
Abstract
West Nile virus (WNV) cases were seasonally recorded in humans and animals in Greece, from 2010 to 2014, and circulation of the virus was detected in different Regional Units of the country. Small scale entomological surveillance activities were carried out by several regions and regional units in Greece, during 2014–2016, with the participation of subcontractors for the vector control programs aiming to record presence/absence of mosquito species, and monitor and control mosquito populations. Mosquito traps were placed in rural and urban sites; specimens were collected, morphologically characterized, and pooled by date of collection, location, and species types. Mosquito pools containing Culex pipiens, Aedes caspius, and Aedes albopictus were examined for the presence of WNV and positive pools were detected in different areas of the country. Sequencing of a selected number of amplicons revealed WNV lineage 2 partial NS5 gene sequences. In this study, we present data on the mosquito species composition in the areas of study and WNV detection from several parts of Greece, in 6, 11, and 26 mosquito pools corresponding to the years 2014, 2015, and 2016, respectively. A total of 15 WNV human infections were reported to the public health authorities of the country in 2014, whereas no human cases were detected for 2015–2016. Taking into consideration the complex epidemiological profile of WNV and unforeseen changes in its circulation, re-emergence of WNV human cases in Greece was possible and expected, thus rendering surveillance activities imperative.
Introduction
Mosquitoes (
West Nile virus (WNV) infection is recognized as an ongoing threat in Europe, since virus circulation has been recorded both in Central Europe and the Mediterranean basin (Hubalek and Halouzka 1995, Calistri et al. 2010). West Nile fever (WNF) has been notifiable at the European Union (EU) level since 2008, with a fluctuating annual number of cases. From 2011 to 2017, the European Center for Disease Control (ECDC) has reported a total of 1,207 WNV human cases across different EU countries [European Center for Disease Control (ECDC)/Historical data by year].
WNF human cases have been recorded in Greece consecutively from 2010 to 2014, with a total number of 446 human neuroinvasive disease cases (West Nile Neuroinvasive Disease [WNND]) being recorded for that particular period [National Public Health Organization (NPHOa)].
During the summer of 2010, Greece underwent one of the largest WNV infection outbreaks in Europe, with a total of 262 clinical human cases and 35 deaths [National Public Health Organization (NPHOb)].
In the following years, 2015–2016, no human cases were detected in Greece for the first time since 2010 [European Center for Disease Control (ECDC)/Historical data by year, NPHOa].
To date, nine genetic lineages have been proposed for phylogenetical division of WNV (Lanciotti et al. 2002, Charrel et al. 2003, Pachler et al. 2014). WNV strains were considered to differ substantially in terms of virulence and neuroinvasiveness, with lineage 1 strains considered the only pathogenic forms of the virus and relatively few cases of neurological disease caused by lineage 2 strains (Venter and Swanepoel 2010). In 2004, WNV lineage 2 was first detected in Europe, isolated from the brain of a goshawk (Accipiter gentiles) in Hungary (Bakonyi et al. 2006). Subsequently, WNV lineage 2 emerged in Greece after 2010 in mosquito populations, wild resident birds, and humans (Papa et al. 2011b, Valiakos et al. 2012, Barzon et al. 2013b, Patsoula et al. 2016), and thereafter in Italy in 2011 (Bagnarelli et al. 2011), causing repeated WNF outbreaks and resulting in hundreds of human cases of WNND (Hernández-Triana et al. 2014).
Outbreaks of WNV have been associated worldwide with increased mosquito populations, usually after flooding, subsequent dry and warm weather, and the formation of suitable larval breeding habitats (Hubálek 2000). WNV has been isolated from over 40 species of mosquitoes (Diptera: Culicidae); however, the principal mosquitoes involved in WNV transmission belong to the genus Culex, particularly species in the Culex pipiens complex (Crook et al. 2002, Higgs et al. 2004). In Europe, the major vector species is Cx. Pipiens, which is widely distributed in the continent, and Culex (Barraudius) modestus Ficalbi 1889 found in wetland and fishpond areas of southern and central Europe are also suitable vectors (Bakonyi et al. 2006, Engler et al. 2013). In Greece, WNV outbreaks have been associated with Cx. pipiens, the most abundant Culex species in the country and most probably involved in the enzootic and epidemic transmission of the virus (Papa et al. 2011b, Chaskopoulou et al. 2016, Patsoula et al. 2016). So far, four European mosquito species, Aedes albopictus, Aedes detritus, Cx. Modestus, and Cx. pipiens, have been experimentally confirmed to be competent to transmit WNV, whereas Aedes caspius and Aedes japonicus japonicus are not considered to be competent as vectors for WNV (Balenghien et al. 2007, 2008, Fortuna et al. 2015, Brustolin et al. 2016, Vogels et al. 2017). The highest transmission rates in field studies were found for Cx. pipiens, rendering it the most important vector for WNV in Europe (Engler et al. 2013).
In this study, we present entomological and WNV detection data from several Regional Units (R.U.s, NUTS3 level) in Greece for 2014–2016, related to local vector surveillance activities, focusing on circulation of the virus in 2015–2016, before the re-emergence of human cases in 2017.
The goals of this study included recording of mosquito species in the studied areas, as well as emphasizing the importance of having entomological surveillance programs in place for early detection of WNV circulation in mosquito populations.
Materials and Methods
Surveyed regions, mosquito collection, and identification
In Greece, from 2010 to 2014, active mosquito surveillance programs were implemented nationwide or in certain parts of the country, on the basis of collaboration among public health authorities, and national and local bodies (Patsoula et al. 2016). For the 2014 period, an active surveillance program was implemented by the National Public Health Organization (NPHO) in collaboration with local authorities and subcontractors, while for the 2015–2016 period, small scale vector surveillance programs were conducted and funded by Regions (R, NUTS2 level), with the collaboration of subcontractors and in collaboration with the National School of Public Health (NSPH).
Two different types of traps, CO2 mosquito traps with dry ice (CDC Miniature Light Trap; BioQuip Products, Inc., Rancho Dominguez, CA) and Mosquito Traps (Skone Electric Appliance Co., Ltd.), suitable for adult mosquito collections, were used. Traps were placed in urban and rural sites of municipalities, municipal units, and local communities of the R.U.s of the Regions participating in the entomological surveillance program (Table 1), according to instructions provided by the NPHO and the Regions in 2014 and by the Regions in 2015–2016 regarding breeding sites, nuisance reported by citizens, representation of both urban and rural sites, and convenience of sampling (Fig. 1). The whole mosquito trap was collected, immediately frozen in dry ice, and subsequently sent to the laboratory of Medical Entomology of the Department of Parasitology, Entomology, and Tropical Diseases of the NSPH for morphological identification of the mosquitoes and WNV molecular detection. Identification of mosquitoes, based on morphological characters, was performed after careful examination under an SMZ-U Nikon binocular scope, using appropriate dichotomous keys (Darsie and Samanidou-Voyadjoglou 1997, Samanidou-Voyadjoglou and Harbach 2001, Becker et al. 2010, Voyadjoglou-Samanidou 2011, Wilkerson et al. 2015).

Geographical distribution of mosquito traps with a schematic representation of Regional Units participating in the entomological activities. The inset map represents in detail the Attica and Islands Region: 2014: n = 11;  2015: n = 8;
2015: n = 8;  2016: n = 5. Color images are available online.
2016: n = 5. Color images are available online.
Locations of Mosquito Traps by Regions and Regional Units Participating in Entomological Surveillance Programs, Greece (2014–2016)
Regarding morphological identification of members of the Cx. pipiens complex (Darsie and Samanidou-Voyadjoglou 1997, Becker et al. 2010), no male Culex torrentium adults were identified in this study. Regarding identification of adult females, they were characterized morphologically as Cx. pipiens as species are indistinguishable morphologically, and currently, we are not performing molecular studies for further identification.
Adults morphologically identified to belong to the Anopheles maculipennis complex were further examined by molecular amplification methods, following procedures and protocols previously described (Patsoula et al. 2007, Beleri et al. 2017).
Detection of WNV in mosquito pools
All collected mosquitoes were sent to the Department of Parasitology, Entomology and Tropical Diseases (DPETD) in NSPH in closed and chilled containers containing dry ice. They were pooled according to date, location, and species, with a minimum of two and a maximum number of 200 individuals per pool. The pooled mosquitoes were stored and frozen at −80°C.
RNA from mosquito pools was extracted using the Maxwell 16 Automated Nucleic Acid extraction system (Promega, Madison, WI), along with the Maxwell 16 LEV Simple RNA Tissue kit, as previously described (Patsoula et al. 2016). For WNV detection, a TaqMan Real-Time PCR protocol specific for WNV lineages 1 and 2 was utilized (Tang et al. 2006). Positive control, an RNA sample of WNV, and negative controls, RNA samples from several arthropod-borne viruses such as Dengue and Chikungunya viruses, were also included in the reaction and were kindly donated by Dr. Paolo Bonilauri (IZSLER, Reggio Emilia, Italy). A series of representative positive samples from all years were subjected to a flavivirus genus-specific seminested protocol (Scaramozzino et al. 2001), PCR products were excised from a 2% agarose gel, and DNA was isolated using the Invisorb Spin DNA Extraction Kit (Stratec Molecular GmbH), according to manufacturer's instructions. Sequencing was performed by order, using primers employed in the inner nested PCR (Scaramozzino et al. 2001). Obtained sequences were employed to perform basic local alignment search tool (BLAST) in the GenBank library to confirm the specificity of positive reactions and to estimate the degree of identity of previously detected strains. Subsequently, they were aligned with available deposited GenBank sequences by ClustalOmega available at
Results
A total of 51,704 (50,901 ♀, 803 ♂) adult mosquitoes were captured in the surveyed Regions (n = 12), during the 3-year entomological surveillance activities, and 20 species were identified belonging to six genera. In more details, in 2014, a total of 45,547 (88.1%) individuals were trapped representing 16 species and belonging to five genera, while in 2015, a total of 2,182 (4.2%) individuals were trapped, representing 14 species belonging to 6 genera, and in 2016, a total of 3,975 (7.7%) individuals were trapped, representing 18 species and belonging to 6 genera (Table 2 and Fig. 2). Species composition and relative abundance of mosquitoes trapped per region and per sampling year during the entomological surveillance programs (2014–2016) are presented in Table 3 and Fig. 2.
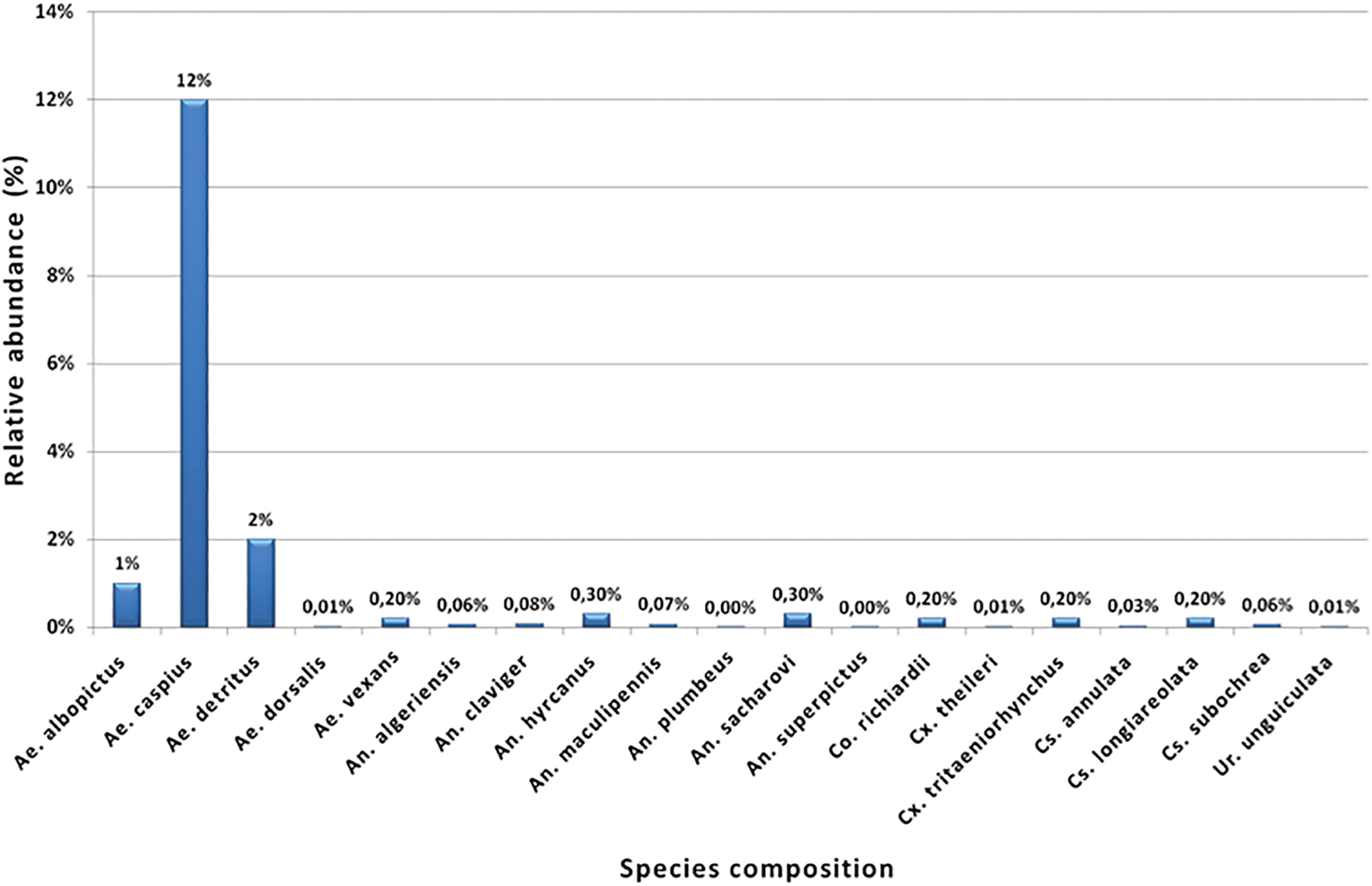
Morphological identification and relative abundance (%) of mosquito species collected during the entomological surveillance programs (2014–2016). Culex pipiens (n = 43,500, 83%) were by far the most abundant species (not included in this Figure), followed by Aedes caspius (n = 6,131, 12%). Color images are available online.
Number of Mosquitoes Tested for West Nile Virus (Nt) and Number of West Nile Virus-Positive Mosquito Pools (Np) per Mosquito Species, Region, and Year, Greece, 2014–2016
Np, number of pools positive for WNV; Nt, number of tested pools; WNV, West Nile virus.
Species Composition and Relative Abundance in Adult Mosquitoes Trapped per Region and per Sampling Year
Aedini denomination according to Wilkerson et al. (2015).
Adult morphologically characterized as members of the An. maculipennis complex was further examined by molecular amplification methods (Patsoula et al. 2007, Beleri et al. 2017), and according to their RFLP profiles, they were verified as An. maculipennis and Anopheles sacharovi, as presented in Table 3.
Out of the 12 Regions that participated in this survey, entomological data were systematically gathered from Attica, Crete, East Macedonia and Thrace, and North Aegean. The composition of mosquito assemblages differed by study area; however, the most abundant species was Cx. pipiens, representing 84% of all collected individuals, followed by Ae. caspius, 12%. A diverse and very abundant mosquito fauna was recorded in the region of East Macedonia and Thrace, as noted in previous studies (Beleri et al. 2017).
A total of 49,262 female mosquitoes, captured during the 3-year activities, belonging to Cx. pipiens (n = 42,953), Ae. caspius (n = 6,098), and Ae. albopictus (n = 211) were pooled and tested for WNV. Out of 1,122 mosquitoes tested, a total of 43 mosquito pools were tested positive for WNV, including 37/43 positive samples (86%) among Cx. pipiens mosquitoes, 1/43 (2.35%) positive samples among Ae. albopictus, and 5/43 (11.65%) positive samples among Ae. caspius (Table 3 and Figs. 3 and 4).
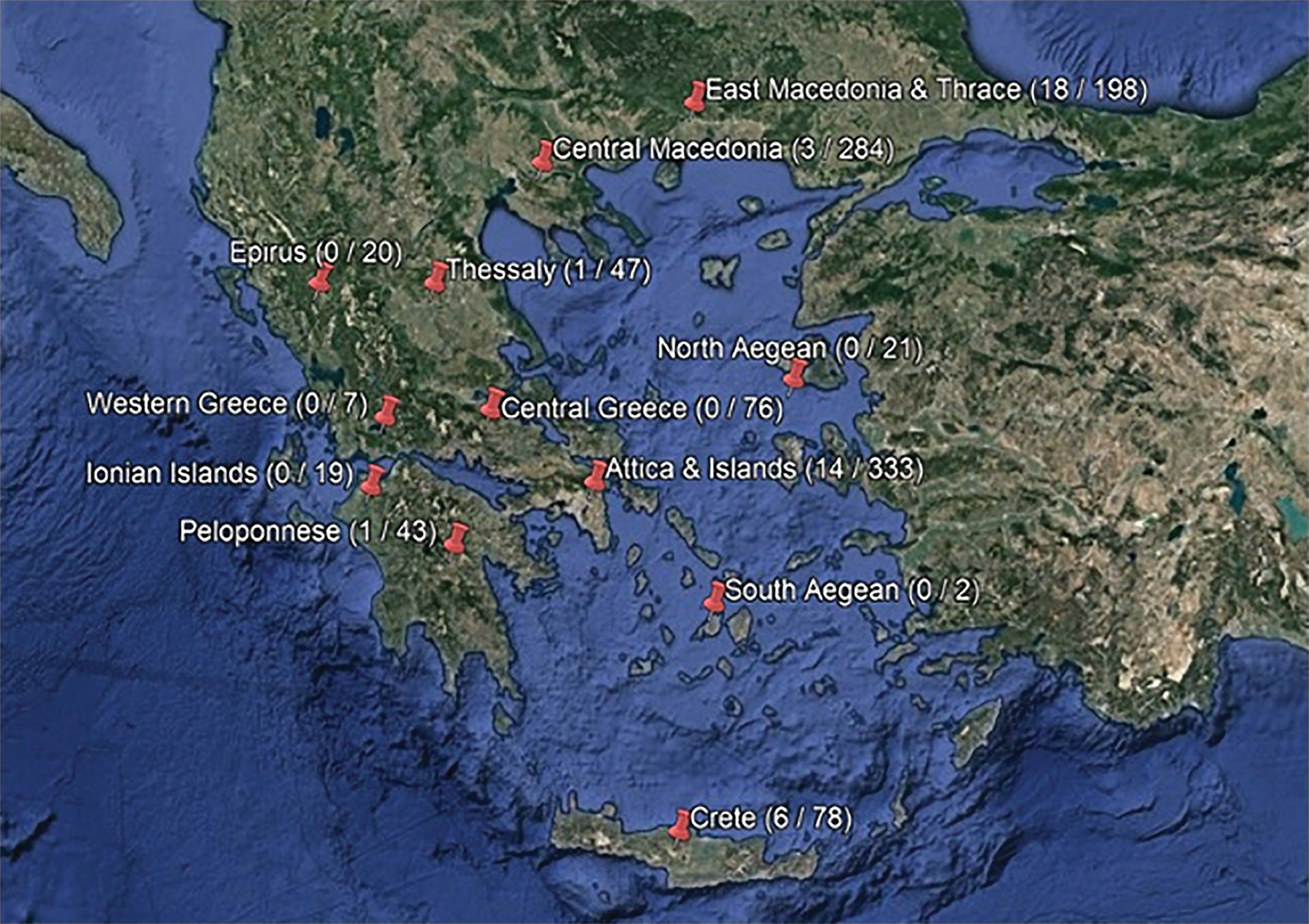
Regions with positive mosquito pools and total number of WNV-positive pools per region, 2014–2016. 2014: Culex pipiens (n = 6): Attica & Islands (1); Central Macedonia (3); Crete (1); Thessaly (1); 2015: Cx. pipiens (n = 9): Attica & Islands (7); Crete (1); Peloponnese (1); Aedes caspius (n = 1): East Macedonia and Thrace; Aedes albopictus (n = 1): East Macedonia and Thrace; 2016: Cx. pipiens (n = 22): Attica & Islands (6); Crete (4); East Macedonia and Thrace (12); Ae. caspius (n = 4): East Macedonia and Thrace. WNV, West Nile virus. Color images are available online.
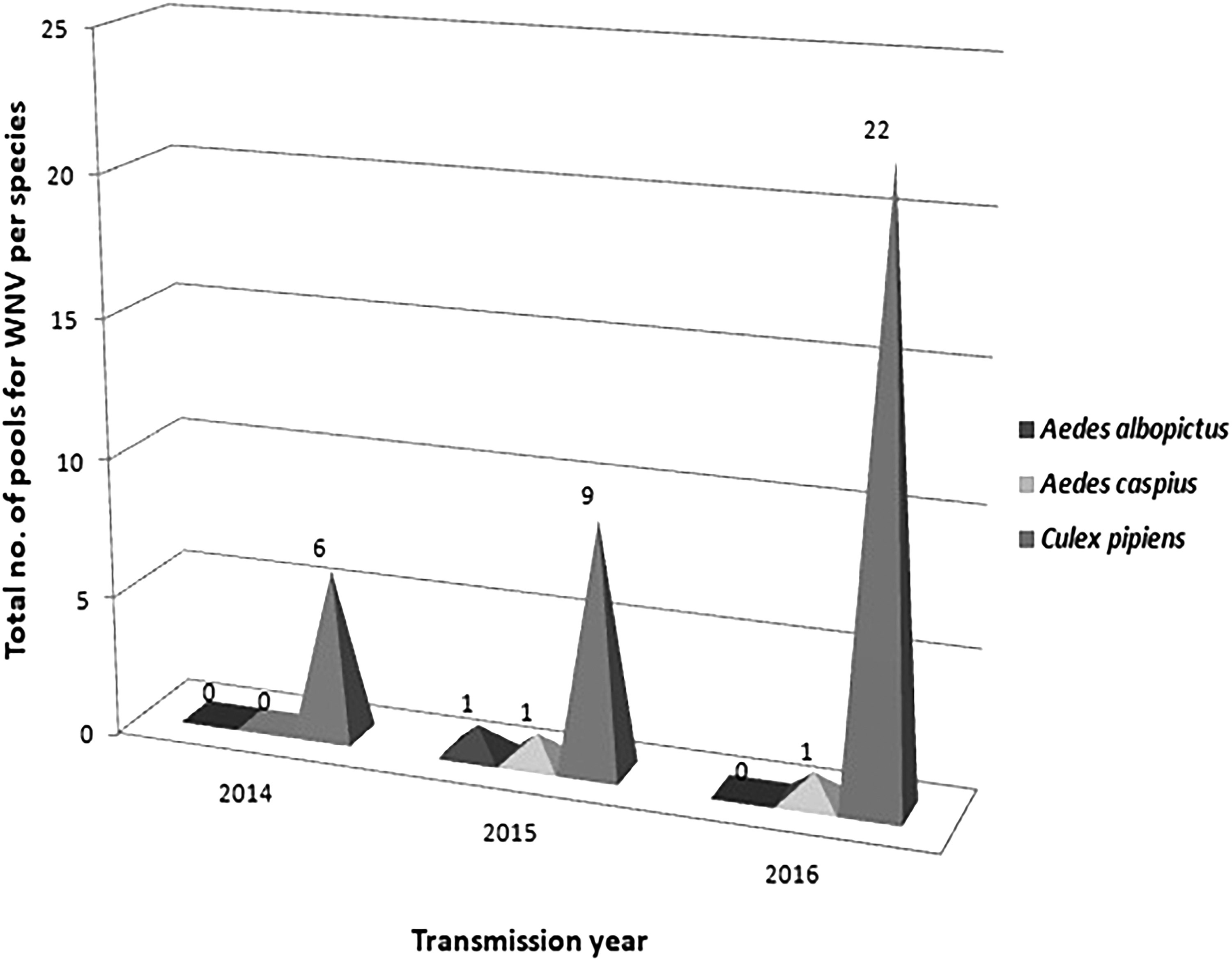
Total number of WNV-positive pools per species per transmission year.
For 2014, 514 Cx. pipiens, 40 Ae. albopictus, and 139 Ae. caspius pools were examined for the presence of WNV. For the following year, 113 Cx. pipiens, 22 Ae. albopictus, and 22 Ae. caspius, while for 2016, 208 Cx. pipiens, 17 Ae. albopictus, and 47 Ae. caspius pools, respectively, were checked for the virus presence (Table 2).
PCR products of the flavivirus genus-specific protocol (Scaramozzino et al. 2001), for 10 positive WNV samples from Cx. pipiens pools (3 for 2014 from Attica & Islands, Thessaly, and Central Macedonia, 3 for 2015 from Crete and Attica & Islands, and 4 for 2016 from Attica & Islands, Crete and East Macedonia and Thrace), were sequenced and the respective sequences were aligned with WNV lineage 2 strains deposited in GenBank by ClustalOmega, to estimate the degree of identity of previously detected strains. The obtained sequences presented similarities that ranged from 98.8% to 100% with the HQ537483 GenBank sequence (nucleotides 9,115–9,282), WNV lineage 2 isolate Nea Santa-Greece-2010 (Papa et al. 2011a, 2011b), and with previously deposited sequences of WNV partial gene for polyprotein, region of nonstructural protein 5 (Patsoula et al. 2016), from our previous studies in Greece. These 10 sequences from different Regional Units and different years were deposited in GenBank with accession numbers MK795947 to MK795956. Percentages of nucleotide identities among representative WNV lineage 2 genomes of the European clade with sequences reported from our study are presented in Table 4.
Percentages of Nucleotide Identities Among Representative West Nile Virus Lineage 2 Sequences of the European Clade with Sequences Reported from this Study
Discussion and Conclusion
Greece is one of the most WNV-affected European countries with outbreaks of the virus being recorded since 2010. As such, a well-established and sustainable vector surveillance network at national level, for the monitoring of mosquito vectors and pathogen detection, should be implemented.
The conductance of such studies, on a small scale, in 12 different Greek regions, revealed 20 out of a total of 60 mosquito species that have so far been recorded in Greece (Darsie and Samanidou-Voyadjoglou 1997, Samanidou-Voyadjoglou and Harbach 2001, Voyadjoglou-Samanidou 2011). The extracted data were particularly important for several regions (Crete, Epirus, Ionian islands, North Aegean, South Aegean, and Western Greece), where entomological data were scarce or incomplete. Three (3) different species were recorded in the majority of the selected regions, which have been studied in foreign literature for their medical and veterinary importance (Cx. pipiens, Ae. albopictus, and An. sacharovi) (Schaffner et al. 2001, Becker et al. 2010, Voyadjoglou-Samanidou 2011, WHO 2014).
Cx. pipiens was the most abundant species in this study, present in all regions, and is considered a main vector of WNV in Europe (Hubalek and Halouzka 1995, Gabriel et al. 2008). Similar findings were reported in a recent Greek study (Mavridis et al. 2018), implicating Cx. pipiens as the most likely vector associated with disease occurrence in affected and vulnerable areas in Greece for the year 2017. Endemic outbreaks of WNF, which have been reported in Italy, France, Romania, Hungary, Russia, and Spain, have also been attributed to that particular species (Chaskopoulou et al. 2016). Ae. albopictus (Asian tiger mosquito) has already been recorded in Greece from previous studies (Patsoula et al. 2006, Beleri et al. 2017, LIFE CONOPS, Badieritakis et al. 2018) and is a vector of many pathogens (Semenza and Suk 2018). In this study, adults of Aedes (Stegomyia) cretinus Edwards 1921 (Darsie and Samanidou-Voyadjoglou 1997) were not identified. This species is indigenous in Greece and shares many common morphological features and bioecological similarities with the invasive species Ae. albopictus (Giatropoulos et al. 2012, 2015). Ae. cretinus has a limited distribution across the world, Cyprus, Greece, Georgia, and Turkey (Becker et al. 2010), while in Greece, it has been reported from Crete, Macedonia, and some suburbs of northeastern Athens (Samanidou-Voyadjoglou and Koliopoulos 1998). In recent years, there has been observed the limited presence of Ae. cretinus following the invasion and spread of Ae. albopictus in Greece (Giatropoulos et al. 2012). The study of Giatropoulos et al. (2015) revealed that Ae. albopictus was superior competitor concerning the interspecific larval competition and the interspecific crosses of adults, and that is a possible explanation for Ae. cretinus displacement.
Mosquito populations of different Anopheles species (An. maculipennis, An. sacharovi, and Anopheles superpictus), important in terms of public health, have also been recorded in Greece from previous studies (Beleri et al. 2017, MALWEST). Species such as Anopheles hyrcanus and Anopheles claviger, which are considered elsewhere as potential malaria vectors, should be further studied (Rueda et al. 2006).
In 2014, 15 cases of WNV infection were recorded in humans in Greece (NPHOa), in the R.U.s of Ilia in Western Greece, East Attica in Attica Region, and Rodopi and Xanthi in the East Macedonia and Thrace Region. Regarding the presence of the virus in mosquito populations, only 6 positive Cx. pipiens pools were detected in a total test population of 693 pools, in the Regions of Central Macedonia, Crete, Thessaly, and Attica. The fact that the geographical areas with recorded human WNV cases did not participate or partially participated at the 2014 entomological surveillance program limits the validity of conclusions.
The following years, 2015–2016, no human cases were reported to the NPHO (NPHOa); however, based on our findings, the virus was still circulating in mosquitoes as it was detected in 11 and 26 mosquito pools, respectively, in 3 Regions in 2015 and 3 Regions in 2016, in Greece. Possible explanation for the absence of human cases in 2015–2016 included different seasonal patterns of WNV circulation and activity, in terms of virulence, bird population size and immunity, vector abundance, local weather conditions, human behavior, or the underdiagnosis of human WNV disease in the country. WNV has been circulating in the country and the WNV epidemic in humans has been present since 2010; therefore, as a consequence, part of the population has been somehow exposed to the virus and acquired immunity. The decrease of WNV-reported human cases in the years 2013–2014 (86 and 15, respectively) is noteworthy, the majority being WNND with high mortality rates, indicating that, mainly severe cases have been diagnosed and recorded by the national public health authorities (NPHOa).
A discrepancy between detection of WNV lineage 2 in vectors and absence of human WNV cases of the same type of infection has also been noticed in Italy in 2011 and 2012 (Barzon et al. 2013a). In the Emilia—Romagna region, human cases were reported in 2009 and 2013 (Angelini et al. 2010, Calzolari et al. 2012, Bellini et al. 2014), while no cases were reported between 2010 and 2012. WNV-positive mosquito pools were collected only in 2009, 2010, and 2013 (Bellini et al. 2014). Furthermore, in Vojvodina, Serbia, positive mosquito pools have been detected since 2010, and the first human cases were diagnosed only in 2012 (Petrić et al. 2017). Also, in the 2014 season, WNV activity was detected in four out of seven districts surveyed at a very low level, while no human cases were registered (Petrić et al. 2017). According to a recent study (Marini et al. 2018), WNV is more likely to be reactivated at the beginning of each vector breeding season by diapausing mosquitoes previously infected.
Regarding analysis of positive mosquito pools by species, the presence of WNV RNA was demonstrated mainly in Cx. pipiens (86%) (Gabriel et al. 2008, Patsoula et al. 2016) and occasionally in the other species tested, Ae. albopictus (2.35%) and Ae. caspius (11,65%) (Barzon et al. 2013a, Patsoula et al. 2016). WNV-positive pools were detected in the areas with known WNV circulation during the previous years (Patsoula et al. 2016), including the Regions of Attica & Islands, Central Macedonia, East Macedonia and Thrace, and Thessaly, as well as in additional areas, such as the Region of Crete. WNV positivity in mosquitoes was not documented in the Regions of Epirus, Ionian Islands, North Aegean, Western Greece, Central Greece, and South Aegean, although for most areas, the number of pools tested was low due to the lack of systematic entomological surveillance activities or difficulties in participation.
In this study, sequencing revealed WNV partial gene sequences for NSP5 region, identical to WNV lineage 2 strain 2010 Nea-Santa, with no significant sequence diversity among different locations and collection periods, indicating that the lineage is well established in Greece.
From the available vector surveillance data from the years 2011–2012, it was demonstrated that in municipalities with both positive mosquito pools and human cases recorded, the detection of positive mosquito pools preceded human cases with a median number of days of 29 (3–42) and 43 (5–93), for 2011 and 2012, respectively (Patsoula et al. 2016). Therefore, entomological surveillance data detected virus circulation before the recording of human cases in some areas, and in several cases, entomological surveillance data on vector density and rate of infected mosquitoes could provide an early prediction of the occurrence of human cases (Gobbi et al. 2014).
This study highlights that knowledge about mosquito fauna in Greece is mostly related to human disease outbreaks, rather than the result of systematic entomological surveillance. WNV is likely to cause sporadic cases and large and small unforeseeable clusters for decades to come and is considered unpredictable and difficult to control (Petersen and Fischer 2012). Therefore, in the context of the One Health approach, a multitask surveillance network is recommended, including humans, animals (horses and birds), and mosquitoes. The implementation of the One Health approach for WNV surveillance in Europe is progressively highlighted in the literature (Gossner et al. 2017).
Nationwide systematic vector surveillance programs should be established to monitor mosquito species and vector distribution in place and time, and to predict WNV circulation at the local level, to assess the risk and inform local authorities.
Footnotes
Acknowledgments
The authors would like to thank all Regions and Regional Units, local authorities, and the subcontractor companies for mosquito management for participating in entomological surveillance activities from 2014 to 2016, including adult mosquito collection, and sending samples to the Laboratory of Medical Entomology of NSPH. Finally, we thank As. Professor Olga Kavoura (NSPH) for performing an English language review of the article.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
The laboratory work was partly funded by the National Public Health Organization (NPHO) under the framework of the strategic collaboration program NPHO-NSPH.