
Abstract
Introduction:
Borrelia burgdorferi sensu lato (s.l.), the causative agent of Lyme borreliosis, is the most prevalent tick-borne pathogen in Europe, where it is mainly transmitted by Ixodes ricinus. This tick also circulates Borrelia miyamotoi, a member of the relapsing fever group of species.
Objectives:
A longitudinal study was performed to assess the prevalence of Borrelia spp. in questing ticks. Relationships between Borrelia prevalence in ticks and some variables such as tick development stage, sampling area, and questing tick density were assessed; in addition, the distribution of B. burgdorferi s.l. was analyzed to establish the periods of acarological risk.
Methods:
Ticks were collected monthly by flagging in three different ecological areas (coast, plateau, and mountain) in north-western Spain during a 2-year study. Borrelia DNA was detected by PCR, targeting the flagellin (fla) gene. Positive samples were also characterized at the rrfA-rrlB intergenic spacer region and the glycerophosphodiester phosphodiesterase (GlpQ) gene.
Results:
B. burgdorferi s.l. DNA was detected in 11.84% of I. ricinus. Five Lyme Borrelia species were identified (Borrelia afzelii, B. burgdorferi sensu stricto, Borrelia garinii, Borrelia lusitaniae, and Borrelia valaisiana). One single relapsing fever species (B. miyamotoi) was detected (0.85%). Questing Ixodes frontalis, Ixodes acuminatus, Dermacentor reticulatus, and Dermacentor marginatus yield negative results. The prevalence of B. burgdorferi s.l. was significantly higher in female ticks and in the mountain area. In addition, a seasonal pattern in the B. burgdorferi s.l. prevalence distribution throughout the study was not detected.
Conclusions:
The detection of a noticeable prevalence of B. burgdorferi s.l. in questing I. ricinus suggests a high acarological risk, especially in mountain area. There is no evidence of a relationship between the prevalence of B. burgdorferi s.l. and the density of questing ticks, nor traces of a seasonal pattern in the values of prevalence in ticks.
Introduction
Tick-borne diseases are a worldwide public health concern (Dantas-Torres et al. 2012). Ticks are the second most important vectors of pathogens to humans following mosquitoes, but they rank first considering the diversity of carried pathogens (Jongejan and Uilenberg 2004). In this regard, Borrelia burgdorferi sensu lato (s.l.), the causative agent of Lyme borreliosis (LB), is the most prevalent tick-borne pathogen in Europe (Barandika et al. 2008).
B. burgdorferi s.l. is mainly transmitted by ticks belonging to the Ixodes ricinus complex. I. ricinus is the main vector in Europe (Gern and Humair 2002). Up-to-now, 10 B. burgdorferi s.l. species have been identified in Europe: Borrelia garinii and Borrelia afzelii are the most frequent and associated with neuroborreliosis and atrophic chronic acrodermatitis, respectively. Borrelia bavariensis, Borrelia bissettiae, Borrelia finlandensis, Borrelia lusitaniae, Borrelia spielmanii, B. burgdorferi sensu stricto (s.s.), and Borrelia valaisiana are less common (Tappe et al. 2014). In addition, Borrelia miyamotoi, a member of the relapsing fever group, and Borrelia turcica, a species associated to reptiles, have been also found in ticks from Europe (Takano et al. 2012).
It has been reported that the distribution of vector-borne diseases is conditioned by the presence of suitable tick vectors and reservoirs (Estrada-Peña and de la Fuente 2014). Environmental conditions should also be considered since some of them could affect tick mortality rates or timing of tick activity. Thus, several investigations have suggested a relationship between temperature or vapor pressure deficit and the prevalence of B. burgdorferi s.l. in ticks (Gatewood et al. 2009, Estrada-Peña et al. 2011), probably as a consequence of the contact rates between ticks and reservoirs (Estrada-Peña and de la Fuente 2014). The knowledge of factors driving the prevalence of Borrelia in ticks is of major interest (Mysterud et al. 2013). Thus, field studies are the main source of information relating to the distribution of B. burgdorferi s.l. with the questing activity of their tick vectors.
LB is one of the most prevalent vector-borne diseases in Europe (van den Wijngaard et al. 2017). In Spain, LB cases have been detected in all provinces, with an average incidence of 0.25 cases per 100,000 inhabitants and year (Bonet-Alavés et al. 2016); nevertheless, the highest incidence values were detected in northern Spain, considered an endemic area (Hofhuis et al. 2015, Vázquez-López et al. 2015, Bonet-Alavés et al. 2016). Galicia, located in the northwest, is one of the Spanish regions showing the highest incidences of LB, especially the province of Lugo (2.96/100,000 inhabitants and year) where high percentages of questing ticks were also positive to B. burgdorferi s.l. (Vázquez-López et al. 2015, Bonet-Alavés et al. 2016, Díaz et al. 2017); in fact, a noticeable increase of LB incidence in humans has been detected in this region from 2007 (2.64 cases/100,000 inhabitants/year) to 2012 (11.61 cases/100,000 inhabitants/year) (Vázquez-López et al. 2015). Thus, we performed a 2-year longitudinal study to assess the prevalence of Borrelia spp. in ticks collected monthly in three different ecological areas from north-western Spain. The analysis of the monthly distribution of B. burgdorferi s.l. prevalence throughout the study will allow capturing the acarological risk in space and time. We further focused on the effect of some variables such as the sampling area and the density and stage of development of questing ticks on the percentage of ticks infected by B. burgdorferi s.l.; this information will be useful for identifying risk areas and periods for acquiring LB, leading to the optimization of diagnosis and treatment of the disease.
Materials and Methods
Study area
Field studies were conducted in three different ecological regions (coast, plateau, and mountain) (Lago et al. 2012) of Galicia, a 29,574 km2 region located in north-western Spain (Fig. 1), from November 2015 to October 2017. This region has an Oceanic-Atlantic climate with mild temperatures and high precipitation. Grasses such as the meadow fescue (Festuca pratensis), Kentucky bluegrass (Poa pratensis), and ray grass (Lolium perenne) are the most common herbaceous plants, and oaks (Quercus robur) and sweet chestnut (Castanea sativa) are the major tree species (Pato et al. 2013). One single representative sampling area was chosen in each ecological region; distinctive meteorological and topographical features of each area are summarized in Fig. 1.

Localizations and geoclimatic characteristics of the three sampling areas.
Tick collection and identification
Ticks were collected monthly by flagging in each sampling area. A 1 m2 piece of white cotton flannel was passed over the vegetation along the same 300 meters transect. Ticks were picked up from the blanket at 5–10-meter intervals and stored in tubes with 70% ethanol. A total of 6956 questing ticks were collected. Ticks were identified to species and stage based on morphological features (Pérez-Eid 2007, Estrada-Peña et al. 2017). Five species were found: I. ricinus (652 larvae, 5767 nymphs, 227 males, and 258 females), Dermacentor marginatus (4 males and 15 females), Dermacentor reticulatus (7 males and 10 females), Ixodes frontalis (1 larva and 14 nymphs), and a single male of Ixodes acuminatus. Tick specimens were stored in 100% ethanol at −20°C until DNA extraction.
DNA extraction and detection of Borrelia spp.
The minimum number of ticks to be processed for DNA extraction was calculated with a 95% confidence interval and a 5% type II error to detect differences of 10% in the prevalence of Borrelia spp. in ticks between sampling areas. We thus analyzed 652 nymphs, 202 females, and 202 males of I. ricinus, together with 12 I. frontalis nymphs and all collected D. marginatus, D. reticulatus, and I. acuminatus.
Each tick was longitudinally cut with a sterile scalpel and homogenized in 200 μL of Tissue Lysis Buffer using a tissue homogenizer (MagNaLyser; Roche, Germany) at 6000 rpm during 60 s. DNA was extracted using a commercial kit (High Pure PCR Template Preparation Kit; Roche Diagnostics GmbH®, Mannheim, Germany), following the manufacturer's instructions. Samples were analyzed by a nested PCR targeting the flagellin (fla) gene, using previously described protocols (Barbour et al. 1996, Stromdahl et al. 2003). Positive samples were selected and sequenced; sequences were aligned and edited using ChromasPro (Technelysium, Brisbane, Australia), and consensus sequences were scanned against the GenBank database using the Basic Local Alignment Search Tool (BLAST 2019;
Statistical analysis
The possible influence of the sampling area and the stage of development of ticks on the prevalence of B. burgdorferi s.l. in I. ricinus was analyzed using a logistic regression. Factors were eliminated from the initial model using a backward and forward conditional method based on the Akaike Information Criterion value to build the best model, and all pairwise interactions were evaluated. Odds ratio values were computed by raising “e” to the power of the logistic coefficient over the reference category. In addition, the influence of the studied variables on the prevalence of each B. burgdorferi s.l. species was analyzed by means of a chi-squared test. The possible relationship between the density of I. ricinus and the prevalence of B. burgdorferi s.l. in each season was assessed using a Spearman correlation. All statistical analyses were performed using the statistical software R (R Core Team 2018).
Results
Borrelia spp. DNA (fla gene) was detected in 130 out of 1056 I. ricinus ticks (12.31%). No Borrelia-positive I. frontalis, I. acuminatus, D. reticulatus, and D. marginatus were detected. All fla-positive samples were successfully sequenced; in addition, all PCRs targeting the IGS or the GlpQ gene were positive, and sequence analysis confirmed fla results. Lyme Borrelia species were predominant since they were found in 125 positive ticks (96.15%), whereas relapsing fever species were detected in 9 I. ricinus (6.92%). Mixed infections with both Lyme and relapsing fever Borrelia species were detected in three nymphs and one I. ricinus female (3.07%). Five B. burgdorferi s.l. species plus B. miyamotoi were identified. B. afzelii was the most frequently recorded, followed by B. garinii and B. valaisiana. B. burgdorferi s.s., B. lusitaniae, and B. miyamotoi were only occasionally found (Table 1). The detected co-infections included B. miyamotoi-B. valaisiana (n = 2) and B. miyamotoi-B. garinii (n = 2). Most Borrelia spp. sequences were identical to reference sequences deposited in the GenBank database or showed a single-nucleotide polymorphism.
Prevalence of Borrelia burgdorferi sensu lato and Borrelia miyamotoi in Questing Ixodes ricinus Ticks from North-Western Spain Considering the Different Variables Studied
s.l., sensu lato; s.s., sensu stricto.
The total prevalence of LB spirochetes was significantly higher in the mountain area compared to plateau or coastal regions (Table 2). B. miyamotoi was detected in all sampling areas with prevalences lower than 2%. The prevalence of all B. burgdorferi s.l. species, except for B. valaisiana, was also highest in the mountain (Table 1), but these differences were only significant for B. afzelii (χ2 = 16.498; p < 0.001). In addition, the mountain showed the highest diversity of species, since all the five B. burgdorferi s.l. species and B. miyamotoi were identified (Table 1). B. lusitaniae was not found in either the plateau or coastal areas, whereas B. miyamotoi was not detected in the plateau area.
Logistic Regression Model for the Prevalence of Borrelia spp. Factors were Removed Following the Akaike Information Criterion Value Until the Best Model was Built
CI, confidence interval; OR, odds ratio.
All identified Borrelia species were found in nymphs and adults of I. ricinus (Table 1). The individual prevalence of B. burgdorferi s.l. was significantly higher in females than in nymphs or males (Table 2). Similarly, B. miyamotoi and all B. burgdorferi s.l. species were more frequent in females, except for B. burgdorferi s.s. that was more prevalent in males (Table 1) without significant differences (p > 0.05). No correlation between tick density and Borrelia spp. prevalence was found with the Spearman correlation test (p > 0.05). A seasonal pattern of prevalence of Borrelia spp. in questing I. ricinus ticks was not found (Fig. 2). Positive ticks were found throughout the year regardless the area of collection.
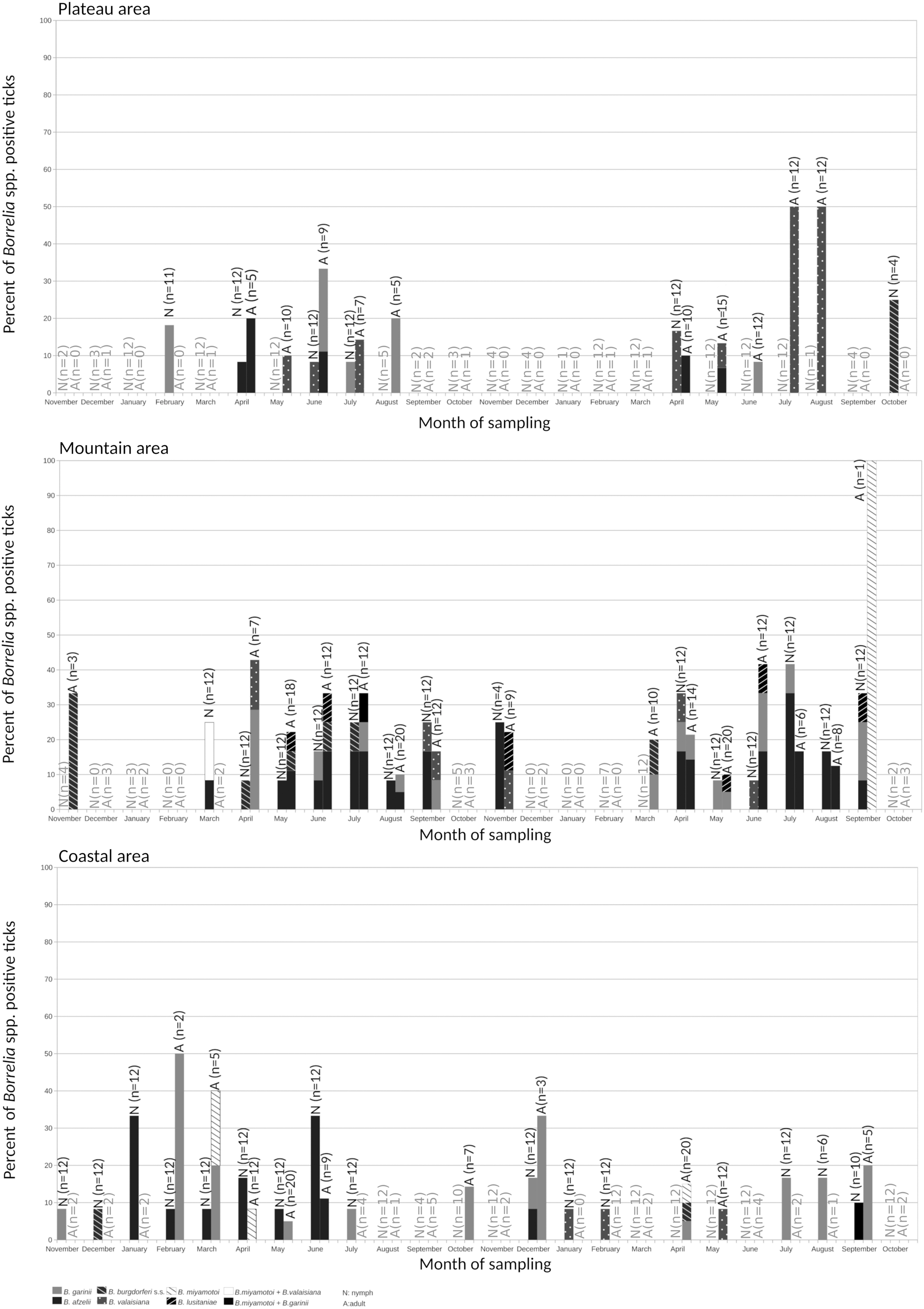
Borrelia spp. prevalence in questing Ixodes ricinus ticks during both years of study.
Discussion
Our data reveal that I. ricinus is the most abundant questing tick and the only carrier of B. burgdorferi s.l. in north-western Spain, agreeing with the many studies carried out in Europe (Gern and Humair 2002, Strnad et al. 2017). Density of other tick species was low, and B. burgdorferi s.l. remained undetected in I. acuminatus, I. frontalis, D. reticulatus, and D. marginatus. I. frontalis is an ornithophilic tick found in both nests and vegetation and considered a competent vector of B. burgdorferi s.l. (Estrada-Peña et al. 1995, Heylen et al. 2013). In addition, the presence of B. burgdorferi s.l. DNA in questing Dermacentor spp. from Europe has been reported in a limited number of investigations, with lower prevalence than that detected in I. ricinus and never exceeding 3.4% (Bonnet et al. 2013, Reye et al. 2010, Zając et al. 2017). It has been suggested that those results may be due to the inactivation of B. burgdorferi s.l. spirochetes by the immune system of Dermacentor since these ticks are not considered competent vectors (Johns et al. 2001, Grubhoffer et al. 2005).
Our results confirm that B. burgdorferi s.l. is prevalent and widely distributed in questing I. ricinus in Galicia (north-western Spain). These bacteria are considered endemic in other northern regions of Spain, with largely variable prevalence ranging from 0.6% to 7.1% in nymphs and from 2.1% to 6% in adults (Barral et al. 2002, Espí et al. 2017). Our results showed that the epidemiological situation in Galicia involves a great risk for transmission of LB agents to humans, paralleling the increasing incidence of patients with LB recently reported in this region (Vázquez-López et al. 2015).
Significant variations in the prevalence of B. burgdorferi s.l. in questing ticks among variable ecological conditions were already reported (Gray et al. 1995, Jouda et al. 2004, Richter et al. 2013). Therefore, our results fill the gap providing human health authorities with the prevalence of Borrelia spp. in questing ticks. These data pinpoint the significantly higher prevalence of B. burgdorferi s.l. in the mountain, where the highest incidence of LB was previously reported (Vázquez-López et al. 2015). It seems that the correct conditions for spatial overlapping of vectors and reservoirs occur in this area, where the highest altitude (>650 meters) and lowest average annual temperature values (<10.9°C) were recorded (Fig. 1). These differences could be related to variables such as climate, density of reservoirs, land use, and human actions on landscape driving habitat fragmentation. It has been reported that most of these variables locally affect the prevalence of Borrelia spp. in questing ticks (Estrada-Peña and de la Fuente 2014). As an example, higher prevalence of B. burgdorferi s.s. and B. garinii was reported in ticks collected in oak forests than those from pine forests (Gray et al. 1998, Tack et al. 2011, Ruyts et al. 2017) because the former provide a better microclimate for ticks and show a higher density of suitable reservoirs. Our study addressed the importance of the qualitative features of landscape, considering only the differences observed among the three surveyed ecosystems. Obvious differences in the prevalence of Borrelia spp. were found, but a rigorous analysis of the impact of climate variables on the activity of local reservoirs warrants future assessment.
We did not find a statistical correlation between the density of questing nymphs and adults of I. ricinus and the prevalence of B. burgdorferi s.l. in ticks. In addition, the prevalence of B. burgdorferi s.l. in I. ricinus was similar through both years of study and a pattern on the Borrelia prevalence distribution was not detected, agreeing with previous reports (Jouda et al. 2004, Estrada-Peña et al. 2011, James et al. 2012). Nevertheless, other studies suggested a seasonal pattern in both questing ticks positive to Borrelia spp. (Talleklint and Jaenson 1996) and LB patients (Kurtenbach et al. 2006); these investigations were performed in European regions with harsh winters, suggesting that different variables, especially low winter temperatures, may negatively affect the activity of local reservoirs leading to a decrease in the percentage of Borrelia-positive questing ticks. In contrast, less marked seasonal climatic fluctuations, such as those recorded in other European regions, including north-western Spain, allow a more constant reservoir activity throughout the year and thus may explain the lack of a seasonal pattern in the prevalence of B. burgdorferi s.l. in questing ticks. Considering the absence of this temporal pattern, the risk of acquiring LB will only increase with tick density due to a higher risk of exposure to tick bites.
The prevalence of B. burgdorferi s.l. was significantly higher in female I. ricinus than in nymphs or males, as reported in other studies carried out in Europe (Jouda et al. 2004, Kampen et al. 2004, Soleng et al. 2013, Sormunen et al. 2015, Espí et al. 2017), a feature already linked to the number of blood meals or even the amount of blood consumed during the feeding process (Matuschka et al. 1993, Dusbábek 1996).
Borrelia garinii and B. afzelii were the predominant species, matching results of a recent study carried out in Asturias, an adjacent area from north-western Spain (Espí et al. 2017). Both species were also most commonly reported in human patients from Galicia (Vázquez-López et al. 2016a, b). These differences in the ecological distribution of B. burgdorferi s.l. species have been related to the presence of a particular reservoir composition, since these spirochetes infect different groups of animals (Kurtenbach et al. 2002, Estrada-Peña et al. 2011). Thus, B. afzelii and B. burgdorferi s.s. are associated with rodents (Huegli et al. 2002, Hanincová et al. 2003), B. valaisiana and B. garinii with birds (Hanincová et al. 2003), and B. lusitaniae with lizards (Richter and Matuschka 2006). These relationships reflect the extremely local regulatory factors that affect the circulation of the species of B. burgdorferi s.l.
The finding of B. miyamotoi in both nymphal and adult I. ricinus from coastal and mountain areas confirms that this species is circulating in north-western Spain. Since this spirochete was identified as pathogenic for humans (Wagemakers et al. 2015), its presence in the studied areas should be considered in the differential diagnosis of clinical patients (Palomar et al. 2018).
Conclusions
The noticeable prevalence of B. burgdorferi s.l. in questing I. ricinus ticks from three different ecological areas in north-western Spain suggests a high acarological risk, especially in those regions showing the highest altitude and lowest average annual temperature. In addition, the prevalence of B. burgdorferi s.l. in questing I. ricinus was not related to the density of questing ticks in the studied areas. There is no evidence of a monthly or seasonal pattern in the prevalence of B. burgdorferi s.l. in questing ticks. Consequently, the acarological risk for acquiring LB increases only with the density of questing ticks. Further research assessing the abundance and relative contribution of reservoirs in the area and their role circulating the pathogens is needed. Such information could be used to determine and predict risk areas for acquiring LB.
Footnotes
Acknowledgments
This research was supported by the Programme for Consolidating and Structuring Competitive Research Groups (GRC2015/003; Xunta de Galicia, Spain) and by a predoctoral grant (European Social Fund, Secretaría Xeral de Universidades, Xunta de Galicia).
Author Disclosure Statement
No competing financial interests exist.