
Abstract
Getah virus (GETV) is a mosquito-borne alphavirus that is considered to be an emerging pathogen. To date, reverse transcription loop-mediated isothermal amplification (RT-LAMP) has not been used to detect GETV. Therefore, we describe a novel, fast, and sensitive LAMP method to detect GETV. Amplification of GETV RNA can be obtained within 50 min at 65°C. This RT-LAMP method was verified to be highly specific for GETV, with no cross detection of other viruses. The assay was 103 and 101 times more sensitive than RT-PCR and RT-qPCR, respectively, for the detection of GETV RNA. This novel RT-LAMP method provides a practical and economical alternative for detecting GETV in mosquitoes and serum samples that can be used even in the field.
Introduction
Getah virus (GETV) is an arbovirus in the genus Alphavirus and the family Togaviridae. This family comprises single-stranded positive-sense RNA viruses. GETV is transmitted by mosquitoes, and vertebrate animals can act as amplifying hosts during seasonal outbreaks (Bannai et al. 2016, Yang et al. 2018). GETV can cause rashes, fever, and swollen legs in horses; miscarriage and death in pigs; and even fever in humans (Yago et al. 1987, Li et al. 1992, Nemoto et al. 2015). Therefore, GETV can cause significant losses in animal husbandry and human health. There is a need for sensitive techniques to allow the detection of GETV at an early stage, even before the onset of symptoms. Several diagnostic assays have been developed to detect GETV, including virus isolation and virus neutralization (VN) testing (Shibata et al. 1991, Bannai et al. 2016), hemagglutination (HA) assays (Shibata et al. 1991), transmission electron microscopy (TEM) (Li et al. 2017), RT-PCR (Nemoto et al. 2015), and RT-qPCR (Sam et al. 2018, Shi et al. 2018). However, these methods are time consuming, lack sensitivity, or require specialized laboratory facilities and personnel training.
Loop-mediated isothermal amplification (LAMP) was developed by Notomi et al. (Notomi et al. 2000). LAMP has high specificity and sensitivity and is simple to use, with no need for specialized equipment. LAMP is performed with Bst DNA polymerase (i.e., modified DNA polymerase from Bacillus stearothermophilus), which has strand displacement activity. The temperature ranges from 60°C to 65°C, and the assay is completed within 30–60 min under isothermal conditions. The final amplification product is a mixture of stem-loop DNAs with different stems that can be analyzed by direct observation or through agarose gels (Mori et al. 2001, Nagamine et al. 2002). The LAMP method was developed to detect a variety of viruses such as Japanese encephalitis virus (JEV) (Liu et al. 2012), foot-and-mouth virus (Yamazaki et al. 2013), dengue virus (Parida et al. 2005, Hu et al. 2015), West Nile virus (Cao et al. 2016), and Batai virus (Liu et al. 2016) but not GETV. In this study, we developed an RT-LAMP procedure that has promising potential as a tool to test for the presence of GETV in cattle serum samples and mosquitoes.
Materials and Methods
Cells and viruses
GETV strain JL17/08 was isolated from field-caught mosquitoes and preserved in our laboratory (Shi et al. 2018). The virus was propagated in Vero cells, and cultured supernatants were harvested at 4 days postinfection, clarified through centrifugation at 12,000 g for 30 min. JEV (YN0901 strain), Akabane virus (AKAV, NM/BS/1 strain), pseudorabies virus (PRV, DL14/08 strain), Batai virus (BATV, NM/12 strain), porcine reproductive and respiratory syndrome virus (PRRSV, CC-1 strain) or porcine circovirus 2 (PCV2, NM2002 strain), and Sindbis virus (SINV, YN1008 strain) were isolated and identified by our laboratory, were stored at −80°C until use.
RNA extraction and reverse transcription to cDNA
GETV RNA was extracted from 200 μL of Vero cell culture supernatant containing 105 plaque-forming units of either GETV strain JL17/08, JEV, AKAV, PRV, BATV, PRRSV, PCV2, and SINV using a QIAamp DNA/Viral RNA Mini Kit (Qiagen). The quantity and purity of the GETV RNA was determined by spectrophotometry and expressed as the number of molecular copies. The resultant virus RNA was reverse transcribed into cDNA using a Vazyme HiScript II 1st Strand cDNA Synthesis Kit (Vazyme Biotech Co., Ltd, China) (Shi et al. 2018).
Primer design
We used Primer Explorer version 5 software to design two pairs of primers from a conserved region of the GETV nsP1 gene deposited in GenBank for the RT-LAMP assay, whereas RT-PCR and RT-qPCR primers were previously described (Shi et al. 2018). All of the primers used in this study were analyzed with BLASTn (
Location of forward/reverse outer primer and forward/reverse inner primers in the GETV nsP1 gene. The length and position of the primers were marked with underline. The primers included two outer primers (forward outer primer and reverse outer primer) and two inner primers (forward inner primer, F1c+F2 and reverse inner primer, B1c+B2).
Sequences of Primers Used for RT-LAMP, RT-PCR, and RT-qPCR Analysis
RT-LAMP assay optimization
LAMP was performed in a 25-μL mixture comprising the following: 1.0 μL each of 5.0 μM F3 and B3 primers, 1.0 μL each of 40 μM forward and back inner primers, 2.5 μL of 10 × ThermoPol reaction buffer, 1.0 μL of 8 U/μL Bst DNA polymerase (New England Biolabs, Ipswich, MA), 2.5 μL of 10 mM dNTP mix (Takara, Dalian, China), 4.0 μL of 5.0 M betaine (Sigma-Aldrich, St. Louis, MO), 5.0 μL of 30 mM MgSO4 (Takara), 3.0 μL of target cDNA, and 3.0 μL of nuclease-free water. Reactions were conducted at 65°C for 10, 20, 30, 35, 40, 45, 50, 55, and 60 min to determine the optimal reaction time. The products were resolved through 2.5% (w/v) agarose gel electrophoresis. The RT-PCR and RT-qPCR assays were conducted as previously described (Shi et al. 2018).
Sensitivity and specificity of RT-LAMP detection
GETV RNA in a 10-fold dilution series, ranging from 2.61 × 105 to 2.61 × 10−1 copies/μL, was used as a reverse transcription (RT) template to evaluate and compare the sensitivity of the RT-LAMP, RT-PCR, and RT-qPCR assays. The following important infectious viruses of domestic animals that can cause similar disease in horse, pigs, or cattle, including JEV, AKAV, BATV, PRRSV, PRV, PCV2, and other representative mosquito-borne alphavirus such as SINV, were used to evaluate the specificity of the RT-LAMP assay.
Detection of GETV from field cattle serum samples and mosquitoes by the RT-LAMP assay
A total of 25 serum samples from cattle and 50 pools of Culex tritaeniorhynchus mosquitoes (100 mosquitoes/tubes) were selected based on morphological criteria and a taxonomic key. The mosquitoes were mechanically grinded in 2 mL of PBS. The supernatant was transferred to a fresh microtube after centrifugation at 12,000 g for 30 min at 4°C. Then, the supernatant was used to confirm the accuracy of the RT-LAMP, RT-qPCR, and RT-PCR assays. The accuracies of the RT-LAMP, RT-qPCR, and RT-PCR methods were verified by virus isolation and identification as previously reported (Shi et al. 2018).
Results
Optimization of GETV RT-LAMP
The RT-LAMP assay reactions were run for varying lengths of time, which showed that RT-LAMP at 65°C with a 50 min reaction time yielded appropriate apparent band intensities (Fig. 2).
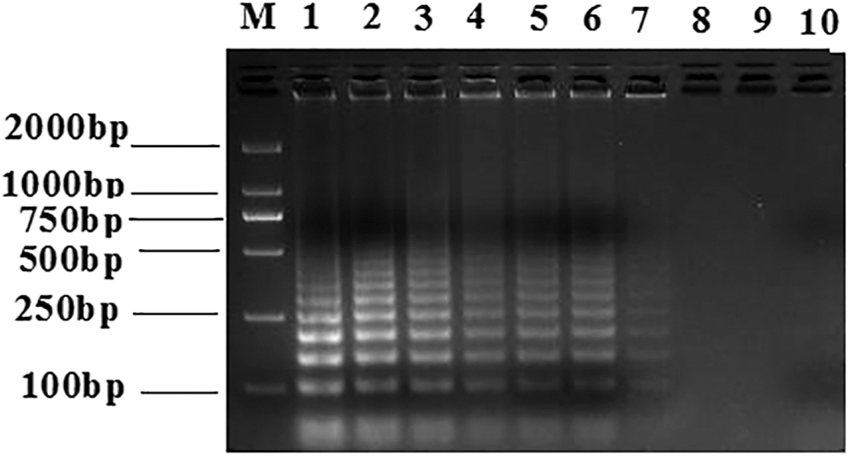
Test for optimized time conditions for RT-LAMP detection of the GETV nsP1 gene. The reaction products were resolved through agarose gel. The RT-LAMP reactions were performed at 65°C for 60, 55, 50, 45, 40, 35, 30, 20, or 10 min (Lanes 1–9, respectively). Lane 10, negative control. M, DNA marker.
Sensitivity of the RT-LAMP assay
The detection limits of the RT-LAMP assay, observed through 2.5% agarose gel electrophoresis or the addition of 1.0 μL of SYBR Green I (Invitrogen) under ultraviolet transillumination, revealed the intensities of the amplification products (Fig. 3A, B). The GETV RNA detection limit for RT-LAMP was 2.61 × 100 copies/μL, whereas it was 103 copies/μL for RT-PCR and 101 copies/μL for RT-qPCR (Shi et al. 2018). The results indicated that the RT-LAMP assay was 10 and 1000 times more sensitive than the RT-qPCR and RT-PCR assays, respectively (Table 2).

The RT-LAMP assay sensitivity for GETV RNA detection. GETV RNA concentrations from 2.61 × 105 to 2.61 × 10–1 copies/μL shown as
Detection Sensitivities of RT-LAMP, RT-PCR, and RT-qPCR According to GETV RNA (Copies/μL)
+, positive reaction; −, negative reaction.
Specificity of RT-LAMP detection
The specificity of RT-LAMP detection was determined with RNA samples from GETV, JEV, AKAV, BATV, PRRSV, PRV, PCV2, and SINV that were isolated in China. The reaction was performed at 65°C for 50 min. No amplification products were observed when cDNA/DNA templates from JEV, AKAV, BATV, PRRSV, PRV, PCV2, and SINV were used (Fig. 4). This indicated that RT-LAMP is highly specific for GETV.

Specificity of the RT-LAMP assay for detection of Getah virus. RT-LAMP reaction products were resolved on an agarose gel. Lane 1: GETV. Lanes 2–8, AKAV, BATV, JEV, PRRSV, PRV, PCV2, and SINV, respectively; Lane 9, negative control.
Detection of GETV in cattle serum samples and mosquitoes
The RT-LAMP assay showed that eight positive samples, including three samples from mosquito (Nos. M15, M27, and M41) and five cattle serum samples (Nos. B5, B11, B16, B18, and B23), tested positive for the presence of GETV RNA. However, four (Nos. M41, B11, B18, and B23) and two (Nos. B11 and B23) samples tested negative for the presence of GETV RNA by RT-PCR and RT-qPCR, respectively. The conventional RT-PCR and RT-qPCR assays did not detect GETV RNA in two samples (Nos. B11 and B23), which might be because their sensitivities are limited and the viral RNA could not be detected (Table 3). The RT-LAMP assay showed good repeatability across three trials.
Detection of GETV RNA in the Mosquito Pools and Cattle Serum Samples from Jilin, Northeastern China by RT-PCR, RT-LAMP, RT-qPCR, and Virus Isolation
+, positive reaction; −, negative reaction.
CPE, cell pathology effect.
Initially, the 50 mosquito suspensions and 25 serum samples were propagated in Vero cells (Shi et al. 2018). The analysis showed a cell pathology effect that is specific for GETV in the three mosquito suspensions and five cattle serum samples. These results were confirmed by RT-LAMP. None of the samples that tested positive for the presence of GETV RNA using RT-PCR and RT-qPCR were undetected by RT-LAMP. All negative samples tested negative for GETV RNA according to all the tests. Finally, we amplified two complete genes from GETV isolates from mosquitoes and cattle that were then sequenced and submitted to GenBank (Nos. MH722255 and MH722256).
Discussion
GETV has a broad geographical distribution and strong host adaptability, as shown by molecular and serological surveillance (Shibata et al. 1991, Sugiyama et al. 2009, Nemoto et al. 2015, Li et al. 2017). It has caused several disease outbreaks in pigs and horses for the past 20 years (Yago et al. 1987, Nemoto et al. 2015, Yang et al. 2018). The present method to monitor GETV has some insufficiencies, such as its high cost, time-consuming nature, complex operation, and inaccuracy. Therefore, a novel method to monitor GETV in remote rural areas is urgently needed.
In this study, we are the first to describe a fast, sensitive, and specific method performed in a single tube and at constant temperature to detect the GETV virus using RT-LAMP. Unlike RT-PCR and RT-qPCR, this method does not require a thermal cycler or expensive equipment, and the amplification process is completed in 50 min. The amplification products of the RT-LAMP reaction produce characteristic ladder-like patterns through agarose gel electrophoresis. Using SYBR Green I stain, the amplification products are detectable by naked-eye inspection under ultraviolet transillumination.
In previous reports, the sensitivity of RT-LAMP for arboviruses and animal viruses was higher than that of RT-qPCR and RT-PCR (Baek et al. 2018, Lopez-Jimena et al. 2018, Zheng et al. 2018, Xia et al. 2019). The detection limits for GETV were 101 and 103 copies/μL by RT-qPCR and RT-PCR assays, respectively (Shi et al. 2018). We have determined the detection limit of RT-LAMP by 10-fold serial dilutions of RNA from virus propagated in cell culture and evaluated the results relative to those from RT-PCR and RT-qPCR. However, the RT-LAMP detection limit for GETV was similar to that for other arboviruses: 2.61 copies/μL GETV RNA could be detected (Kim et al. 2018). The specificity of the RT-LAMP assay was good, as no positive results were found using RNA/DNA from other viruses such as AKAV, BATV, JEV, PRRSV, PCV2, PRV, and SINV, which can cause similar disease in horse, pigs, or cattle, and mosquito-borne alphavirus. The study limitation of only SINV of Alphavirus genus members was used to evaluate the specificity of the RT-LAMP assay; other alphaviruses such as EEEV, VEEV, and Semliki forest virus were not available for testing. However, sequence alignment between the GETV nsP1 gene sequences and other Alphavirus genus members confirmed that RT-LAMP used with these primers would be a low possibility of detecting other alphaviruses. The evaluation of mosquito suspensions and cattle serum for the presence of GETV RNA revealed the repeatability and reliability of this assay. The key to the success of the LAMP method was the use of two pairs of primers that corresponded to conserved regions of the nsP1 gene in GETV.
Conclusions
In this study, we developed an RT-LAMP method for the visual detection of GETV RNA that is useful and economical for monitoring and controlling GETV. This method is especially advantageous for use in developing countries, where expensive and technically demanding equipment and techniques are not readily available, but epidemiological surveillance is necessary to detect or confirm disease outbreaks and limit their spread.
Footnotes
Acknowledgments
This study was supported by the National Natural Sciences Foundation of China (31802199), the National Key Research and Development Program of China (2017YFD0500104), and the Key Laboratory for Preventive Research of Emerging Animal Diseases in Foshan University (KLPREAD201801-07).
Author Disclosure Statement
No competing financial interests exist.