our study was designed to complement a previous field study of C. quinquefasciatus blood feeding patterns in Atlanta, GA, which found American robins, northern cardinals, and members of the Mimidae family to be the most important bloodmeal hosts of C. quinquefasciatus mosquitoes (Levine et al. 2016). A key finding from that study was an observed shift in host selection by C. quinquefasciatus during the WNv epidemic period: mosquitoes fed preferentially on American robins early in a transmission season and then shifted to northern cardinals later in a season when peak virus transmission occurs (Levine et al. 2016). Shifts in mosquito feeding to low-competence species such as northern cardinals may explain reduced rates of WNv spillover in human populations throughout the southeast United States region (Levine et al. 2016).
Vector–host encounters depend on multiple factors, including the spatial distribution of birds or bird roosts (Diuk-Wasser et al. 2010, Benson et al. 2012, Janousek et al. 2014, Komar et al. 2015), spatial distribution and abundance of vectors (Thiemann et al. 2011, O'Brien and Reiskind 2013), and specific landscape characteristics (Vazquez-Prokopec et al. 2010). This may lead to heterogeneities in estimates of host utilization, depending on the sampling method and location, as well as time of the year. By controlling for variability in host availability and vector abundance, our experiments show that C. quinquefasciatus feeds opportunistically among available species rather than preferentially on a single species. Although species-specific estimates of vectorial capacity were different among species combinations, estimates of total capacity were similar, which indicates that WNv transmission could be equally intense among the different combinations of avian species used in this experiment. Our capacity estimates were also qualitatively similar to our estimates of R 0, which predicted that several combinations of host species were capable of sustaining WNv transmission. Several field studies have found that species other than American robins can contribute to the WNv transmission cycle, even if they are not the most preferred or competent species (Reisen et al. 2006, Levine et al. 2016, Komar et al. 2018). The similarity of our WNv transmission intensity estimates among multiple combinations of avian species may help explain why WNv epizootics occur in very different avian communities across the United States as well as in other countries. Further research is needed to determine the factors that ultimately influence successful bloodmeal acquisition and WNv amplification among C. quinquefasciatus and American robins, northern cardinals, and other abundant species in the southern United States.
Patterns of C. quinquefasciatus host choices vary widely across North America, and our data cannot directly address how the availability of other avian or mammal species alters the feeding patterns of C. quinquefasciatus mosquitoes. In a field study of WNv seroprevalence in small rodent populations in Atlanta, GA, antibodies to WNv were detected in eastern gray squirrels (Sciurus carolinensis), suggesting that WNv-infected C. quinquefasciatus mosquitoes may occasionally bite mammals (Bisanzio et al. 2015). Other field studies of C. quinquefasciatus blood feeding in the southern United States have also shown variable rates of C. quinquefasciatus blood feeding on mammalian hosts, including humans (Dennett et al. 2007, Molaei et al. 2007). Because C. quinquefasciatus is an opportunistic feeder, future experiments could be designed to directly test the likelihood that C. quinquefasciatus mosquitoes from the southeast United States feed opportunistically among avian and mammalian hosts, which could better inform the risk of WNv spillover in the region. Future experiments could also be designed to better isolate host preferences, such as by pairing a hypothesized preferred host species among numerous alternative host species and then quantifying host selection probabilities weighted by abundance.
An important limitation of our study was our inability to separate the magnitude of C. quinquefasciatus host preferences from avian defensive behaviors. However, previous studies have shown that host defensive behaviors do not significantly limit mosquito blood feeding success. Darbro and Harrington (2007) determined that house sparrows were commonly fed upon by C. pipiens pipiens mosquitoes despite elevated defensive behaviors (Darbro and Harrington 2007). A study by Edman et al. (1974) also did not observe mosquito host switching from highly defensive hosts to less defensive hosts (Edman et al. 1974). In addition to limitations of unquantified defensive behaviors, we cannot determine whether missing mosquitoes were eaten or not by hosts, although this is the most likely explanation for not recapturing all released mosquitoes. We also acknowledge the low statistical power of our experiments, which was due to the logistical difficulty of capturing specific wild host species in sufficient numbers to conduct all pair-wise combinations. While implementing the analysis using GLMMs partially addressed this limitation (Chaves 2010), we expect that findings from our study can further the research of mosquito host choice in variable host species combinations and its relationship with pathogen transmission.
Conclusions
Mosquito host preferences remain critically important to understanding WNv transmission and persistence. By experimentally manipulating host species availability, we quantified the individual feeding response of C. quinquefasciatus in specific two-host species combinations. Importantly, our study predicted (using a mathematical formulation for vectorial capacity and R 0) that generalist blood feeding on mildly competent birds is capable of generating transmission potential estimates similar to those derived from aggregated feeding on highly competent birds. This may partially explain the occurrence of WNv epizootics and epidemics in regions with different avian fauna. Additionally, our results provide further evidence that American robins and northern cardinals are important amplifying hosts of WNv in the southeast United States.
Footnotes
Acknowledgments
The authors thank their anonymous reviewers for their thoughtful comments and suggested improvements for the manuscript. This work was funded by departmental startup funds belonging to U.K. and G.V.P.
Authors' Contributions
J.R.M., U.K., D.M., and G.V.P. conceived the experiments; J.R.M., C.H., and G.V.P. performed the experiments; J.R.M. and P.L.M. conducted molecular analyses; J.R.M. analyzed all the data and wrote the first manuscript; and all authors contributed significantly to revisions of the manuscript. All authors read and approved the final version of the manuscript.
Availability of Data and Materials
Data can be made available upon request.
Author Disclosure Statement
No conflicting financial interests exist.