
Abstract
We have reported high seroprevalence to Thailand orthohantavirus (THAIV) or THAIV-related orthohantavirus (TRHV) among patients with chronic kidney disease of unknown etiology in Girandurukotte, Sri Lanka. THAIV or TRHV infection is considered to be transmitted by rodent hosts in this area, but its reservoir rodents have not yet been identified. Hence, 116 rodents were captured, and seroprevalences were examined by indirect immunofluorescent antibody assay (immunofluorescence assay [IFA]) using antigens of THAIV strain Thai749-infected Vero E6 cells and recombinant nucleocapsid protein of THAIV expressed in Vero E6 cell. Molecular biological species identification of rodents was carried out by sequencing rag1, irbp, and mitochondrial cytb genes. The majority (112/116) of the captured rodents were lineage Ib of black rats (Rattus rattus). Among them, 19.6% (22/112) of the rats possessed antibodies against THAIV. Also, a lesser bandicoot rat (Bandicota bengalensis), which belongs to the Sri Lankan endemic genetic lineage, was seropositive (1/1). Two Mus booduga and one Murinae sp. were seronegative. Rodent sera showed less cross-reactivities to antigens of Vero E6 cells infected with Hantaan orthohantavirus (HTNV), Seoul orthohantavirus (SEOV), and Puumala orthohantavirus (PUUV) in IFA. These results suggest that the hantavirus present in rodents in Sri Lanka is related to THAIV or TRHV rather than to SEOV, HTNV, or PUUV. However, it might be serologically distinct from the prototype THAIV strain, Thai749, used in this study. This study revealed that black rats and lesser bandicoot rats belonging to Sri Lankan endemic lineages are possible reservoirs for THAIV or TRHV in Girandurukotte. Further multiple geographical studies are needed to confirm the THAIV or TRHV reservoir status of black and lesser bandicoot rats in Sri Lanka.
Introduction
Hantaviruses are rodent-borne viruses that belong to the genus Orthohantavirus in the family Hantaviridae. The virus genome comprises three single-stranded negative-sense RNA genome segments, called large (L), medium (M), and small (S), encoding viral polymerase, two envelope glycoproteins (Gn and Gc), and the nucleocapsid protein (NP) (Elliot 1990).
There are two clinical manifestations caused by hantaviruses in humans: hemorrhagic fever with renal syndrome (HFRS), which is typically distributed in Eurasia, and hantavirus pulmonary syndrome in the Americas (Jonsson et al. 2010). About 90% of the HFRS cases have been reported in East Asia, and epidemiological information about hantaviruses in South Asia is limited (Kariwa et al. 2007). The pathogenicity of THAIV is still controversial due to a lack of information. Up to date no confirmed clinical HFRS cases have been reported from Sri Lanka according to the literature.
Each hantavirus has a single predominant rodent species that serves as its natural reservoir, and it is thought to coevolve with its host rodent (Plyusnin et al. 1996, Schmaljohn and Hjelle 1997). HFRS in Asia is caused by Hantaan orthohantavirus (HTNV) carried by striped field mice (Apodemus agrarius), Seoul orthohantavirus (SEOV) carried by brown rats (Rattus norvegicus), and Puumala orthohantavirus (PUUV) carried by voles (Myodes sp.) (Schmaljohn and Hjelle 1997). Epidemiological studies have shown that hantavirus infections spread through rodents in Southeast Asia (Reynes et al. 2003, Truong et al. 2009, Luan et al. 2012, Ibrahim et al. 2013). Thailand orthohantavirus (THAIV) is the only hantavirus species carried by Bandicota indica in Thailand (Elwell et al. 1985). Several studies have shown seroprevalence of THAIV in human in Thailand, India, and Sri Lanka (Pattamadilok et al. 2006, Chandy et al. 2009, Gamage et al. 2011). These findings indicate that THAIV and THAIV-related orthohantavirus (TRHV) are circulating throughout South and Southeast Asia.
Chronic kidney disease (CKD) is a worldwide health concern, and its most common etiologies are diabetes mellitus, hypertension, and glomerulonephritis. A high prevalence of CKD of unknown etiology (CKDu) in Sri Lanka over the past two decades has been reported (Chandrajith et al. 2011). CKDu usually affects rural agricultural communities (Athuraliya et al. 2011), and males are more often affected than females (Jayasekara et al. 2015). However, the etiology of CKDu has not been determined yet.
Recently, we have reported that 72 (54.5%) of 132 CKDu patients in Girandurukotte District Hospital in Uva Province of Sri Lanka possessed serum antibodies to hantavirus (Gamage et al. 2017). Simultaneously, 89 antihantavirus antibody positive human sera obtained from Girandurukotte were serotyped, and the existence of THAIV or TRHV infection in that area was confirmed (Yoshimatsu et al. 2019). These results suggest that transmission of THAIV or TRHV from reservoir rodents to residents occurs in that area. Furthermore, a recent study on CKDu patients in Polonnaruwa General Hospital in North Central Province of Sri Lanka showed elevated immunoglobulin G (IgG) antibodies for hantavirus (Wijkstrom et al. 2018). Both Girandurukotte and Polonnaruwa are known as highly endemic areas for CKDu in Sri Lanka (Chandrajith et al. 2011). These results supported the hypothesis that hantavirus could be a risk factor for the development of CKDu in Sri Lanka.
There has been no report on hantaviruses and their reservoir rodents in a CKDu endemic area of Sri Lanka. In this study, rodents were captured in Girandurukotte, and their sera were screened for antihantavirus antibodies for THAIV by an indirect immunofluorescent antibody assay (immunofluorescence assay [IFA]). The usefulness of the recombinant NP (rNP)-based IFA for serological screening of rodent sera was evaluated by comparison with the reactivity to the hantavirus-infected cell-based IFA. The usefulness of the recombinant NP (rNP)-based IFA for serological screening of rodent sera was evaluated by comparison with the reactivity to the hantavirus-infected cell-based IFA, designated as authentic antigen.
In addition, molecular biological species identification was done to reveal precise species and lineage information of captured rodents from Girandurukotte, Sri Lanka.
Materials and methods
Cells
The virus was propagated in Vero E6 cells (ATCC C1008). Vero E6 cells were cultured in a T-75 cm2 tissue culture flask with a vented cap in Eagle's minimum essential medium (MEM; Gibco, Life Technologies Corporation, NY) supplemented with 5% heat-inactivated fetal bovine serum (Biowest, Nuaillé), MEM nonessential amino acids (Gibco), insulin–transferrin–selenium (Gibco), penicillin (50 U/mL) and streptomycin (50 μg/mL) (Sigma-Aldrich Co., St Louis, MO), and Gentamicin (100 μg/mL) (Sigma) in a 5% CO2 incubator at 37°C.
Viruses
Prototype THAIV strain Thai749 was isolated from a great bandicoot rat in Thailand (Elwell et al. 1985). SEOV strain SR-11, PUUV strain CG1820, and HTNV strain 76–118 were used as representative viruses (Lee et al. 1978, Kitamura et al. 1983, Gott et al. 1997). These viruses were propagated in Vero E6 cells, and the culture supernatant was collected. The virus stock was dispensed into vials and stored at −80°C until use.
Sampling
Wild rodents were captured in December 2016 (n = 13) and June 2017 (n = 103) for a survey of hantavirus infection at various locations in Girandurukotte, Sri Lanka (Fig. 1). This survey was carried out by using protocols approved by the Ethics committee of the Faculty of Veterinary Medicine and Animal Sciences of University of Peradeniya, Sri Lanka (VER-16-007). Rodents were trapped and collected from the houses of CKDu patients and neighbors. Sex, body weight and lengths of the head and body, tail, ear, skull and hind foot pad of the rodents were recorded for subsequent identification of rodent species. Blood samples were collected by cardiac puncture under anesthesia, and sera were separated. Before processing, kidney tissues were collected and preserved in 99% ethanol at 4°C.
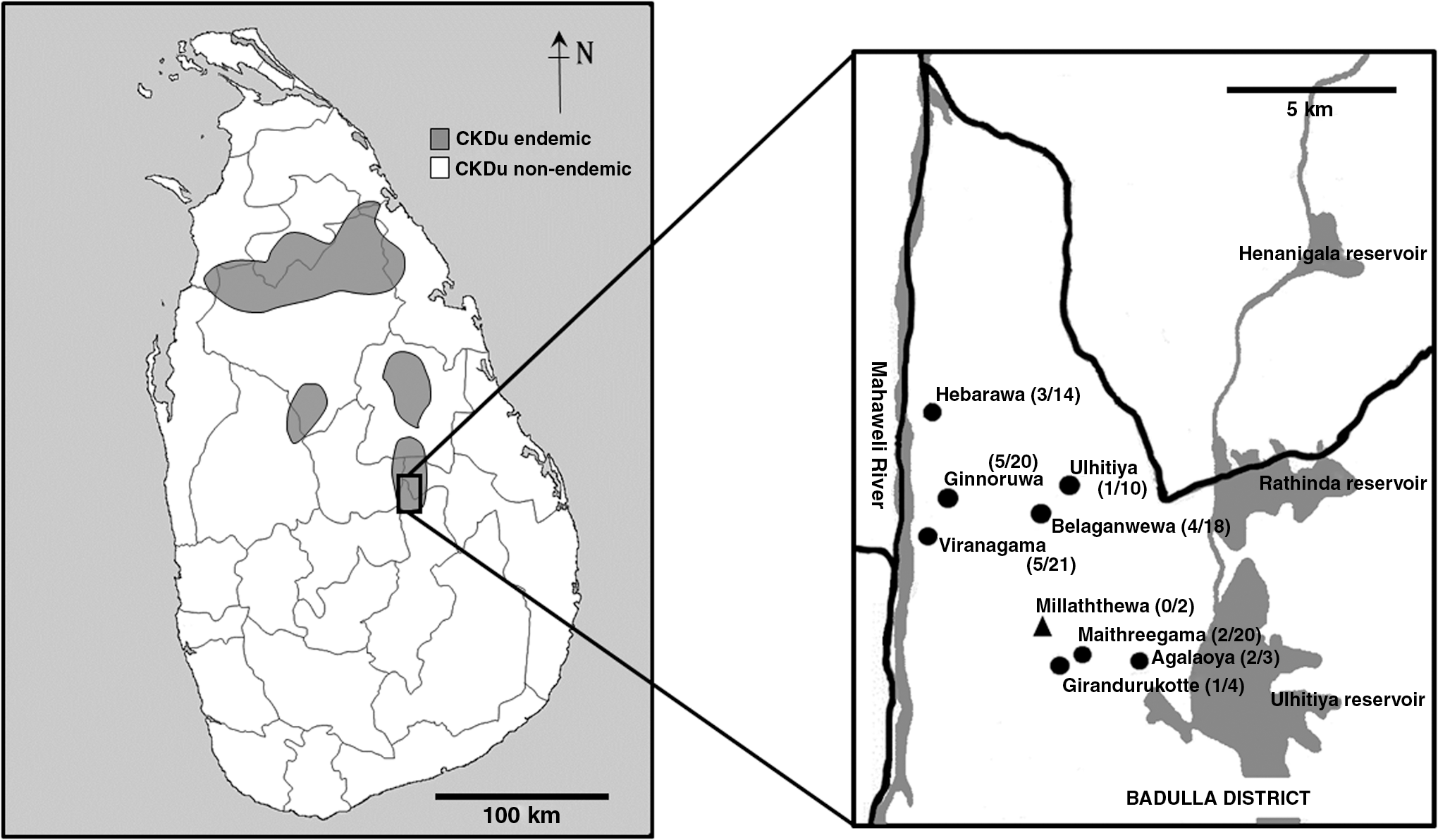
Map of Sri Lanka showing the locations of sampling sites (gray—chronic kidney disease of unknown etiology endemic areas, white—nonendemic areas). Sera were collected from Girandurukotte Town and eight surrounding villages that belong to Badulla District (Hebarawa, Ginnoruwa, Ulhitiya, Belaganwewa, Viranagama, Millaththewa, Maithreegama, and Agalaoya). Each location (number of seropositive rodents/numbers of captured rodents) is shown. Circles show the sites inhabited by seropositive rodents and triangles show the sites inhabited by seronegative rodents.
DNA isolation, amplification, and sequencing
DNA was extracted from rodent kidneys using DNAZOL Reagent (Invitrogen, Thermo Fisher Scientific, Carlsbad, CA) according to the manufacturer's protocol. A DNA fragment containing the coding region of the mitochondrial cytochrome b gene (cytb) was amplified by using primer pairs H15300 (5′-GTTTACAAGACCAGAGTAAT-3′) (Yasuda et al. 2005), L497A (5′-CCTAGTAGAATGAATCTGAGG-3′), and H655A (5′-TGTGTAGTATGGGTGGAATGG-3′) (Yasuda et al. 2014). Coding regions of nuclear gene, recombination-activating gene 1 (rag1) were also amplified by using primers +IRBP217 (5′-ATGGCCAAGGTCCTCTTGGATAACTACTGCTT-3′), −IRBP1531 (5′-CGCAGGTCCATGATGAGGTGCTCCGTGTCCTG-3′) (Stanhope et al. 1992), IRBP-335F (5′-CATCTCAGACCCTCAGACGCT-3′), IRBP-976R (5′-CACCTAGGGGTCCCAGGGACCT-3′), IRBP-932F (5′-GCCCTGGACCTCCAGAAGCT-3′), and IRBP-1530R (5′-TGATGAGGTGCTCCGTGTCCT-3′). Furthermore, the nuclear gene encoding interphotoreceptor retinoid-binding protein (irbp) was amplified by using primers RAG1-F1851 (5′-ACATGGAAGAAGACATCTTGGAAGG-3′) (Sato et al. 2004) and RAG1-R2864 (5′-GAGCCATCCCTCTCAATAATTTCAGG-3′) (Teeling et al. 2000). The nucleotide sequences of amplified cytb, rag1, and irbp were determined using the BigDye Terminator v3.1 cycle sequencing kit (Applied Biosystems, Life Technologies, Warrington) and Genetic Analyzer 3130xl.
Preparation of authentic antigen for IFA
Vero E6 cells were inoculated with THAIV, SEOV, HTNV, and PUUV at a multiplicity of infection of 0.01. After 5 days of incubation at 37°C in a 5% CO2 incubator, cells were harvested and cultivated on 24-well glass slides (Matsunami Glass, Osaka) overnight in a CO2 incubator. Cell sheets on the wells were washed with phosphate-buffered saline, and then fixed with absolute acetone at room temperature for 10 min and rinsed with distilled water. After drying, the slides were stored at −80°C until further use (Yoshimatsu et al. 1993).
Preparation of rNP-based antigen for IFA
Total RNA was obtained from the culture supernatant of Vero E6 cells inoculated with THAIV by using a QiaAmp Viral RNA Mini kit (QIAGEN, Hilden). cDNA was synthesized from total RNA by using Super Script II reverse transcriptase (Thermo). The coding region of NP of THAIV was amplified from cDNA by PCR using the following primer pair: THAIVS-46F-ClaI (5′-CCTATCGATATGGCAACTATGG-3′) and THAIVS-1335R-XhoI (5′-ACACTCGAGTTAGAGTTTTAATGG-3′). The amplified NP coding region tagged with ClaI and XhoI recognition sites was cloned into a mammalian expression plasmid vector, pCAGGS/MCS (Niwa et al. 1991). Vero E6 cells were transfected with pCAGGS/MCS containing the coding information of THAIV NP using TransIT-LT1 transfection reagent (Mirus Bio, Madison) according to the manufacturer's instructions. Preparation of IFA slides containing the rNP-based antigen was done as previously described.
Immunofluorescence assay
The IFA was performed by the same method as described previously (Yoshimatsu et al. 1993) with Alexa Fluor 488®-conjugated goat antirat (1:250 dilution) and antimouse (1:250 dilution) IgG antibodies (Invitrogen) as secondary antibodies for rat sera (R. rattus and B. bengalensis) and mouse sera (Mus booduga, Murinae sp., and Mus musculus), respectively. Screening tests at the highest serum dilution of 1:100 were performed for all rodent sera. Fluorescent images were captured on Eclipse E600 microscope equipped with a digital camera (Nikon, Kawasaki). Immune laboratory rat sera experimentally inoculated with THAIV were used as positive controls (Yasuda et al. 2012). Laboratory rat sera without inoculation were used as negative controls. Rodent serum showing a clear immunofluorescence pattern in the cell cytoplasm was considered as positive by the IFA, and a rodent serum specimen not showing a specific fluorescent pattern was considered as negative. Judgment of the IFA tests was carried out double blinded by two to three examiners.
IFA antibody titers of rat sera were determined by using twofold serial dilutions starting from 1:100 on IFA slides of authentic antigens of THAIV, SEOV, HTNV, and PUUV. The IFA antibody titers were expressed as the reciprocals of serum dilution that caused clear immunofluorescence in the cell cytoplasm.
Next, the monoclonal antibody E5G6 labeled with Alexa Fluor 594® (Invitrogen) fluorescent dye was prepared as previously described (Yoshimatsu et al. 1996, Ogino et al. 2004), and a direct IFA was carried out for comparison of image intensity between rNP-based and authentic antigens of THAIV. A total of 260 fluorescent images of positive cells (130 images of each antigen) were captured on the microscope (Nikon) under constant shooting conditions: ×20 objective lens, ×10 eyepiece, and exposure time of 1/2 s. Acquired images were analyzed by using ImageJ software to calculate the average pixel intensity of each cell,

Qualitative and quantitative comparisons of authentic and rNP-based antigens of THAIV in IFA.
Focus reduction neutralization test
The endpoint titers of neutralizing antibodies against SEOV and THAIV were determined by the focus reduction neutralization test (FRNT) as described previously (Pattamadilok et al. 2006). We selected seropositive rodent sera GR2, GR22, GR90 and seronegative rodent sera GR10. In brief, 140 μL of serial twofold dilutions of serum were mixed with an equal volume of virus suspension containing 100 focus-forming units of virus at 37°C for 1 h. Fifty microliters of the mixture was inoculated into Vero E6 cell monolayers in 96-well tissue culture plates (Corning Incorporated, Costar, ME). After being incubated for 7 days in a CO2 incubator, the monolayers were fixed and stained. The FRNT titer was expressed as the reciprocal of the highest serum dilution, which resulted in a >80% reduction in the number of infected cell foci.
Results
Species identification of rodents
As shown in Table 1, the majority (112/116) of rodents were considered to be R. rattus by appearance and measurement data. Among them, 13 rats were selected, and their cytb gene was sequenced (LC426026-34, LC426358, and LC426359). From phylogenetic analysis, these sequences were found in lineage Ib, which is a Sri Lankan endemic lineage of R. rattus (Yasuda et al. 2014). One of the rodents, which was sequenced by using cytb, irbp, and rag1 genes and identified as B. bengalensis (LC426360, LC426364, and LC426368), was also considered to be the same lineage of B. bengalensis captured in Kandy, Sri Lanka (Yasuda et al. 2014). Two rodents were identified as M. booduga by sequencing of cytb, irbp, and rag1 genes (LC426362, LC426363, LC426366, LC426367, LC426370, and LC426371). They were captured as a distinct lineage from Indian M. booduga. Even though the sequences having significant identity, were determined from the mouse GR59 by using cytb (LC426361), irpb (LC426365), and rag1 (LC426369), they were not found in DNA databases used to determine the species. GR59 was a young adult female rodent captured inside a grocery store. Its profile is as follows: head and body length, 6.4 cm; tail length, 10.4 cm; body weight, 10 g; hind foot length, 1.8 cm; ear, 1.3 cm; and skull, 2.3 cm. The overall results suggested that R. rattus is the dominant rodent species in Girandurukotte. Nevertheless, other endemic lineages and species of rodents were also found in this area.
Rodent Species and Seroprevalence of Thailand Orthohantavirus
Immunofluorescent profiles in IFA tests
IFA profiles of authentic and rNP-based antigens of THAIV-expressing Vero E6 cells are shown in Figs. 2A and 3. Monoclonal antibody E5G6 labeled with Alexa Fluor 594 fluorescent dye was used for detection of NP in the IFA (Fig. 2A[1], A[4]). The distribution pattern of NP in authentic antigen of THAIV-infected Vero E6 cells was fine granular or fibrous. In contrast, the distribution pattern of rNP in THAIV Vero E6 cells was regular or homogeneous fluorescence across the cytoplasm. Transfection efficiency of Vero E6 cells was estimated to be ∼5–10%. These immunofluorescence patterns in the cytoplasm for both antigens were observed by rat serum experimentally inoculated with THAIV (Fig. 2A[2], A[5]). Similar positive patterns were observed in THAIV authentic antigen stained with serum of rodent ID, GR2, GR22, and GR90 (Fig. 3[1–3], respectively) as well as rNP-based antigen stained with serum as in preceding (Fig. 3[5–7], respectively). On the contrary, no specific fluorescent patterns against authentic or rNP-based antigens were observed in laboratory rat serum without inoculation (Fig. 2A[3], A[6]), and similar pattern observed with rodent serum of GR100 (Fig. 3[4], 3[8]).

IFA profiles of rodent sera against authentic and rNP-based antigens of THAIV. Prototype THAIV authentic antigen
As shown in Fig. 2B, the average intensity of images of rNP-based antigen of THAIV-expressing Vero E6 cells was 17.8 and that of authentic antigen was 5.3. These results indicate that the intensity of immunofluorescence images of rNP-based antigen of THAIV-expressing Vero E6 cells was three times higher than that of the authentic antigen.
Reactivities of rodent sera to authentic and rNP-based antigens in the IFA
A total of 116 rodents' sera were screened for THAIV antibodies in the IFA with authentic and rNP-based antigens (Table 1). According to the results obtained by both IFA tests which were on agreement, a total of 23 (19.8%) sera were positive while 93 (80.2%) of them were negative. To compare antibody titers, 23 seropositive sera were selected. As shown in Table 2 and Fig. 2C, IFA titers of sera to rNP antigen were higher than those to the authentic antigen.
Immunofluorescence Assay Antibody Titers of Rodent Sera Against Hantavirus Antigens
<100.
THAIV, Thailand orthohantavirus; HTNV, Hantaan orthohantavirus; SEOV, Seoul orthohantavirus; PUUV, Puumala orthohantavirus; IFA, immunofluorescence assay.
Seropositivity in each rodent species
As shown in Table 1 and Fig. 1, 23 of the 116 rodents captured in various regions of Girandurukotte were seropositive. Among the 23 seropositive rats, 22 rats were R. rattus. The only B. bengalensis was captured, which was also recognized as seropositive. In contrast, two M. booduga and Murinae sp. rodents were seronegative. Therefore, R. rattus and B. bengalensis are candidates of host rodent species in this area.
Antibody titers to different antigens
IFA antibody titers of rodents' sera against authentic antigen of THAIV were 100–3200. Those to SEOV and HTNV were 100–200, but those to PUUV were <100 (Table 2). Overall, IFA titers of rodent sera against SEOV, HTNV, and PUUV were lower than those to THAIV antigen. On the contrary, rat sera experimentally inoculated with authentic antigen of THAIV or SEOV showed high cross-reactivities to THAIV, SEOV, and HTNV. These results suggested that the hantavirus present in rodents in Sri Lanka is related to THAIV rather than to SEOV, HTNV, or PUUV. For further antigenicity comparison, four R. rattus sera were selected and FRNTs were performed (Table 3). Seropositive sera (GR2, GR22, and GR90) and seronegative serum (GR10) showed low FRNT titers (<20) to both THAIV and SEOV.
Analysis of Rodent Serum Samples in Focus Reduction Neutralization Test Using Thailand Orthohantavirus and Seoul Orthohantavirus
Antibody positive to THAIV shown in Table 3.
Pattamadilok et al. (2006).
Discussion
In this study, we found high seroprevalence to hantavirus among rodents in Girandurukotte, which is known to be a hotspot of CKDu in Sri Lanka. As mentioned previously, high seroprevalence to THAIV or TRHV infection among CKDu patients in Girandurukotte was observed (Yoshimatsu et al. 2019). Hantaviruses are usually carried by rodents, thereby this study aimed to capture the rodents residing in Girandurukotte area.
For molecular biological detection of hantavirus from rodents, lung tissues were collected from all rodents and immersed in RNALater (Qiagen). RNA fractions were extracted from the tissues and examined by RT-PCR using universal primer sets for the L genome segment (Klempa et al. 2006) and S segment (Arthur et al. 1992) and other original primer sets. Despite numerous attempts, obtaining any molecular biological information proved futile, possibly due to a lack of specific primers or failure to maintain the cold chain for sample transportation.
We carried out a serological survey for rodents by using IFA for which the preparation of authentic antigen of THAIV requires a specific containment facility such as BSL-3. In this study, we observed that the IFA image intensity of the rNP-based antigen of THAIV was higher than that of the authentic antigen when tested with a monoclonal antibody directed to NP. It gives the certainty to replace authentic antigen with rNP-based antigen of THAIV Vero E6 cells in IFA as an ideal nonbiohazardous serological diagnostic method for hantavirus infection in Sri Lankan rodents.
Although Vitarana et al. (1988) reported SEOV infection among brown rats (R. norvegicus) at port area in Colombo, Sri Lanka, no brown rats were captured in Girandurukotte, where the dominant rodent species were black rats (R. rattus). While commensals such as black rats prefer to inhabit around human residences, lesser bandicoot rats (B. bengalensis) tend to live in farming fields in close association with humans. We set traps for rodents only in houses and their surroundings, which resulted in the capturing of mostly black rats. Consequently, we identified two seropositive rodent species: black rat and lesser bandicoot rat. Furthermore, molecular biological analyses of rodent genes showed that both black rats and bandicoot rats in Girandurukotte belong to the Sri Lankan-specific lineage reported previously (Yasuda et al. 2012).
Unfortunately, in this survey, we could not get permission for rat trapping in rice field to avoid the capture of preserved wildlife. To clarify the host rodents in this CKDu endemic area, it is essential to capture rats not only indoors but also outdoors.
Pattamadilok et al. (2006) reported that the greater bandicoot rat (B. indica) is the species with the highest seroprevalence against THAIV in Thailand. On the contrary, in some countries such as Cambodia, Indonesia, Mayotte Island, and Madagascar, THAIV was detected from black rats (Reynes et al. 2003, 2014, Plyusnina et al. 2009, Johansson et al. 2010, Filippone et al. 2016). These results showed the distribution of THAIV and TRHV infections in the area of the Indian Ocean, including Sri Lanka, where no HFRS case has been reported.
In our previous study, sera from CKDu patients in Girandurukotte showed cross-reactivities against prototype THAIV and SEOV in IFA and FRNT (Yoshimatsu et al. 2019). On the other hand, rats in Girandurukotte showed a lower cross-reaction pattern to SEOV in FRNT than those of CKDu patients. A similar difference in cross-reactivities in humans and rodents was reported for vole-borne hantaviruses (Tegshduuren et al. 2010). Sera from PUUV-infected HFRS patients showed high cross-reactivities against Tula orthohantavirus (TULV). In contrast, sera from voles experimentally inoculated or naturally infected with PUUV and TULV showed type-specific antibody reactions. In the future, molecular biological identification of hantavirus-infected CKDu patients and rats in Girandurukotte will be required to confirm hantavirus transmission from black rats or bandicoot rats to humans in Sri Lanka. For this purpose, it is necessary to establish molecular biological diagnostic methods such as real-time PCR. In addition to results from IFA, under detectable level of FRNT to THAIV were shown. These results indicate that Sri Lankan hantavirus might be serologically distinct from the prototype THAIV and SEOV used in this study. Therefore, further serological studies are needed to discuss serological profile of THAIV and TRHV.
In conclusion, black rats and lesser bandicoot rats belonging to Sri Lankan endemic lineages were shown to be possible reservoirs for THAIV or TRHV infection in Girandurukotte. Also, the hantavirus present in rodents in Sri Lanka is related to THAIV rather than to SEOV, HTNV, or PUUV. However, it might be serologically distinct from the prototype THAIV.
Footnotes
Acknowledgments
We acknowledge SES English Service for revising the grammar in the final draft. This work was partially supported by a JSPS KAKENHI grant (number 15K08796) and U.S.-Japan Cooperative Medical Sciences Program Collaborative Award (AMED grant number 18950142), JSTA Sakura Science Plan (S2017F0228119) received by K.Y., and a grant provided by the National Science Foundation of Sri Lanka (RPHS/2016/CKDu/06) received by Prof. Rohana Chandrajith (Study 03 PI—Dr. C.D.G.), Department of Geology, Faculty of Science, University of Peradeniya, Sri Lanka.
Author Disclosure Statement
No conflicting financial interests exist.