
Abstract
Ixodes scapularis, the black-legged tick, harbors multiple organisms and transmits several pathogens to animals and humans. To determine the presence of tick-borne microorganisms carried by I. scapularis in Pennsylvania, 299 adult I. scapularis ticks were collected from across the state and tested with a multiplex bead panel targeting 20 microorganisms. The Luminex bead-based xMAP® MultiFLEX Mega Tick Panel detected microorganisms in these ticks, including Anaplasma spp. (1.7%), Borrelia spp. (45.8%), Babesia spp. (16.1%), and Rickettsia spp. (22.1%) at the genera level and identified Anaplasma phagocytophilum (1.7%), Babesia microti (0.7%), Borrelia burgdorferi sensu stricto (45.5%), Borrelia miyamotoi (0.3%), and Rickettsia parkeri (0.7%) at the species level. Babesia spp. reactivity was found to be due to Ba. odocoilei, and Rickettsia spp. reactivity was mainly due to rickettsial endosymbionts.
Introduction
I
Methods for diagnosing tick-borne diseases in humans and animals include microscopic blood smear examinations, serology, Western blot, and PCR (Sanchez et al. 2016). Microscopy has diagnostic value for organism-based detection, but the interpretation of results is subjective. Multiplex real-time PCR provides rapid and specific detection, but the number of pathogens a specimen can be screened for is limited by fluorescence, and machine capabilities usually to no more than five targets per reaction. Multiplex bead-based detection technology and next-generation sequencing have emerged as important tools for detecting multiple microorganisms in a sample (Tokarz et al. 2019). The Luminex® technology offers a novel platform that readily allows bead-based multiplexing, and is ideally suited for high-throughput detection. Use of the technology has been described for a variety of applications, including pathogen, antibody, and cytokine detection (Dunbar 2006). The xMAP® MultiFLEX Mega Tick Panel (GenArraytion, Inc., Rockville, MD) was developed to detect a wide variety of tick-borne pathogens found globally, including 15 species-level targets and 7 targets that detect multiple species within a particular genus. Detection of different microbial pathogens simultaneously, including species-level determination within the Borrelia, Babesia, Anaplasma, Rickettsia, and Ehrlichia genera, can be a useful diagnostic tool for tick testing and evaluating human and animal infections.
The aim of the current study was to evaluate the use of the xMAP MultiFLEX® Mega Tick Panel through a survey of 299 ticks, collected from Pennsylvania, for 20 genomic targets, including bacteria, protozoans, and viruses, associated with tick-borne disease (Table 1).
Microbial Pathogens Targeted in Adult Black-Legged Ticks (Ixodes scapularis) with a Luminex Multiplex Bead Assay
Materials and Methods
Ticks
Adult I. scapularis ticks were collected opportunistically by dragging a cloth material across the low-lying ground cover and under vegetation on the edges of trails, where people would be most likely to encounter questing ticks, during the spring and fall activity periods of 2013. The tick collection sites included county parks, state parks, state game lands, state forest lands, and other similar habitats where deciduous forests predominated. For this study, the counties of Pennsylvania were separated into 6 regions (Northcentral, Southcentral, Northeast, Southeast, Northwest, and Southwest) and an effort was made to randomly select up to 10 ticks from each of the 5 representative counties from each region (299 ticks selected out of 1,363 total ticks) for analysis of multiple genetic targets. The ticks were stored at room temperature in 70–80% ethyl alcohol until processing. As previously reported, these collected ticks were then identified morphologically and placed individually into 1.5-mL cryotubes with a drop of ethyl alcohol before nucleic acid extraction (Keirans and Litwak 1989, Hutchinson et al. 2015).
Nucleic acid extraction
Nucleic acids were extracted using an E.Z.N.A. tissue DNA kit, which copurifies DNA and RNA (OMEGA Bio-Tek, Norcross, GA), according to the manufacturer's recommendation with some modifications. An individual tick was placed on a glass slide, cut into four pieces with a disposable blade, transferred to a 1.5-mL cryotube, and incubated at 55°C overnight in an Eppendorf thermomixer at 400 rpm with TL buffer and proteinase, followed by centrifugation at 10,000 rpm for 3 min. Buffer BL was added to the supernatant and incubated for 10 min at 70°C. Nucleic acid was precipitated with absolute ethanol, then washed with HB buffer and DNA washing solution, and captured on a filter. The filter was dried at 14,000 rpm for 2 min, and nucleic acids were reconstituted in 50 μL of elution buffer and stored at −25°C before analysis (Hutchinson et al. 2015).
PCR analysis
The extracted specimens were screened for tick-borne microorganism nucleic acids using an xMAP MultiFLEX Mega Tick Panel with the MAGPIX® instrument (Luminex, Austin, TX). The xMAP MultiFLEX Mega Tick Panel includes 22 beads coupled with cap oligonucleotides specific for DNA and RNA targets associated with tick-borne microorganisms. Fifteen of the beads detect one species and seven detect multiple species in a particular genus (genus level), but only 20 of the 22 targets were used in the analysis. The two Borrelia spp. targets were not included in the analysis as the preassessment of the sensitivity and specificity data from the assay development showed that the genus-level assay was not as sensitive as species-level identification. Borrelia spp. data were derived from the number of positives for both B. burgdorferi and B. miyamotoi. The panel also includes a nonspecific binding control, fluorescence control, and an instrument control bead to monitor assay performance. Positive controls were obtained from GenArraytion, Inc., and negative controls were also included in all runs. The reverse transcriptase and PCR steps were carried out as per the xMAP MultiFLEX Mega Tick Panel package insert in a 25 μL reaction with 5X Qiagen One-Step RT-PCR buffer, a final concentration of 0.4 mM dNTP, 5 μL of xMAP MultiFLEX Primer Mix (Luminex), 1 μL of Qiagen One-Step Enzyme Mix (Qiagen, Germantown, MD), and 8 μL of nucleic acid template. The amplification was performed using a GeneAmp® PCR System 9700 (Applied Biosystems, Foster City, CA) with the following cycling parameters: 50°C for 30 min and then 95°C for 15 min, followed by 40 cycles of 95°C for 45 s and then 60°C for 45 s (Campagnolo et al. 2018).
The subsequent hybridization, labeling, and data acquisition were carried out according to the manufacturer's guidelines (Luminex). Briefly, the hybridization was carried out in a 50 μL reaction with 5 μL of Luminex magnetic beads that had immobilized beads, 35 μL of buffer A (Luminex), and 10 μL of PCR product. This mixture was heated at 95°C for 4 min followed by a 15-min incubation at 52°C to allow denaturation and renaturation with probes on beads. The Luminex beads and bound PCR product were pelleted using a magnetic plate separator. Labeling was then carried out by suspending the pellets in a mixture of streptavidin-conjugated R-phycoerythrin (SAPE) and buffer B and incubating at 52°C for 15 min. Following these steps, the plate was loaded on the MAGPIX for data acquisition. Test Data Analysis Software (TDAS) was used for data analysis with a target-specific signal/noise ratio of 5 FU as the positive threshold.
Positive specimens were further analyzed by stand-alone real-time PCR tests and Sanger sequence-based analysis with genus appropriate target regions (Eurofins, KY). A segment of 18S rRNA was targeted for sequencing the Babesia spp.-positive specimens (Michel et al. 2014). Primers targeting the first 500 base pairs of the 16S rRNA region were used for the sequencing of Rickettsia spp.-positive specimens (Tewari et al. 2011). Rickettsia parkeri was sequenced by GenArraytion, Inc. (Rockville, MD) targeting a 200 base pair region of the F pilus assembly protein TraF gene (GenBank acc. no. AFC74910.1).
All ticks included in the study were also previously tested for B. burgdorferi, A. phagocytophilum, and Ba. microti with singleplex assays as described previously (Hutchinson et al. 2015). The xMAP MultiFLEX Mega Tick Panel results were compared with the available results for concordance. Discrepant specimens were analyzed by a duplex PCR targeting B. burgdorferi and A. phagocytophilum as previously described, which also helped in assessing diagnostic sensitivity and specificity (Hojgaard et al. 2014).
The specificity and sensitivity data for the xMAP MultiFLEX Mega Tick Panel were provided by GenArraytion (Supplementary Table S1). No cross-reactivity between targets was observed. Primers used for sequencing and confirming with PCR are described in Supplementary Table S2.
Results
The xMAP MultiFLEX Mega Tick Panel detected target(s) in 180/299 (60.2%) of the ticks surveyed. Ticks carrying a single type of microorganism were most common (39.8%; 119/299), while 20.4% (61/299) of surveyed ticks carried more than one organism, thus identifying coinfections (Table 2). The maximum microbial targets in any tick analyzed with xMAP MultiFLEX Mega Tick Panel were 4, which included Rickettsia spp., Babesia spp., R. parkeri, and B. burgdorferi out of 20 total targets tested. The tick testing results were also concordant with the results of a previous study for the three tested targets (Hutchinson et al. 2015). All Ba. microti and A. phagocytophilum results from the xMAP MultiFLEX Mega Tick Panel were identical to the previous PCR results. The B. burgdorferi results matched, with the exception of results from 12 of the 299 ticks tested.
Detection of Microorganisms in Adult Ixodes scapularis Ticks Collected from Pennsylvania
Ticks were tested using the xMap® MultiFLEX Mega Tick Panel.
Multiple organisms (>1–4) detected.
Among the 20 microbial targets analyzed, the xMAP MultiFLEX Mega Tick Panel identified 5/299 (1.7%) tick specimens positive for Anaplasma spp. and A. phagocytophilum, 48/299 (16.1%) were positive for Babesia spp., 2/299 (0.7%) were positive for Ba. microti, 136/299 (45.5%) were positive for B. burgdorferi, 1/299 (0.3%) positive for B. miyamotoi, 66/299 (22.1%), positive for Rickettsia spp., and 2/299 positive for R. parkeri (0.7%) (Table 3). All tick specimens in this survey were negative for Bartonella spp., Borrelia afzelii, Borrelia garinii, Coxiella burnetii, Ehrlichia spp., E. canis, E. chaffeensis, Powassan virus, Rickettsia helvetica, Rickettsia japonica, Rickettsia rickettsii, and tick-borne encephalitis virus.
Detection and Distribution of Microorganisms and Coinfections in Ixodes scapularis Ticks Collected from Pennsylvania
Tick-borne microorganisms were detected either at the genusa or species levelb (A. phago.: A. phagocytophilum; B. burg.: B. burgdorferi; Ba. microti: Babesia microti using the xMap MultiFLEX® Mega Tick Panel from Ixodes scapularis (n = 299). Specimens were negative for targets Borrelia afzelii, Borrelia garinii, Ehrlichia spp., E. chaffeensis, E. canis, Powassan virus, R. rickettsii, R. japonica, R. helvetica, Bartonella spp., C. burnetii, and tick-borne encephalitis virus in Pennsylvania (PA) at the following locations Northcentral (NC), Southcentral (SC), Northeast (NE), Southeast (SE), Northwest (NW), and Southwest (SW). Rickettsial endosymbionts are not included in coinfections.
All Anaplasma spp.-positive specimens were identified as A. phagocytophilum, thus matching previous reported finding using a singleplex PCR. A. phagocytophilum-positive ticks were only recovered from the Northcentral region of Pennsylvania (Table 3). Sequencing of one of the A. phagocytophilum-positive specimens showed this particular strain to share 100% similarity with the following top 5 A. phagocytophilum strains: Norway variant 2 (GenBank acc. no. CP015376.1), dog 2 (GenBank acc. no. CP00668.1), JM (GenBank acc. no. CP006617.1), HZ2 (GenBank acc. no. CP006616.1), and HZ (GenBank acc. no. CP000235.1).
Two of the 48 tick specimens positive for Babesia spp. were identified as Ba. microti. These particular tick specimens were also previously reported as positive for Ba. microti with a singleplex PCR assay. The remaining 46 Babesia spp.-positive specimens were analyzed by real-time PCR, and sequencing of the 18S rRNA region for these Babesia specimens showed a 100% match to Ba. odocoilei (e.g., GenBank acc. no. M045131.1). Babesia were uniformly distributed geographically (Table 3).
Sixty-six of the surveyed ticks were positive for Rickettsia spp. R. rickettsii, R. japonica, and R. helvetica were not identified, but R. parkeri was detected from two ticks (0.7%); one in each of the Northwest and Northcentral regions. This finding was further confirmed with a real-time PCR followed by gene sequencing of the F pilus assembly protein TraF, with the nucleic acid showing a 100% match to R. parkeri strain Portsmouth (GenBank acc. no. CP003341.1; Fig. 1). In addition, 16S rRNA sequencing of 24 randomly selected Rickettsia spp.-positive specimens showed the Rickettsia spp. to belong to the nonpathogenic Rickettsial endosymbiont group (GenBank acc. no. D84558.1).
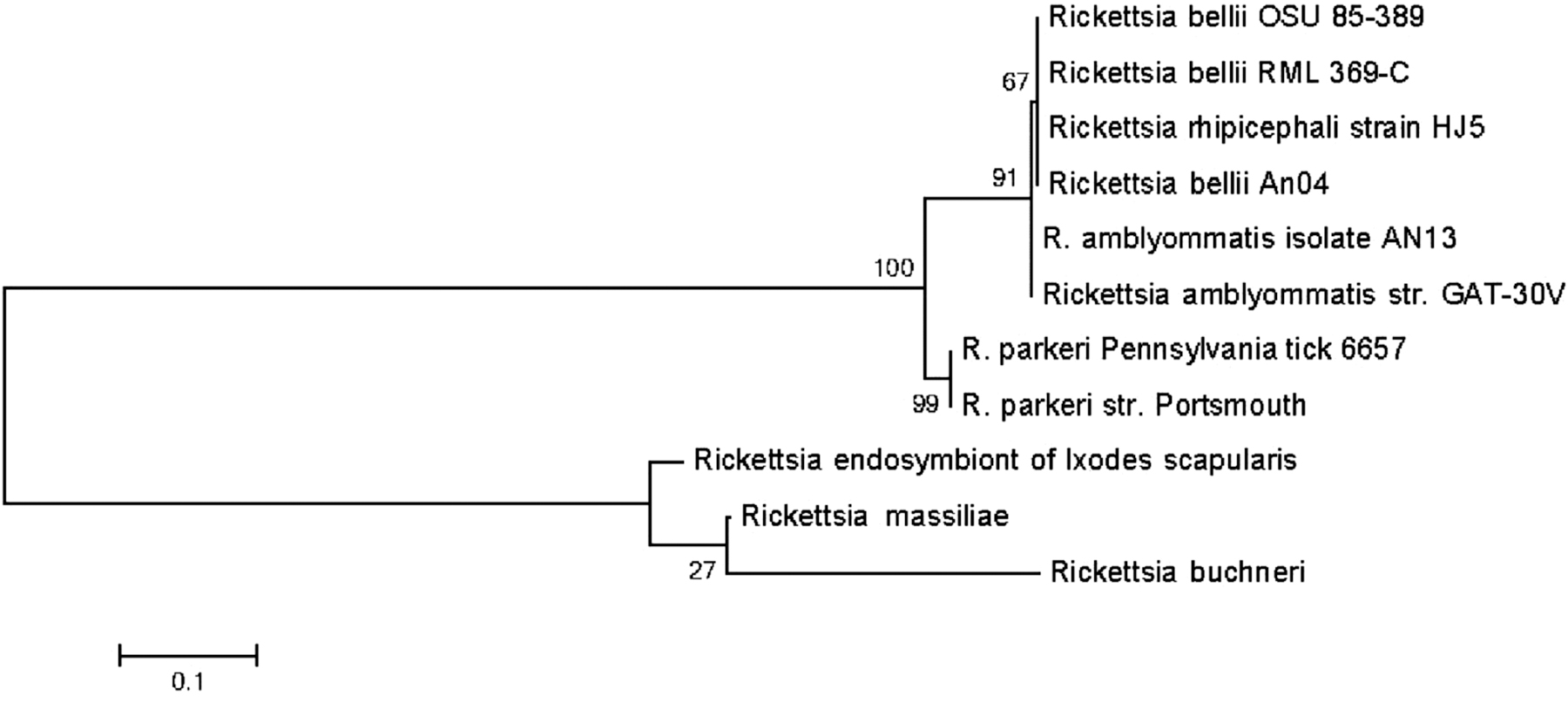
Phylogenetic relationship of Rickettsia parkeri with the F pilus assembly protein TraF gene sequencing. Phylogenetic tree was constructed with Mega software v. 6.06. The sequences were aligned using ClustalW, and the tree represents maximum likelihood with 500 bootstrap iterations with the bootstrap values at the node.
A substantial number of ticks were positive for B. burgdorferi (136/299; 45.1%), and a single tick (0.3%) was positive for B. miyamotoi with a combined detection rate of 45.5% at the Borrelia genus level. There were 12 mismatches between the results of multiplex bead assay and the previously published gel-based PCR for B. burgdorferi. Three ticks that were previously determined to be positive were negative by the xMAP MultiFLEX Mega Tick Panel, and eight previously negative samples were found to be positive with the xMAP MultiFLEX Mega Tick Panel. The discordant xMAP MultiFLEX Mega Tick Panel results of these 11 samples matched the results of the duplex real-time PCR assay. One B. burgdorferi-negative sample was determined to give a false-positive signal with the multiplex bead assay, and this finding was confirmed with the duplex real-time PCR. Despite a higher limit of detection noted, as shown in Supplementary Table S1 (Supplementary Data), test sensitivity for B. burgdorferi in the multiplex assay was 100% and specificity was 99.4%. B. burgdorferi was found to be uniformly distributed throughout the state of Pennsylvania, and B. miyamotoi was detected only from the Southeast region of the state (Table 3).
A combination of Babesia spp. and B. burgdorferi was found to be carried by 24 ticks (8.0%), a combination of A. phagocytophilum and B. burgdorferi was found in two ticks (0.7%), a combination of Ba. microti and B. burgdorferi was found in two ticks (0.7%), and a combination of B. burgdorferi and R. parkeri was found in one tick (0.3%). A single tick (0.3%) was determined to carry three tick-borne pathogens: Babesia spp., B. burgdorferi, and R. parkeri confirming coinfections.
Discussion
Tick-borne pathogens are increasingly being recognized as a risk for causing disease in humans and animals. In studying tick-borne microorganisms, the challenge often is the inability to study multiple agents beyond the analytical capabilities of existing assays that rely on multiplex real-time PCR (Tokarz et al. 2017). The Luminex bead-based technology offers the ability to analyze multiple targets with relative ease in a high-throughput format and can identify coinfections rather easily. To the authors' knowledge, this is the first report documenting the simultaneous screening of 20 genetic targets from microorganisms in I. scapularis ticks with a single multiplex assay. The multiplex bead assay was slightly more sensitive compared with the B. burgdorferi assay described by Hutchinson et al. (2015). Coinfections in ticks with multiple microorganisms were also detected with the assay and these can be medically important (Steiner et al. 2008, Hutchinson et al. 2015).
The assay determined ticks throughout the state to be positive for Rickettsia spp., but further analysis showed this Rickettsia spp. to be Rickettsia endosymbionts in the spotted fever group, which thus far have not been shown to play a direct role in human or animal disease and are vertically transmitted. The detection of the Rickettsia endosymbiont in only a fraction of the total ticks was concordant with other studies, where 46–65% detection rates were noticed (Steiner et al. 2008, Tokarz et al. 2019).
This is the first report of the detection of R. parkeri nucleic acids from I. scapularis in Pennsylvania, although there have been multiple confirmed cases of spotted fever and rickettsial pox in the state (Nelder et al. 2016). Two ticks tested positive for R. parkeri, which causes R. parkeri rickettsiosis, easily confused with the Rocky Mountain spotted fever caused by R. rickettsii or other spotted fever rickettsioses of undefined origin (Parola et al. 2013, Kelman et al. 2018). This was surprising as the primary vector of R. parkeri is Amblyomma maculatum, and R. parkeri is infrequently detected in I. scapularis ticks, although a previous study documented 18 I. scapularis found in Louisiana to be harboring R. parkeri (Nelder et al. 2016). Ticks may transfer microorganisms to other ticks while cofeeding, and the transfer of R. parkeri from A. maculatum to Amblyomma americanum during cofeeding has already been documented (Wright et al. 2015). Although A. maculatum has not been identified as far north as Pennsylvania to date, it is known to be established in the neighboring states of Maryland and Delaware, and the continued northward expansion in habitat of A. maculatum is expected (Sonenshine 2018). It is possible that A. maculatum has already begun to establish habitat in Pennsylvania, warranting the need for further investigation and monitoring.
The detection of Ba. microti (0.7%) was similar to a previously published detection rate of less than 0.5% from I. scapularis in Pennsylvania (Edwards et al. 2015). The 18S rRNA sequencing to further characterize the Babesia species from all positive specimens identified these as Ba. odocoilei, a tick-borne pathogen that is increasingly being recognized as a threat to cervids (Bartlett et al. 2009). To date, Ba. odocoilei has not been implicated in human disease, but Ba. odocoilei is closely related to other species of Babesia, which have zoonotic potential (Herwaldt et al. 2003). Continued surveillance of tick populations in Pennsylvania will help focus future efforts in recognizing, diagnosing, treating, and preventing babesiosis.
The prevalence of B. burgdorferi throughout Pennsylvania as determined by this study was very similar to previous studies, and the finding that nearly half of the adult I. scapularis surveyed carry B. burgdorferi is consistent with the fact that the Northeastern United States accounts for more than 95% of all documented Lyme disease cases in humans (CDC 2018). B. burgdorferi was also noticed to be part of coinfections in ticks. A single tick also tested positive for B. miyamotoi. Previous studies have also reported B. miyamotoi from areas of the Northeastern United States, including Pennsylvania; however, the prevalence of B. miyamotoi is reported to be much lower than that of B. burgdorferi (Edwards et al. 2015).
In the current study, A. phagocytophilum was only detected from five ticks in a single county in the Northcentral region with an overall prevalence of 1.7%. This low detection rate is similar to earlier studies reporting between 0.5% and 3.3% (Steiner et al. 2008, Edwards et al. 2015, Hutchinson et al. 2015) and low reported human infection rates in Pennsylvania (CDC 2018). Nationwide, the incidence of human anaplasmosis is rising, and although anaplasmosis is not as commonly reported in Pennsylvania as in more northern states, monitoring of A. phagocytophilum infections in humans, animals, and ticks is needed (Brown et al. 2015).
The I. scapularis ticks surveyed tested negative for Bartonella spp. and Powassan virus, which were previously reported in I. scapularis ticks collected from Pennsylvania (Nelder et al. 2016, Campagnolo et al. 2018). Interestingly, our previous study using a Luminex-based single bead-based assay had successfully detected Powassan virus, although at a very low level, using a different tick population in the same study area (Campagnolo et al. 2018). Such detection discrepancies among different studies can arise from tick stages surveyed, method of tick collection (collection of host-seeking ticks collected by dragging vs. ticks collected from reservoir hosts), engorgement levels of ticks, and molecular methods or target genes used for testing. As the xMAP MultiFLEX Mega Tick Panel was developed to have global use, the panel included targets that were not expected to be identified in the current study, due to either geographical limitations or vector competence. These targets, which tested negative, included B. afzelii, B. garinii, C. burnetii, E. canis, E. chaffeensis, R. helvetica, R. japonica, R. rickettsia, and tick-borne encephalitis virus.
Conclusion
Based on the results of this evaluation, the xMAP MultiFLEX Mega Tick Panel appears to be sensitive and specific for the screening of tick specimens to detect multiple microorganisms. With the completion of the initial evaluation using I. scapularis ticks, further testing of this panel will include cost comparisons with next-generation sequencing and multiplex PCR along with additional testing for tick-borne microorganisms in a variety of other tick vectors and host species from risk-based geographic areas in and out of Pennsylvania.
Footnotes
Acknowledgments
The authors are thankful for sequencing support received from Robert Vereen, Doreen Robinson, and R. Paul Schaudies of GenArraytion, Inc., Rockville, MD, for F pilus assembly protein TraF gene.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
Funding was received from the Pennsylvania Department of Agriculture and Pennsylvania Department of Health.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.