
Abstract
Cowpox virus (CPXV), genus Orthopoxvirus, family Poxviridae, is a zoonotic pathogen in Eurasian wild rodents. High seroprevalences have been reported previously for vole and murine species in Europe. In contrast, viral DNA was only rarely detected, and very few reservoir-derived CPXV isolates exist. In this study, CPXV DNA and CPXV-reactive antibodies were monitored in wild small mammals for 5 years in four German federal states. Screening of liver tissues of 3966 animals by CPXV real-time PCR (qPCR) revealed five voles of two species positive for CPXV DNA. Two positive bank voles (Myodes glareolus) and two positive common voles (Microtus arvalis) originated from two plots in Baden-Wuerttemberg. One positive bank vole originated from Mecklenburg-Western Pomerania. None of the small mammals from Thuringia and North Rhine-Westphalia was positive in the qPCR. CPXV antigen-based indirect immunofluorescence assays of 654 highly diluted chest cavity fluid samples detected two bank voles and two common voles from the same sites in Baden-Wuerttemberg to be highly seroreactive. Five animals were CPXV DNA positive, and four other animals were orthopoxvirus seropositive. Our study indicates both a very low prevalence and a patchy occurrence of CPXV in common and bank voles and absence in other rodent and shrew species in Germany. The multiple detection of infected voles at one site in Baden-Wuerttemberg and continued detection in a region of Mecklenburg-Western Pomerania classify these regions as potential endemic foci.
Cowpox virus (CPXV), family Poxviridae, genus Orthopoxvirus, is a reemerging zoonotic pathogen in Europe (Essbauer et al. 2010). CPXV has a broad host range in Western Eurasia, for example, cats, pet and farm animals, and various zoo animals representing accidental hosts (Essbauer et al. 2010). The transmission to humans is mainly mediated by contact to infected cats, wild rodents, or pet rats (Essbauer et al. 2010, Hoffmann et al. 2015). Contaminated fomites might play an eminent role in the transmission of CPXV and could serve as infectious source for human infection (Andreani et al. 2019).
Serological and molecular surveys in Eurasian regions confirmed that various rodent species may act as natural reservoirs for CPXV (Essbauer et al. 2010). Despite high orthopoxvirus seroprevalence rates in wild rodents, molecular detection and isolation of CPXV are rarely reported (Martina et al. 2006, Kinnunen et al. 2011, Franke et al. 2017, Prkno et al. 2017). Previous investigations in Germany indicated orthopoxvirus-reactive antibodies in bank voles (Myodes glareolus) and the striped field mouse (Apodemus agrarius) and resulted in the molecular detection of CPXV in bank voles, common voles (Microtus arvalis), and Norway rats (Rattus norvegicus) (Kurth et al. 2008, Kinnunen et al. 2011, Franke et al. 2017).
Here, we evaluated small mammals from Germany for the presence of CPXV DNA and orthopoxvirus-reactive antibodies. During 2010–2014, 4023 small mammals including 3306 rodents, 714 shrews, 2 least weasels (Mustela nivalis), and 1 European mole (Talpa europaea) were collected at four sites (Fischer et al. 2018; Fig. 1 and Table 1). Locations represented different geographic regions, with Mecklenburg-Western Pomerania and North Rhine-Westphalia exhibiting a more maritime climate and Baden-Wuerttemberg and Thuringia a more continental one. A mosaic of forest fragments within an agriculturally dominated landscape characterized all locations. Trapping sites were established in forests and grasslands (Fig. 1) to ensure that a broad spectrum of species, representative of the respective area, was caught.
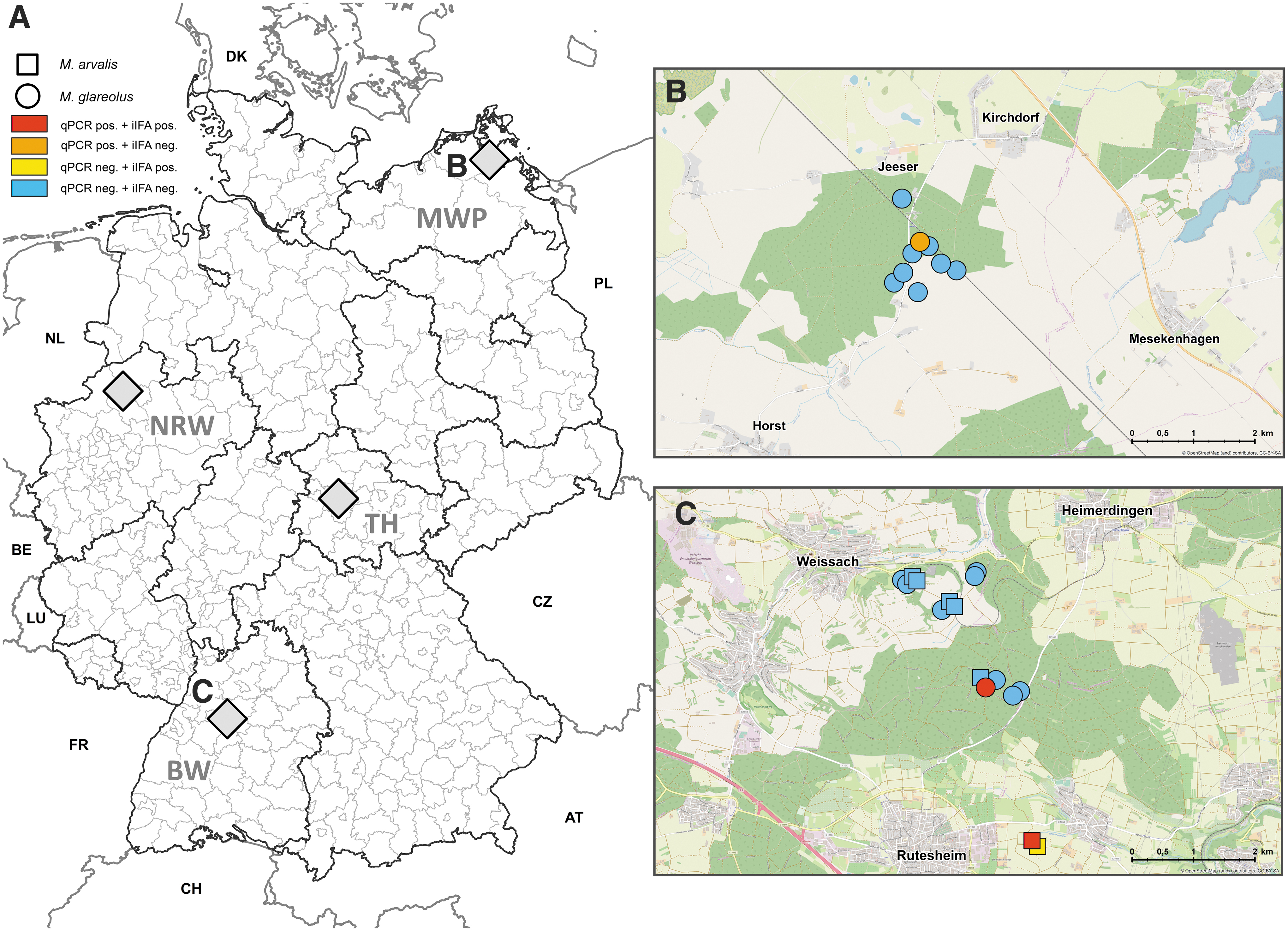
Location of four monitoring regions in Germany
Detection of Cowpox Virus-Specific DNA and Orthopox Virus-Reactive Antibodies Antibodies in Small Mammals Collected During 2010 to 2014 in Four Federal States of Germany by Real-Time PCR and Indirect Immunofluorescence Assay
Species with positive animals in one of the assays are indicated in bold letters.
Includes CPXV isolate FM2292 (Hoffmann et al. 2015).
Includes single specimens of Greater white-toothed shrew (Crocidura russula) and European mole (Talpa europaea).
—, not analyzed; CI, confidence interval; CPXV, cowpox virus; iIFA, indirect immunofluorescence assay; qPCR, real-time PCR.
DNA was extracted from liver tissue of 3966 small mammals and investigated by a real-time PCR (qPCR) assay according to the published standard protocols (Hoffmann et al. 2015). Overall, the qPCR analysis resulted in five positive samples (Table 1 and Supplementary Tables S1 and S2): 2/381 common voles (0.52%, 95% CI [confidence interval]: 0.06–1.88%) and 3/1594 bank voles (0.19%, 95% CI: 0.04–0.55%). The qPCR-positive common voles originated from one plot in Baden-Wuerttemberg (22.22%, 95% CI: 2.81–60.01%) and one plot in Mecklenburg-Western Pomerania (1.92%, 95% CI: 0.05–10.26%; Fig. 1B, C and Supplementary Table S1). From one of the qPCR-positive common voles from Baden-Wuerttemberg, the CPXV isolate FM2292 was previously generated and further characterized (Hoffmann et al. 2015). The two positive bank voles originated from one plot in Baden-Wuerttemberg (0.83%, 95% CI: 0.1–2.97%; Supplementary Table S1).
For serological analysis, the chest cavity of small mammals was washed with 1 mL phosphate-buffered saline. CPXV-based indirect immunofluorescence assay (iIFA) analysis of highly diluted chest cavity fluids (according to Hoffmann et al. 2015; for details, see legend of Fig. 1) of 654 small mammals from the CPXV-positive sites in Baden-Wuerttemberg and Mecklenburg-Western Pomerania resulted in two positive common voles and two positive bank voles, all from Baden-Wuerttemberg, with titers ≥100 (Fig. 1C, Table 1, and Supplementary Tables S1 and S2). The two seropositive bank voles originated from the same plot where two bank voles were qPCR positive (2/243, 0.82%, 95% CI: 0.1–2.97%; Fig. 1 and Supplementary Tables S1 and S2). The seropositive common voles originated from two other neighboring plots in Baden-Wuerttemberg (2/103, 1.94%, 95% CI: 0.24–6.84%; Supplementary Tables S1 and S2). The analyzed voles were either CPXV DNA positive (n = 5) or seroreactive (n = 4) with prevalences below 1% (Table 1 and Supplementary Tables S1 and S2). Males and females were affected by CPXV infection, and all positive animals were adults (weight >19 g; Supplementary Table S2). Furthermore, the CPXV-positive animals were either bank voles of the Eastern and Western evolutionary lineages or common voles (Supplementary Table S2).
CPXV dynamics showed a very site-specific distribution with no expansion to sites >100 meters away over the course of the study period (Fig. 1B, C). In addition, when CPXV persists in common or bank voles at a particular site, none of the other occurring rodents and shrews was tested positive in the qPCR or iIFA (Supplementary Fig. S1). This and the generally very low DNA detection rate is in line with previous molecular CPXV detection studies in voles and reports about CPXV isolation from voles (Kinnunen et al. 2011, Hoffmann et al. 2015, Franke et al. 2017, Prkno et al. 2017). According to previous investigations (Hoffmann et al. 2015, Miranda et al. 2017), liver was, at the time point of this investigation, the most promising tissue for the detection of CPXV DNA. Very recently, we realized that samples from the upper respiratory tract might represent a better target for these investigations (Prkno et al. 2017).
However, the very low seroprevalence rate observed here is in clear contrast to previous studies in bank voles, common voles, field voles (Microtus agrestis), the striped field mouse, and the wood mouse (Apodemus sylvaticus) (Crouch et al. 1995, Boulanger et al. 1996, Tryland et al. 1998, Chantrey et al. 1999, Hazel et al. 2000, Essbauer et al. 2009, Kinnunen et al. 2011). The absence of CPXV infections in field voles is also in contrast to studies in Great Britain where seroprevalence rates can reach 100% and field voles seem to be an endemically infected CPXV reservoir (Cavanagh et al. 2004, Burthe et al. 2006). Reasons could be the use of highly diluted chest cavity fluid samples instead of serum and a higher cutoff value of ≥1:100 applied in our study as it was introduced previously (Franke et al. 2017). Chest cavity fluid samples have to be used here because sera of small mammals were not available due to the trapping procedure. The high dilution was required to allow simultaneous investigations for additional pathogens, for example, hantaviruses (Drewes et al. 2017). In addition, a lower dilution of chest cavity fluid was found to produce nonspecific reactions that limit the results of the analysis (Hoffmann, personal communication). However, we cannot rule out that the analysis of a serum sample might have tested positive for an animal that was found to be negative in iIFA for the chest cavity fluid sample.
The detection of CPXV DNA in Mecklenburg-Western Pomerania confirms previous detections of orthopoxvirus-reactive antibodies in bank voles, striped field mice, and Norway rats from the same region (Kinnunen et al. 2011, Heuser et al. 2017). In this region, the transmission of CPXV from a Norway rat to a zoo elephant (Elephas maximus) was also reported previously (Kurth et al. 2008). Similarly, CPXV was repeatedly detected in voles from closely located sites in Baden-Wuerttemberg and was isolated from common voles at one site from this study and from another site in Baden-Wuerttemberg (FM2292 and Ger/2007/vole) (Hoffmann et al. 2015, Franke et al. 2017). The complete genomes of both strains are genetically closely related and belong to the CPXV-like clade 2 (Franke et al. 2017).
The absence of a parallel detection of CPXV DNA and CPXV-reactive antibodies confirms previous investigations and may suggest virus clearance instead of persistent infections. The detection of CPXV infections in adults only may indicate an age-related infection risk irrespective of sex. However, due to the low number of positive animals, the influence of demographic, seasonal, and spatial factors on the individual infection status could not be determined. Furthermore, the restriction to only very few collection spots might be also a reason for the low detection rate. Putative reservoir small mammals were collected in temporal and spatial proximity to CPXV-DNA-positive or anti-orthopoxvirus-antibody-positive voles but were tested negative. This temporal and spatial proximity of potentially susceptible animals with infected and/or seropositive voles and their low frequency may suggest that the transmission of CPXV, at least in the regions investigated in this study, is rather rare. The observed low prevalence and the likely absence of persistent infections raise questions about the reservoir function of voles.
In conclusion, our monitoring study indicated a low prevalence of CPXV in both common voles and bank voles and complete absence in other small mammal species. The multiple detection of infected voles at one site in Baden-Wuerttemberg and continued detection in a region of Mecklenburg-Western Pomerania suggest these regions as potential endemic foci. Future investigations might give a more comprehensive and reliable information on CPXV prevalence in reservoir hosts in Germany (and elsewhere) by using respiratory tract samples for DNA detection and sera for antibody detection. In addition, future studies should evaluate the stability of CPXV in the environment as a main factor of transmission.
Footnotes
Acknowledgments
We thank Doris Junghans and Mareen Lange for excellent technical assistance.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
The work was funded in part by the German Federal Ministry of Education and Research (BMBF) through the German Research Platform for Zoonoses (“NaÜPa-net,” FKZ 01KI1018 and 01KI1303) and through the consortium InfectControl 2020 (“RAPiD,” FKZ 03ZZ0821A) awarded to R.G.U., by Deutsche Forschungsgemeinschaft (SPP 1596, BE5187) awarded to M.B., and partly commissioned and funded by the Federal Environment Agency (UBA) within the Environment Research Plan of the German Federal Ministry for the Environment, Nature Conservation, Building and Nuclear Safety (BMUB) (grant no. 371348401) awarded to J.J.
Supplementary Material
Supplementary Figure S1
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.