
Abstract
Introduction:
The prevalence of Francisella tularensis in Minnesota ticks is unknown. Ticks collected at seven sites were tested to determine the infection prevalence of F. tularensis in Dermacentor variabilis in Minnesota.
Materials and Methods:
Ticks were collected from two properties at an epizootic site and at five long-term tick research sites. Ticks were pooled by species, sex, date, and site with a maximum of 10 ticks per pool. Ticks were bisected and homogenized; DNA from supernatant was extracted and tested by real-time PCR (RT-PCR). Twice, additional ticks were collected for bacterial culture and isolation of F. tularensis. Proportion of positive pools and minimum infection rate (MIR) were calculated.
Results:
A total of 3527 ticks were tested for F. tularensis including 1601 male D. variabilis and 1926 female D. variabilis. Across all sites, 128 (34%) of 378 pools were RT-PCR positive for F. tularensis. Of 128 positive pools, F. tularensis from 96 (75%) was identified as type A; F. tularensis from 32 pools was unable to be subtyped. The overall MIR was 3.6%. The MIR was significantly lower at the epizootic site compared with Morrison County 1 (3.9% vs. 7.2%; p = 0.02) but did not differ between the epizootic site and Pine County 1 (3.9% vs. 2.1%; p = 0.49). Within the epizootic site, the MIR was significantly higher at Washington County 2 compared with the adjacent property (5.7% vs. 2.3%; p < 0.001). F. tularensis was cultured from 6 (15%) of 40 pools.
Conclusions:
F. tularensis was found in ticks at a majority of sites tested. The MIR of F. tularensis in D. variabilis ticks in Minnesota varied geographically. Our findings support the hypothesis that D. variabilis plays an important role in the natural history of tularemia in Minnesota. Further ecologic studies are needed to fully understand the importance of tick species in the maintenance and transmission of F. tularensis in Minnesota.
Introduction
Tularemia is a zoonotic disease caused by the bacterium Francisella tularensis. Two subspecies of F. tularensis are considered important for human health, F. tularensis tularensis (type A) and F. tularensis holarctica (type B) (Olsufiev et al. 1959). Both type A and type B infections occur in the United States, with isolate data available to the Centers for Disease Control and Prevention (CDC) suggesting that the majority of reported human tularemia cases are caused by type A (Staples et al. 2006, Kugeler et al. 2009). Tularemia ecology is complicated and not wholly understood in the United States; however, the type A cycle generally involves lagomorphs, whereas the type B cycle generally involves rodents and semiaquatic mammals (Farlow et al. 2005, Staples et al. 2006, Keim et al. 2007, Kugeler et al. 2009). Ticks are important in the enzootic maintenance of the bacteria (Gelman 1961, Shaughnessy 1963, Olsen 1975, Morner and Addison 2001, Friend 2006, Keim et al. 2007), and three species are considered important vectors for transmission to humans: Dermacentor andersoni (Rocky Mountain wood tick), Dermacentor variabilis (American dog tick), and Amblyomma americanum (lone star tick) (McDowell et al. 1964, Olsen 1975, Friend 2006, Keim et al. 2007, Petersen et al. 2009). Other arthropods, such as deer flies, have been found to be naturally infected, but their relative importance as vectors of tularemia in the United States is not well understood (Francis 1919, Cox 1965, Klock et al. 1973).
Tickborne tularemia in people was first described in the United States in 1923 in a patient with glandular fever (Jellison 1974), and in Minnesota in 1926 in a man who developed ulceroglandular tularemia following the bite of a D. variabilis tick (Hanson and Green 1929). In 1924, F. tularensis was first isolated from a tick during a study of D. andersoni ticks in Montana (Parker et al. 1924). Since then, ticks have been implicated in multiple outbreaks of tularemia (Warring and Ruffin 1946, Saliba et al. 1966, Schmid et al. 1983, Markowitz et al. 1985), and have been described as the most common source of transmission in both type A and type B sporadic tularemia cases across the United States (Staples et al. 2006).
Minnesota is not a high incidence state for tularemia; however, there has been an increase in human and animal case reports in recent years (Fig. 1). In Minnesota, from 1994 to 2017, 9 (43%) of 21 human tularemia cases were caused by F. tularensis type B, 4 (19%) were caused by type A, and 8 (38%) cases were not subtyped. Nine (43%) of the 21 cases had a likely tickborne exposure (as determined using a standard exposure interview); of the 5 tickborne cases with an identified F. tularensis subspecies, 3 (60%) were infected with type B and 2 (40%) were infected with type A (Fig. 2).
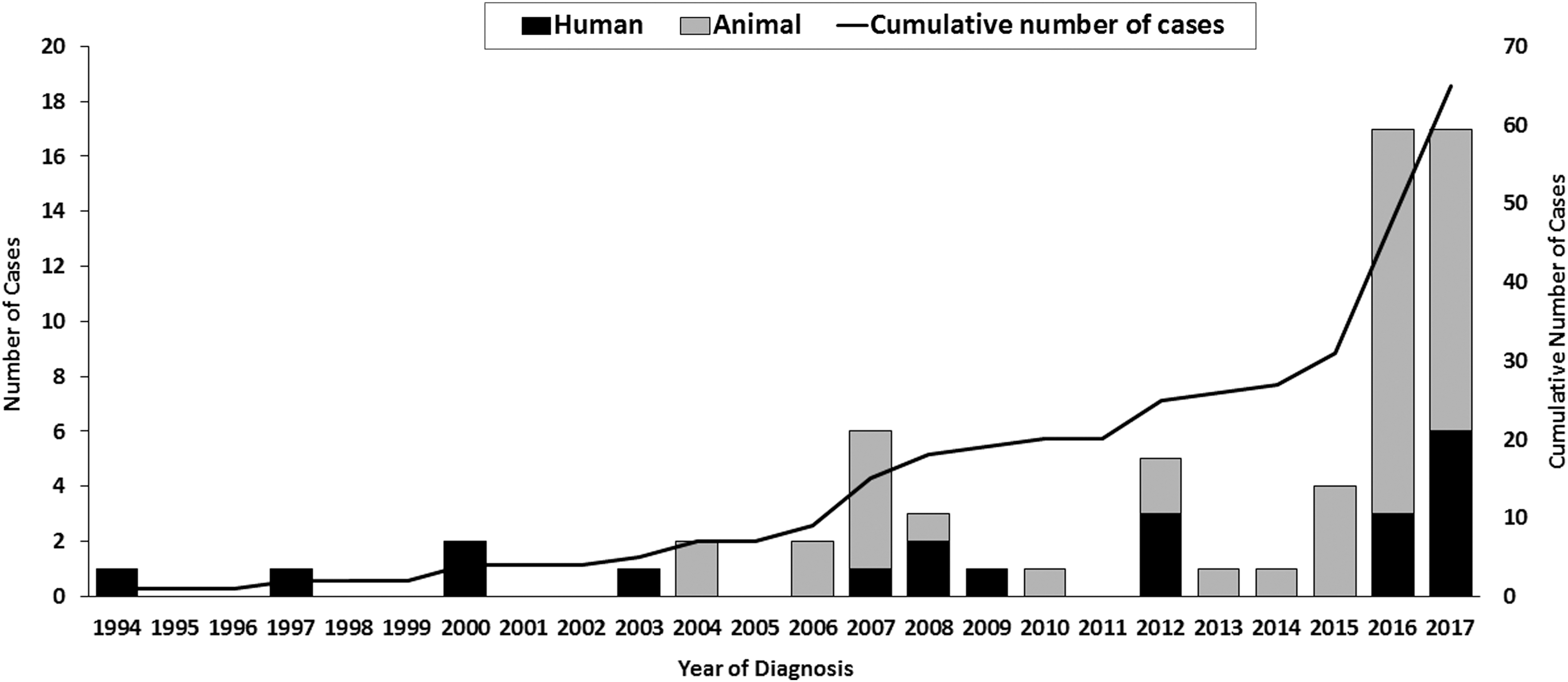
Human and animal cases of tularemia in Minnesota, 1994–2017 (n = 58).

Tularemia subtype of human tularemia cases by most likely exposure route, Minnesota, 1994–2017 (n = 21).
Since the first report of isolation from a tick in 1924, there have been multiple published studies of F. tularensis prevalence in tick species in the United States, concentrated in the 1920s through to the 1960s. The studies, using varying sampling and testing methodologies, estimated the prevalence of F. tularensis in D. variabilis ticks in the United States as 0–4.3% (Green 1931, Parker et al. 1952, Hopla 1953, Calhoun Alford, 1955, Saliba et al. 1966, Goethert et al. 2004, Goethert and Telford 2009). Published estimates of prevalence in Ixodes scapularis are less frequent and range from 0% to 2.9% (Calhoun 1955, Goethert et al. 2004, Eisen 2007). Other studies have focused on A. americanum ticks, which are common in the Southeastern United States; F. tularensis prevalence in this tick species has been reported as 0–0.5% (Hopla 1953, Calhoun 1954, Eisen 2007). Research conducted in 1930 demonstrated the presence of F. tularensis in D. variabilis ticks from Minnesota, at a prevalence <0.1% (Green 1943); this is the only known published study of F. tularensis prevalence in Minnesota tick species.
In early May 2017, the Minnesota Department of Health (MDH) received a report of an apparent epizootic of tularemia in Washington County, located on the southern edge of the Minneapolis–St. Paul metropolitan area. Two adjacent property owners reported an indoor/outdoor cat treated for suspected tularemia, ∼26 dead rabbits across both properties over the course of 1 week, and large numbers of ticks on both properties. We investigated the apparent epizootic to study the ecology of F. tularensis in Minnesota. In Minnesota, I. scapularis commonly transmit Lyme disease and other tickborne diseases; yet, this species has not been shown to be a significant vector for tularemia in the United States. In addition, A. americanum is not established in Minnesota, so D. variabilis is the most likely tick vector for human cases of F. tularensis in this area. Therefore, we focused our available resources on investigating the role of this species in the ecology of F. tularensis in Minnesota. The purpose of this study was to determine and characterize the infection prevalence of F. tularensis in D. variabilis ticks at the apparent epizootic site and compare these data with findings from long-term tick research sites in Minnesota.
Materials and Methods
Field methods
An initial site investigation was conducted using a standard site investigation form to document weather conditions, geographic location, land use, potential animal and arthropod vector species present, and any other notable characteristics at the epizootic site.
Ticks were collected from each of two contiguous properties (Washington County 1 and Washington County 2) at the epizootic site by dragging a 1 m2 white canvas cloth along the ground in wooded and grassy habitat (Fig. 3). Tick collectors wore white painter suits to protect against tick bites and to facilitate tick collection; collectors checked themselves and the drag cloth for ticks every 10–20 meters. Tick dragging was performed weekly from May 11 to July 6, 2017, corresponding to the remaining D. variabilis season in Minnesota. Specific transects were not followed each week; however, the same general areas of each property were sampled until ∼100 ticks were collected each week. Ticks were stored in vials of 70% ethanol; they were identified to species, then sorted by species, sex, collection date, site, and pooled with a maximum of 10 ticks per pool. During two collection weeks, additional ticks were collected into empty microcentrifuge tubes and kept alive until processing for bacterial culture. Of all ticks collected at the epizootic site, 2740 (99.6%) were D. variabilis and 10 (0.36%) were I. scapularis; therefore tick testing focused on D. variabilis.

Dermacentor variabilis sampling sites, Minnesota, 2017.
D. variabilis ticks were also incidentally collected during routine I. scapularis surveillance at five long-term tick research sites across central Minnesota (Anoka County 1, Anoka County 2, Morrison County 1, Clearwater County 1, and Pine County 1) in April, May, and June (Fig. 3). A convenience sample of these D. variabilis ticks was selected for testing. Weeks of specimen collection were matched with the epizootic site when possible.
Laboratory methods
All laboratory work was performed by trained laboratorians in a Select Agent registered space, following all Select Agent program requirements, including Biosafety Level 3 practices. Pooled ticks were laterally bisected and combined in a microcentrifuge tube containing 0.4 mL of 0.01 M phosphate-buffered saline. The mixture was then homogenized by either bead beating or manual grinding using a microcentrifuge tube pellet pestle. Nucleic acid extraction was performed on 0.2 mL of the supernatant from each tick pool using the Roche MagNA Pure Compact instrument with an elution volume of 0.1 mL. The extract was then tested by real-time PCR (RT-PCR) using the U.S. CDC's Laboratory Response Network (CDC LRN) F. tularensis methods. RT-PCR-positive pools were subsequently tested by the U.S. Department of Defense F. tularensis subtyping assay. This assay was used to differentiate between species, F. novicida and F. tularensis, and between the two subspecies of F. tularensis, F. tularensis type A and F. tularensis type B.
Supernatant from live-processed ticks, collected from the epizootic site specifically for culture, was tested by the above RT-PCR assays. Supernatant from positive pools was then plated on modified Thayer Martin agar for bacterial culture and isolation of F. tularensis. Suspected F. tularensis colonies were confirmed using direct fluorescence antibody (DFA) testing that detected F. tularensis cellular surface antigens (Petersen and Schriefer 2015). RT-PCR and bacterial culture were generally performed within 1 week of collection.
Analyses
Proportion of positive pools and minimum infection rate (MIR) were calculated for each site. MIR was calculated as the number of positive pools divided by the total number of ticks tested in all pools, expressed as a percentage. For categorical variables, we performed descriptive statistics including frequency, median, and range, and tabulated percentages of each value. Differences in percentages and MIR were assessed using chi-squared or Fisher's exact tests. A two-sided value of p < 0.05 was considered statistically significant.
Results
A total of 3527 adult D. variabilis ticks were collected and tested for F. tularensis comprising 1601 male D. variabilis and 1926 female D. variabilis. Across all the sites, 128 (34%) of 378 pools were positive for F. tularensis by RT-PCR (range, 0–66% of pools per site) (Table 1); the overall MIR was 3.6% (range, 0–7.2%). Of the 128 positive pools, 96 (75%) were identified as having F. tularensis type A by the RT-PCR subtyping assay; F. tularensis from the other 32 pools was unable to be subtyped using the assay.
Proportion of Adult Dermacentor variabilis Pools Real-Time PCR-Positive for Francisella tularensis by Sex and Collection Site, Minnesota, 2017
Four of the seven sites had positive pools: Washington County 1, Washington County 2, Morrison County 1, and Pine County 1. Overall, the MIR at positive sites was 4.1% (range, 2.1–7.2% by site). The highest MIR detected in this study was 10.1%, during one collection week, at Morrison County 1. The median weekly MIR at positive sites was 4.4% (range, 0–10.1% by site and collection week) (Table 2). Sex of the tick was not significantly associated with positive pools at any of the sites.
Minimum Infection Rate of Francisella tularensis in Adult Dermacentor variabilis Ticks by Sex, Collection Week, and Site, Minnesota, 2017
MIR expressed as number of infected ticks per 100 ticks tested.
The MIR was significantly higher at Morrison County 1 compared with the MIR at the epizootic sites combined (p = 0.01). Within the epizootic site, the MIR was significantly higher at site 2 compared with site 1 (p < 0.001).
MIR, minimum infection rate.
Overall, there was no difference in proportion of positive pools at the epizootic site (Washington County 1 and Washington County 2 combined) compared with all long-term monitoring sites (106 [36%] vs. 22 [26%]; p = 0.07). The overall MIR was significantly lower at the epizootic site compared with the overall MIR at Morrison County 1 (3.9% vs. 7.2%; p = 0.02), but was not significantly different between the epizootic site and the Pine County 1 (3.9% vs. 2.1%; p = 0.49). Within the epizootic site, the overall MIR was significantly higher at the second property (Washington County 2) compared with the adjacent property (Washington County 1) (5.7% vs. 2.3%; p < 0.001).
A total of 369 live-processed D. variabilis ticks, in 40 pools, were screened by RT-PCR for testing by bacterial culture. F. tularensis DNA was detected by RT-PCR in 10 (25%) pools, F. tularensis was determined to be subspecies tularensis type A in 8 (80%) pools (F. tularensis from 2 pools was unable to be subtyped). F. tularensis was isolated and confirmed by bacterial culture and DFA from 6 (60%) pools of the 10 RT-PCR-positive pools (MIR = 1.6/1000).
Discussion
F. tularensis was detected in D. variabilis ticks at a majority (57%) of sites sampled in Minnesota, including an apparent tularemia epizootic site and sites with no reported epizootic or identified cases of sporadic tularemia. One (Clearwater County 1) of the three sites where F. tularensis was not found had a robust sample size (n = 322 ticks); however, the sample sizes from Anoka County 1 (n = 18 ticks) and Anoka County 2 (n = 42 ticks) may not have been sufficient to detect F. tularensis in the collected ticks.
The highest published prevalence in D. variabilis ticks in the United States comes from Martha's Vineyard, with median annual prevalence of 3.4% (range, 2.7–4.3% annually) over the 3-year study (Goethert and Telford 2009). The overall median MIR (expressed as infected ticks per 100 ticks tested and therefore comparable with reported prevalence) of F. tularensis within the positive sites found in our study (4.1%) is similar to the prevalence found in Martha's Vineyard. Other published estimates of prevalence in D. variabilis ticks are lower, often <1% (Green 1931, Parker et al. 1952, Hopla 1953, Calhoun 1954, Saliba et al. 1966, Goethert et al. 2004). Estimates of F. tularensis prevalence in other Dermacentor species in Europe are similar to those in the United States, 1–3%, although they range up to 7%, in Dermacentor reticulatus ticks collected in the Czech and Austrian Republics (Hubalek et al. 1996, 1997, Hubalek and Rudolf 2017).
The MIR of F. tularensis in the cohort of adult D. variabilis ticks at the epizootic site in our study was 3.9% across the 9-week study; this was lower than the 7.2% MIR found at a site with no apparent epizootic of tularemia. Although a site with an active epizootic occurring may be expected to have an increased infection prevalence, we did not find this in our study. However, epizootics may occur at sites without being noticed. The epizootic (at Washington County 1 and Washington County 2) in this study occurred in a residential area where it was likely to be observed. The other long-term tick research sites consist of unpopulated, recreational areas where animals may die in larger numbers and go unnoticed by the public.
At the epizootic site, there were consistently more infected ticks collected at one of the two properties. The MIR of D. variabilis ticks also varied regionally; the MIR at Morrison County 1 was significantly higher than the overall MIR at the epizootic site. A study conducted in Martha's Vineyard also found nonrandom distribution of D. variabilis ticks infected with F. tularensis (Goethert and Telford 2009); the study found a cluster area within the sampling location that was significantly more likely to yield infected ticks and identified a microfocus where tularemia can perpetuate.
In our study, tick MIRs also displayed variation week to week, but the range in this variation was relatively small (0–10%). The reasons for this variability are not clear. The same cohort of ticks was sampled at each site; therefore, we would not expect the infection rate to change significantly at a given site during the same season. Our results may reflect normal variation in infection prevalence over time and space; however, other factors may influence infection prevalence, including weather conditions (e.g., temperature and relative humidity), host and vector population density, and tick metabolomics.
All infected ticks from which F. tularensis was successfully subtyped in this study contained F. tularensis type A, similar to ticks collected from Martha's Vineyard where all ticks were also found to have type A (Goethert et al. 2004). This is interesting because tularemia cases in Minnesota with likely tickborne exposures are caused by both F. tularensis type A and type B, and the proportions of type A (2/5) and type B (3/5) were nearly equal. It is possible that the areas sampled were not ecologically supportive of the type B cycle and therefore explains why no type B-infected ticks were found.
F. tularensis was isolated from 15% of live-processed tick pools; isolates from these pools will be further characterized to determine which F. tularensis strains are carried by D. variabilis in Minnesota. Characterizing strains carried by D. variabilis ticks and isolates from humans, animals, and other tick species will add to the unanswered research questions of whether there are host-specific strain differences, whether there is strain diversity within tularemia subtypes, and whether there are differences in virulence between and within tularemia subtypes (Goethert et al. 2004, Petersen et al. 2009).
The RT-PCR assay used in this study was developed by CDC LRN, and a limitation of our study is that data on its sensitivity and specificity for F. tularensis are not available. Therefore, it is possible that the pools that were unable to be subtyped by the assay actually contained Francisella-like endosymbionts present in the ticks. However, the subtyping assay used was developed by the Department of Defense specifically to differentiate between the subspecies of Francisella; therefore, we made the assumption that it accurately identifies F. tularensis present in the ticks. As F. tularensis from 96 (75%) of all 128 RT-PCR-positive pools was able to be subtyped, at least this proportion of RT-PCR positives likely represented true F. tularensis. To corroborate this conclusion further, F. tularensis from 8 of the 10 live-processed tick pools that were RT-PCR positive was able to be subtyped, and F. tularensis was confirmed by culture and DFA in 6 (60%) pools. An additional limitation of our study includes the absence of consistent transects sampled. Therefore, we did not account for differences in microhabitat that may affect infection prevalence. Finally, this study and previously published studies use differing methodology; therefore, comparison of reported prevalences should be interpreted with caution.
Conclusions
When able to be subtyped, all F. tularensis from ticks in this study was F. tularensis type A, and 40% of Minnesota tickborne tularemia cases had type A tularemia; this suggests that type A cycle of tularemia is important in Minnesota. In addition, because some tickborne cases in Minnesota have had type B tularemia, further study is needed to clarify the epidemiology of all tickborne tularemia cases, and the importance of other ecologic maintenance cycles in Minnesota. Long-term testing of ticks collected from known sites of tularemia activity will help to improve our understanding of natural transmission of tularemia, and the variability in infection prevalence and persistence of the bacteria over time at a given site.
Footnotes
Acknowledgments
The authors thank Matt Springer, MDH intern, for assisting with collection, speciation, sexing, and pooling of collected ticks. The authors also thank the property owners of tick sampling sites for permitting access to their properties.
Author Disclosure Statement
No conflicting financial interests exist.