
Abstract
Background:
Zoophilic mosquitoes play an important role in the transmission of arboviruses of medical importance at human-wildlife interfaces, yet arbovirus surveillance efforts have been focused mostly on anthropophilic mosquitoes. Understanding the diversity of zoophilic mosquitoes and their associated feeding patterns and arboviruses can inform better vector control strategies.
Materials and Methods:
We morphologically identified mosquitoes collected from two game reserves in Kenya, the Maasai Mara National Reserve (MMNR) and locations near the Shimba Hills National Reserve (SHNR). Representative mosquitoes were also identified by cytochrome c oxidase subunit 1 (COI) barcode sequencing. In addition, we identified the vertebrate hosts of mosquito blood meals from the contents of each mosquito's abdomen by high-resolution melting (HRM) analysis and sequencing of COI, 16S ribosomal RNA, and cytochrome b gene PCR products. Similarly, mosquito arbovirus infections were identified by HRM analysis and sequencing of Alphavirus- and Flavivirus-specific RT-PCR products.
Results:
Of 2858 mosquitoes collected, 51 were engorged with blood meals from seven different vertebrate hosts, including humans, birds, domestic, and peridomestic animals and wildlife. Culex was the most abundant mosquito genus, with Culex pipiens being the most abundant species in both study regions. Among MMNR samples, we detected dengue serotype-2 virus (DENV-2) for the first time in Aedes tarsalis and Aedes tricholabis, as well as Sindbis virus in male Cx. pipiens. We also detected DENV-2 in Aedes aegypti sampled from locations near the SHNR. Human and diverse wildlife blood meals were identified, including bushbuck blood in the dengue-infected Ae. tarsalis and both human and hippopotamus blood in a single Eretmapodites chrysogaster mosquito.
Conclusions:
Our findings highlight the potential risk of sylvatic dengue and Sindbis transmission to humans by zoophilic mosquitoes at human-wildlife interfaces in Africa. Of specific importance, we provide evidence of sylvatic DENV-2 in Ae. tarsalis and Ae. tricholabis, representing potential new dengue vectors.
Introduction
Zoophilic mosquitoes (preferring animal blood to human blood) are important vectors that transmit emerging and re-emerging viruses from animals to humans, and thus pose a serious threat to global health and security (Conway et al., 2014). In sub-Saharan Africa, diverse Aedes spp., including Aedes aegypti, which is mainly an anthropophilic species on other continents (McBride et al., 2014), circulates in sylvatic and peridomestic cycles involving wild animals, including birds, domestic mammals, and humans (Sylla et al., 2009). Similarly, Culex mosquitoes mainly obtain blood meals from birds, although mammals are also involved and can also transmit arboviruses to humans (Omondi et al., 2015, Grubaugh et al., 2017). Despite playing an important role in the transmission of arboviruses of medical importance at human-wildlife interfaces, most zoophilic mosquitoes are understudied, with most surveillance efforts focusing on anthropophilic mosquitoes.
Over the last two decades, Kenya has been hit hard by outbreaks of arboviral diseases, including chikungunya fever (WHO, 2018), Rift Valley fever (RVF) (Baba et al., 2016a), and dengue fever (Baba et al., 2016b). Control strategies for these outbreaks are limited by a lack of vaccines, except for RVF and yellow fever, which have vaccines licensed for use in animals (Mansfield et al., 2015) and humans (Monath and Vasconcelos, 2015), respectively. Therefore, early detection of viruses in their vectors and control of vector populations remain indispensable for strategies curbing arboviral disease outbreaks.
In Kenya, as in many other resource-limited countries, systematic epidemiological surveillance of circulating arboviruses in their biological vectors is sporadic and limited in scope, especially in wildlife-livestock-human interfaces. Therefore, we focused on identifying the diversity of zoophilic mosquitoes and their vertebrate blood meals and associated flaviviruses and alphaviruses of public health importance at the human-wildlife interfaces of the Maasai Mara National Reserve (MMNR) in southwestern region of Kenya and the Shimba Hills National Reserve (SHNR) in the south coast of Kenya.
Materials and Methods
Mosquito sampling
We obtained sampling approval from the Kenya Wildlife Service (KWS) and Department of Health offices before sampling mosquitoes in the MMNR and from locations near the SHNR (Fig. 1). The two protected wildlife reserves were selected because of the encroaching human settlements along the reserves' borders and reported cases of human arboviral infections (Eastwood et al., 2017). The MMNR is a game reserve with diverse vertebrate wildlife that covers some 1510 km2 in Narok County, southwestern Kenya, and shares a border with the Serengeti National Park, Tanzania. The “Great Migration” between the Serengeti National Park and the MMNR of large populations of zebra (Equus quagga) and wildebeest (Connochaetes taurinus) was ongoing during the sampling period. The SHNR is a wildlife sanctuary located in Kwale County along Kenya's south coast. It is within the eastern African coastal forest belt and home to a variety of wildlife species, including the endangered sable antelope (Hippotragus niger).
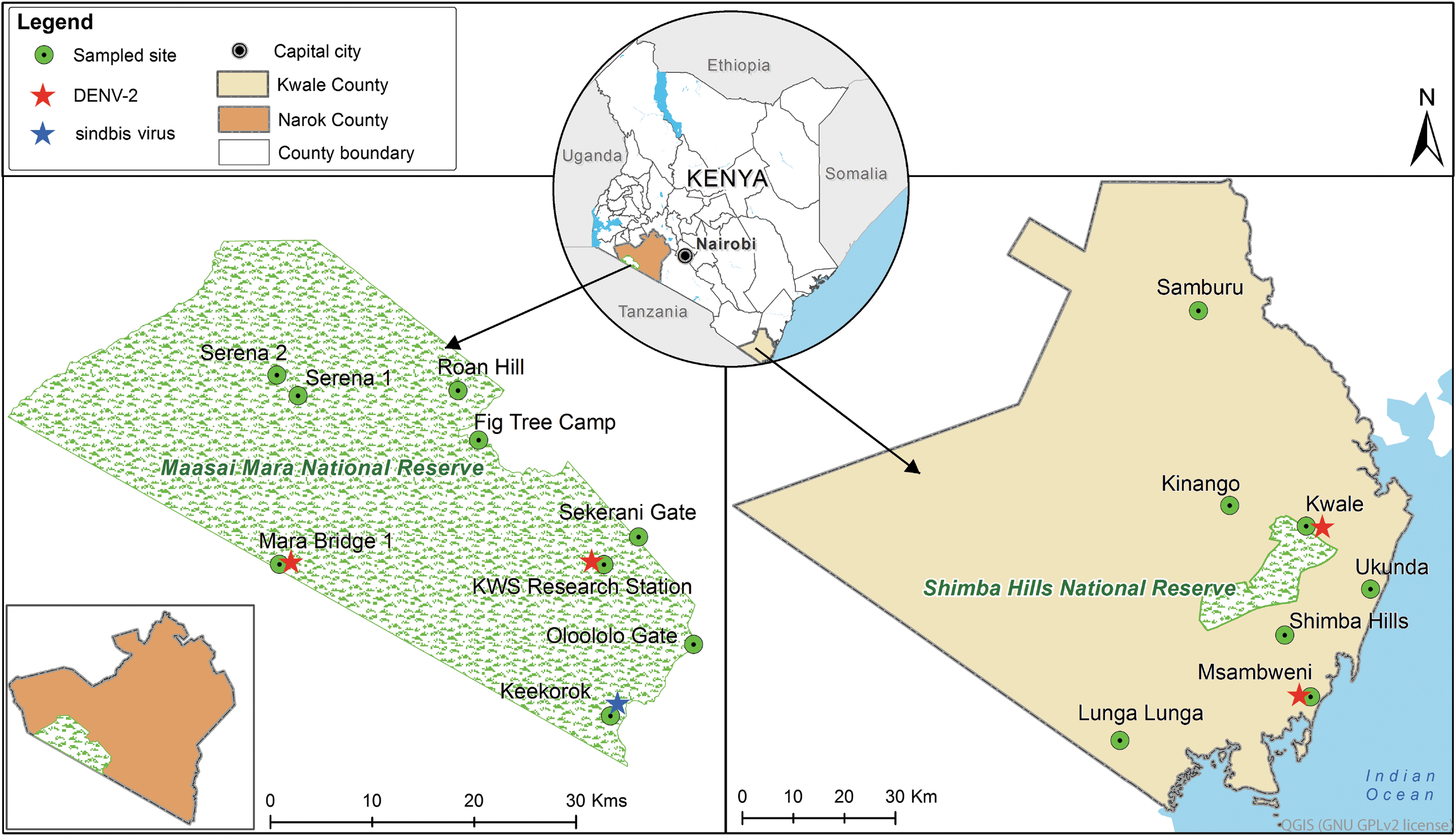
Map of mosquito sampling locations and sites of arbovirus detection in the MMNR in Narok County, Kenya, and near the SHNR in Kwale County, Kenya. The map was prepared using common-license shape files in QGIS software. MMNR, Maasai Mara National Reserve; SHNR, Shimba Hills National Reserve.
Adult mosquitoes were collected between January and June, 2018, using Centers for Disease Control and Prevention (CDC) light traps and Biogent (BG) sentinel traps (Biogents AG, Regensburg, Germany) baited with carbon dioxide (from dry ice). To utilize 24 traps effectively, we placed them randomly within the study sites, at least 50 m from each other. Near the SHNR, traps were placed at the periphery of villages near to the reserve's border and away from artificial lights. Both the CDC light traps and BG sentinel were suspended ∼2 m above the ground. The two types of traps were operated from 4 p.m. to 6:30 a.m. at dawn, and for four consecutive days. The sampled mosquitoes were immediately stored in liquid nitrogen and transported to the laboratory at icipe, Nairobi. Sampling was not done to compare the two study areas, but to have more data on novel arboviruses circulating in mosquito species found along the wildlife-human interfaces, to inform on possible disease outbreaks.
Morphological and molecular identification of mosquitoes
Using morphological identification keys (Edwards, 1941, Gillies and de Meillon, 1968, Gillies and Coetzee, 1987), we identified mosquitoes on Petri dishes placed on ice-chilled blocks under a stereomicroscope. We pooled up to 25 mos- quitoes into sterile 1.5-mL microcentrifuge tubes according to species, sex, site, date of collection, and blood-fed status, and stored them at −80°C until used for molecular tests. Blood-fed mosquitoes were placed individually into tubes with assigned “pool” numbers. Representative individual mosquitoes from both study sites were identified using molecular assays (Folmer et al., 1994). DNA was extracted from one leg of individual mosquitoes using the DNeasy Blood and Tissue Kit (Qiagen, Hilden, Germany) and mitochondrial cytochrome c oxidase subunit 1 (COI) genes were amplified using LCO1490 and HCO2198 primers (Table 1). The PCR conditions included a 15-min initial denaturation step at 95°C, followed by 40 cycles of 1 min at 95°C, 40 s at 50°C, and 2 min at 72°C, and a final 5-min elongation at 72°C.
Primers Used
cyt b, cytochrome b; COI, cytochrome c oxidase subunit 1; HRM, high-resolution melting; rRNA, ribosomal RNA.
Identification of blood meal sources
The individual abdomens of engorged mosquitoes were separated from the rest of the body using sterile dissecting pins and sharp-pointed forceps and placed into individual sterile 1.5-mL microcentrifuge tubes (Omondi et al., 2015). The other body parts of individual mosquitoes were stored in correspondingly labeled sterile 1.5-mL microcentrifuge tubes. Genomic DNA was extracted from the engorged abdomens using an in-house sodium dodecyl sulfate optimized method. Briefly, the abdomens of blood-fed mosquitoes were squashed in 300 μL cell lysis buffer using a handheld battery-operated homogenizer and then incubated at 65°C for 15 min. Thereafter, 100 μL of protein precipitation buffer was added, followed by vortexing for 30 s at room temperature before placing on ice for 5 min, and then centrifuging at 25,000 relative centrifugal force (rcf) for 5 min. The pellets were discarded, and the supernatants were placed in fresh tubes with 300 μL isopropanol, then mixed by inverting 100 times, and centrifuged at 25,000 rcf for 30 min. The supernatants were then removed and 300 μL of cold 70% ethanol was added to each tube. Tubes were inverted 50 times, and then centrifuged for 15 min. The ethanol was pipetted off and the tubes were inverted on paper towels overnight to air dry. The DNA pellets were then eluted with 50 μL of nuclease-free water and stored in the fridge at 4°C for 3 h before use.
Three sets of PCR primers (Table 1) targeting vertebrate COI, cytochrome b (cyt b), 16S ribosomal (r)RNA genes were used to identify blood meal sources in separate 10-μL PCR reactions containing 6 μL of nuclease-free PCR water, 0.5 μM concentration of each primer, 2 μL of 5 × Hot FIREPol EvaGreen HRM Mix (Solis Biodyne, Tartu, Estonia), and 1 μL of DNA as a template, using the same conditions as described by Omondi et al. (2015). Briefly, the thermal cycling conditions used for the cyt b primers were as follows: initial denaturation for 1 min at 95°C, followed by 35 cycles of denaturation at 95°C for 30 s, annealing at 58°C for 20 s, and extension at 72°C for 30 s, followed by a final extension at 72°C for 7 min. Cycling conditions for the 16S rDNA fragment were similar to those of cyt b, except for an annealing temperature of 56°C. The thermal cycling conditions for PCRs utilizing the COI primers included an initial denaturation at 95°C for 15 min, followed by 35 cycles of denaturation at 95°C for 30 s, annealing at 50°C for 30 s, and extension at 72°C for 60 s, followed by a final extension at 72°C for 7 min.
Normalized high-resolution melting (HRM) profiles were generated between ramping temperatures 75°C and 88°C, and compared to those from positive control DNA extracts from Homo sapiens (human), Felis catus (cat), Kobus ellipsiprymnus (waterbuck), Gallus gallus (chicken), Capra hircus (goat), Bos tarsus (cow), Sus scrofa (pig), Oryctolagus sp. (rabbit), Canis familiaris (dog), Nanger granti (gazelle), and Syncerus caffer (buffalo) tissues obtained from previous studies (Omondi et al., 2015, Ogola et al., 2017). Each set of PCRs included a sample of DNA extracted from male mosquitoes as a negative control. We sequenced representative samples to confirm blood meal sources identified by their HRM profiles.
Samples with HRM profiles that did not match any of the control HRM profiles were reamplified using COI barcoding primers (Table 1) in a SimpliAmp thermocycler (Thermo Fisher Scientific, MA). Thermal cyclic conditions included initial denaturation at 95°C for 15 min, followed by 40 cycles of 95°C for 20 s, 57°C for 30 s, and 72°C for 1 min, and a final extension at 72°C for 7 min.
Identification of arboviruses
Pools of 1–25 mosquitoes were homogenized in a Mini-BeadBeater-16 homogenizer (BioSpec, Bartlesville, OK). Remains of the blood-fed mosquitoes after removal of the engorged abdomens were individually homogenized. Briefly, the pools of the mosquitoes were added to microcentrifuge tubes containing 750 mg of 2-mm yttria-stabilized zirconium oxide beads (Glen Mills, Clifton, NJ), 150 mg of 0.1-mm zirconia/yttria beads, and 450 μL of phosphate-buffered saline. The supernatants obtained were loaded into 96-well plates for viral RNA extraction.
Viral RNA was extracted using the MagNA 96 Pure DNA and Viral NA Small Volume Kit (Roche Applied Science, Penzberg, Germany) in a MagNA Pure 96 (Roche Applied Science) automated extractor. RNA extracts were immediately reverse transcribed to cDNA using 10 μL of the viral RNA as a template in 20-μL reactions using the High-Capacity cDNA Synthesis Kit (Life Technologies, Carlsbad, CA). Nonribosomal hexamers (0.6 mM) were used during the reverse transcription (Endoh et al., 2005). We screened for flaviviruses and alphaviruses by PCR-HRM using cycling conditions as published by Villinger et al. (2017) in a Rotor-Gene Q HRM capable thermocycler (Qiagen, Redwood City, CA).
Flaviviruses were screened using the Flavi-JV2a-F and Flavi-JV2a-R primers and alphaviruses were screened using the Vir-2052-F and Vir2052-R primers (Table 1) in 10-μL PCR-HRM assays using 2 μL cDNA template, 5 μL MyTaq HS master mix (Bioline Reagents Limited, London, UK), 1 μL of 50 μM SYTO-9 saturating intercalating dye (Life technologies), 1 μL of the primer mix, and 1 μL of nuclease-free water. Dengue virus (DENV), sindbis virus (SINV), chikungunya virus, and Semliki Forest virus culture stocks were used as positive controls and nuclease-free water was used as a negative control (Villinger et al., 2017). To further characterize alphaviruses identified by PCR-HRM, cDNA of Alphavirus positive samples was re-amplified using Alpha-M2W and Alpha-cM3W primers. These PCR products were diluted 10-fold and 1-μL volumes of these PCR product dilutions were used as templates in hemi-nested PCRs using the same reverse primer (Alpha-cM3W) with a nested forward primer (Alpha-M2W2) (Table 1) in a SimpliAmp thermocycler (Thermo Fisher Scientific). We determined the minimum infection rates (MIRs; number of positive pools/total specimens tested × 1000) of specific arboviruses detected in either sex of specific mosquito species sampled in either the MMNR or from locations near the SHNR.
Sequencing and phylogenetic analyses
Mosquito, vertebrate blood meal, and arbovirus PCR products were electrophoresed alongside 1 kb DNA ladders (Solis Biodyne) for 31 min at 300 A/125 V in 1.5% ethidium bromide-stained agarose gel in Tris-borate ethylenediaminetetraacetic acid (EDTA) buffer. DNA bands of interest were excised from agarose gels with clean, sharp scalpels over a blue-light transilluminator for visualization and purified using GenScript QuickClean II Gel Extraction Kit (GenScript) following the manufacturer's protocol. The blood meal 16S rRNA and cyt b HRM amplicons were purified using the ExoSAP-IT kit (USB Corporation, Cleveland, OH). The purified amplicons were then sent for sequencing in both directions at Macrogen (Netherlands).
All nucleotide sequences were edited and aligned using the MAFFT plugin in Geneious software version 8.1.9 (created by Biomatters) (Kearse et al., 2012). Closely related sequences were identified by querying the GenBank nucleotide database using the Basic Local Alignment Search Tool (
Results
Mosquito diversity and abundance
From the MMNR, 722 mosquitoes, comprising 20 species from five genera, were sampled (Table 2). Culex was the most abundant genus, with Culex pipiens being the most abundant species (35.8%). The Culex genus also had the greatest diversity with nine species identified, followed by five Aedes spp. (GenBank acc. nos. MK533632–MK533636) and four Anopheles spp. (Table 2, Fig. 2). Among the Anopheles, two Anopheles specimens (GenBank acc. nos. MK533642–MK533643) with sequences clustering among Anopheles coustani complex sequences (Fig. 2) were obtained. We also sampled two Eretmapodites chrysogaster mosquitoes (GenBank acc. nos. MK533645 and MK510878) and a single Aedes metallicus mosquito (GenBank acc. no. MK533635).
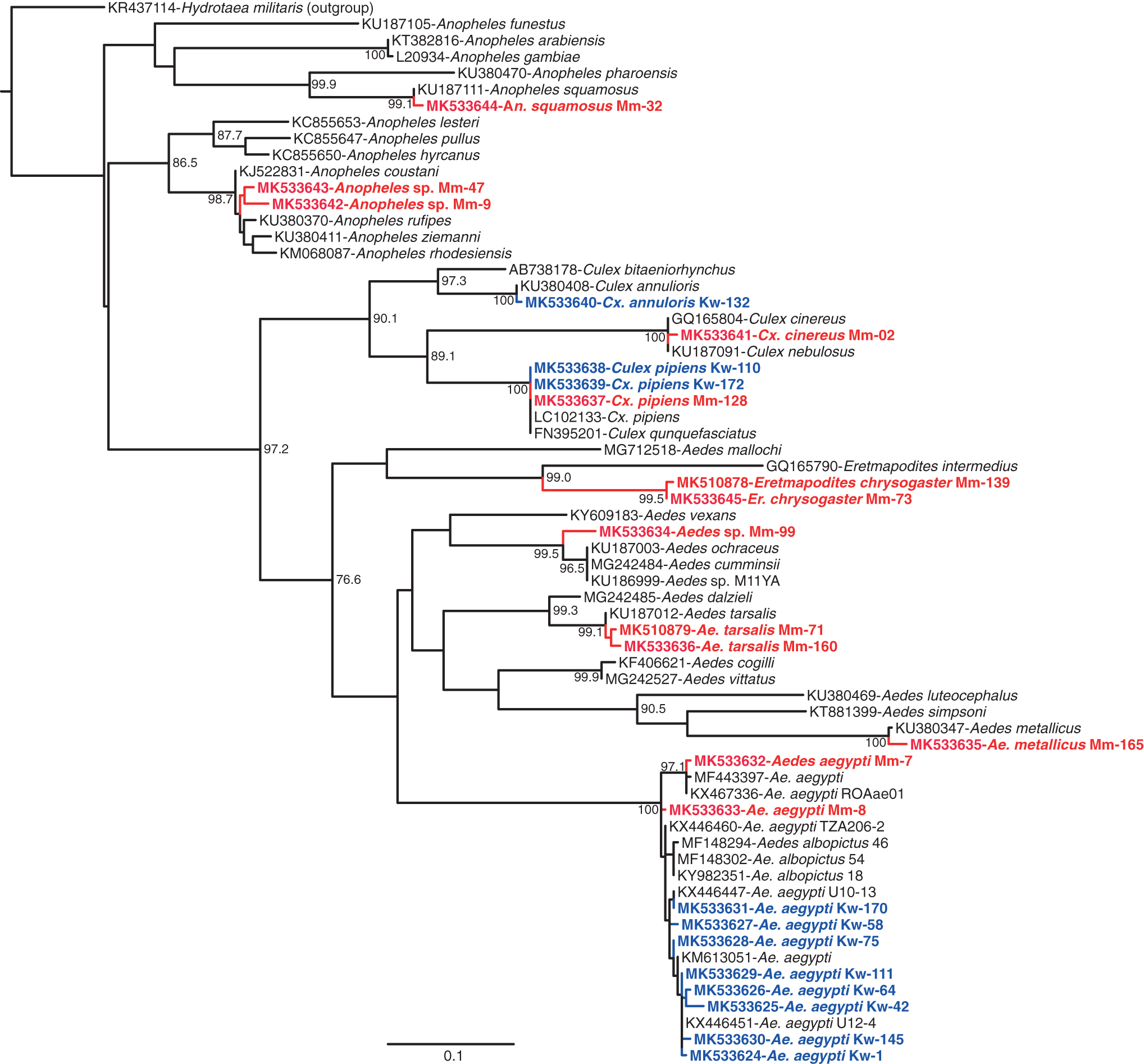
Maximum likelihood phylogenetic tree of 658-nucleotide mosquito COI sequences. GenBank accessions, species, and isolates are indicated. The sequences obtained in this study are indicated in red (from the MMNR) and in blue (from locations near the SHNR). Hydrotaea militaris was used as an outgroup. Bootstrap values at the major nodes are of percentage agreement among 1000 replicates. The branch length scale represents substitutions per site. COI, cytochrome c oxidase subunit 1.
Mosquito Abundance and Diversity in the MMNR and Near the SHNR
MMNR, Maasai Mara National Reserve; SHNR, Shimba Hills National Reserve.
From locations near the SHNR, 2136 mosquitoes were sampled with Cx. pipiens being the most abundant species (61.99%). A single specimen of Culex annuloris (GenBank acc. no. MK533640) was sampled. Phylogenetic analysis revealed that the Ae. aegypti COI gene sequences (GenBank acc. nos. MK533624–MK533631) are highly polymorphic (Fig. 2). Overall, sequencing of COI genes confirmed the identity of 14 out of 15 mosquito species and resolved one species, Cx. annuloris (GenBank acc. no. MK533640), which was morphologically misidentified as Culex ethiopicus.
Arboviruses identified
Out of 170 mosquito pools from the MMNR, three female pools—Aedes tarsalis (MIR = 250.0), Aedes tricholabis (MIR = 58.8), and Ae. aegypti (MIR = 333.3)—were positive for DENV serotype-2 (DENV-2) (Fig. 3a; Supplementary Fig. S1) and one male Cx. pipiens (GenBank acc. no. MK533637) pool was positive for Sindbis virus (SINV) (GenBank acc. no. MK510862) (MIR = 27.8) (Figs. 3b and 4). From locations near the SHNR, out of 216 mosquito pools, 2 (0.9%) female Ae. aegypti pools were positive for DENV-2 (MIR = 12.7) (Table 3).
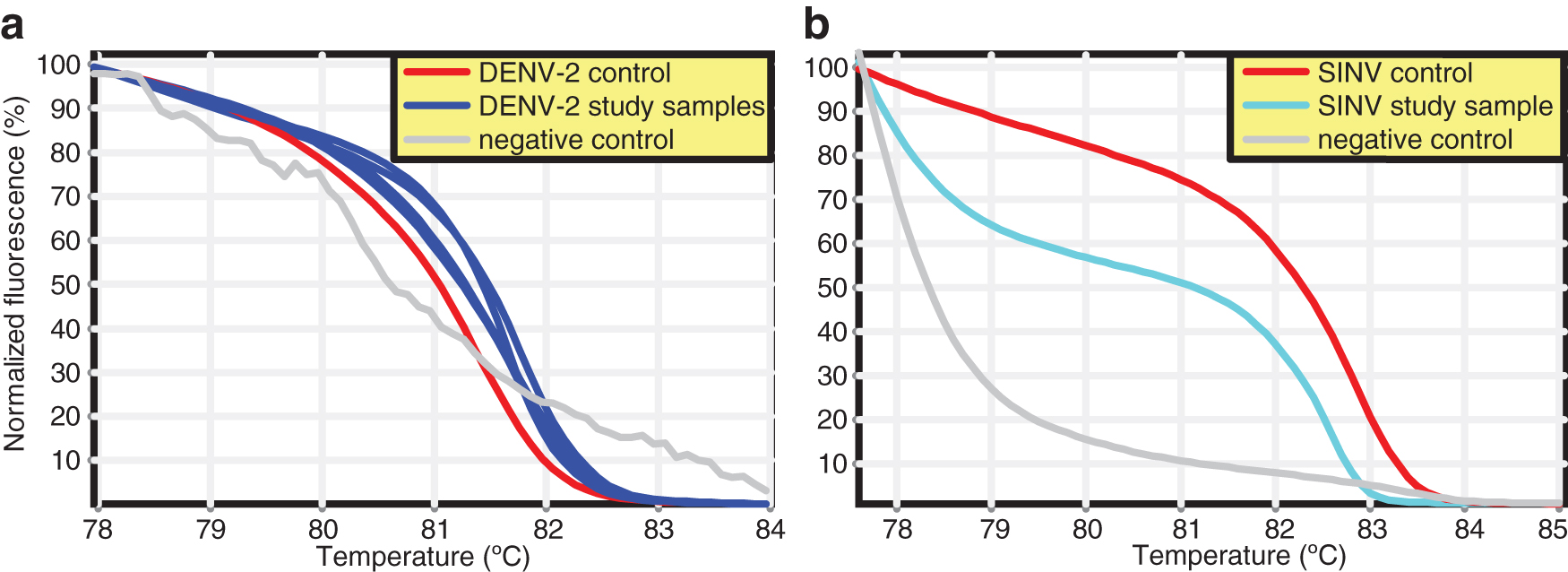
Normalized HRM profiles of RT-PCR of
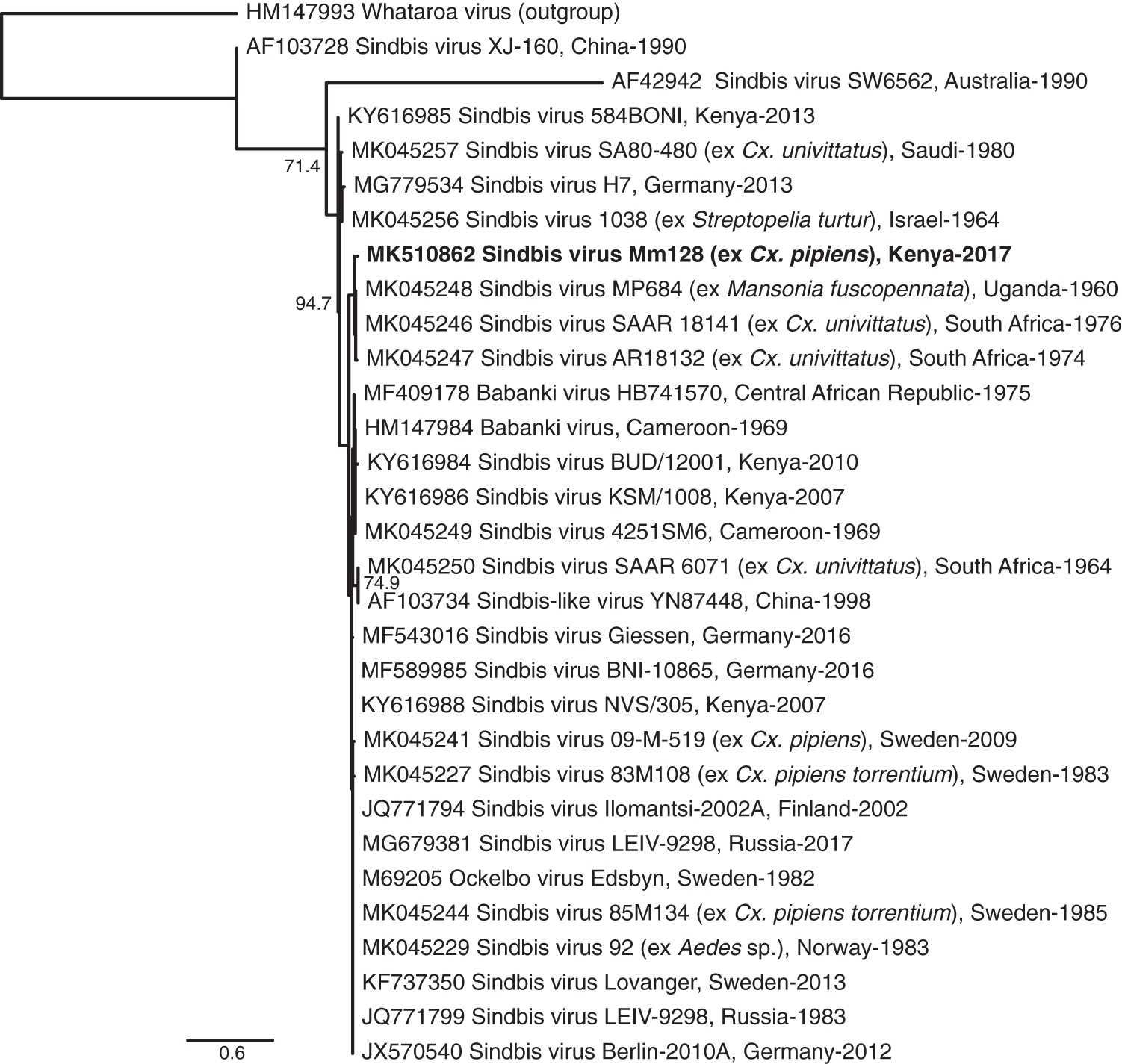
Maximum likelihood phylogenetic tree of 254-nucleotide SINV sequences. GenBank accessions, strain names, country, and year of isolates are indicated. The sequence obtained in this study is red. Whataroa virus was used as an outgroup. Bootstrap values at the major nodes are of percentage agreement among 1000 replicates. The branch length scale represents substitutions per site.
Arboviruses Identified
MIR, minimum infection rate.
Blood meal analysis
Out of the 722 mosquitoes collected in the MMNR, 3 (0.4%) were blood engorged (Table 4). One Er. chrysogaster (GenBank acc. no. MK533645) had fed on both human (cyt b GenBank acc. no. MK510869) and hippopotamus (cyt b GenBank acc. no. MK510870) blood (Supplementary Fig. S2) and a Culex zombaensis had fed on human blood (cyt b GenBank acc. no. MK510866) (Fig. 5a). The DENV-2-positive Ae. tarsalis mosquito had a blood meal from a bushbuck (COI GenBank acc. no. MK510865; cyt b GenBank acc. no. MK510872), although we cannot conclude that this was the source of infection.
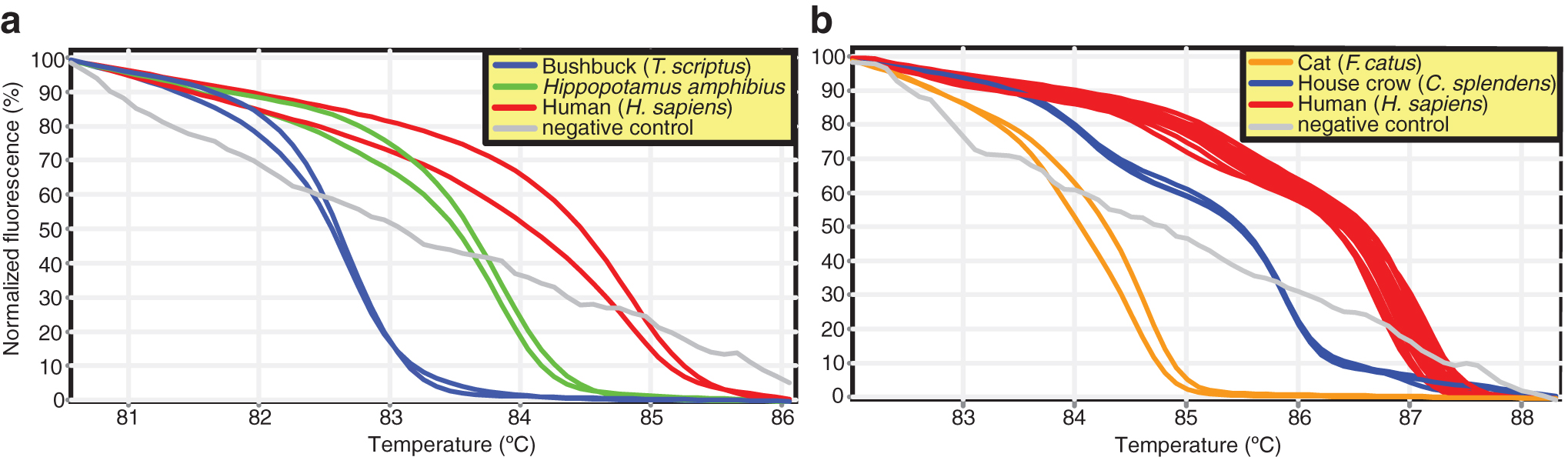
Normalized HRM profiles represented as percent fluorescence of mosquito blood meal sources using
Blood-Fed Mosquitoes and Their Blood Meal Sources in the MMNR and Near the SHNR
Mixed blood meal.
Of 2136 mosquitoes sampled from locations near the SHNR, 48 (2.25%) were blood fed (Table 4). The blood meal sources included humans (COI GenBank acc. nos. MK510863–MK510864), domestic cat (cyt b GenBank acc. no. MK510873), house crow (Corvus splendens; cyt b GenBank acc. nos. MK510874–MK510876), house sparrow (Passer domesticus; Supplementary Fig. S3), chicken (cyt b GenBank acc. no. MK510871), and dog (Canis lupus; cyt b GenBank acc. no. MK510877) (Table 4; Fig. 5b).
Discussion
The arbovirus infections identified in zoophilic mosquitoes from the predominantly wildlife habitats of the MMNR (DENV-2 & SINV) and locations near the SHNR (DENV-2) highlight the importance of their sylvatic transmission in these ecologies, with their potential to also infect humans, as mosquitoes fed both on humans and wildlife. Many of the Aedes, Culex, Anopheles, and Mansonia mosquitoes identified are well known for their role in the transmission of arboviruses in Kenya (Ochieng et al., 2013).
However, to our knowledge, this is the first time DENV has been detected in Ae. tarsalis and Ae. tricholabis. Nonetheless, Ndumu virus (Alphavirus) has been isolated from Ae. tricholabis from North Eastern Province, Kenya (Ochieng et al., 2013), and the orthobunyaviruses, Pongola virus (Mossel et al., 2017), and Beliefe virus (Metselaar et al., 1974) have been isolated from Ae. tarsalis in Uganda. We also detected DENV-2 RNA sequences in Ae. aegypti, the known principal vector of dengue, Zika, chikungunya, and yellow fever viruses to humans (Patterson et al., 2016). The detection of DENV-2 in sympatric Ae. tarsalis and Ae. tricholabis gives insight into the possibility of sylvatic vectors involved in the transmission of dengue that could spill over to urban to human settlements around wildlife reserves leading to outbreaks. We were not able to determine whether the observed DENV-2 sequences in mosquitoes stemmed from established infections in the mosquito or from infected blood meals. Further studies are needed to determine the vector competency of Ae. tarsalis and Ae. tricholabis mosquitoes to transmit DENV-2. Whether DENV-2 viruses in the region stem from sylvatic strains endemic to East Africa, or from more recent introductions from southeast Asia, also requires further investigation (Weaver and Vasilakis, 2009).
As male mosquitoes do not feed on blood, the SINV infection in one pool of male Cx. pipiens in the MMNR is likely due to vertical or environmental transmission. SINV has been isolated from Culex and Aedes mosquito species in Kenya (Sigei et al., 2018) and has been found in a Cx. pipiens mosquito that had fed on human blood in Baringo County, Kenya (Omondi et al., 2015). The SINV sequence identified in this study clustered most closely with recently deposited sequences of historical isolates from Uganda and South Africa isolated from Mansonia fuscopennata and Culex univittatus mosquitoes, respectively (Ling et al., 2019) (Fig. 4). This suggests that the SINV circulation is relatively stable and endemic in African wildlife and zoophilic mosquito populations with potential risk of future disease outbreaks. Birds are thought to serve as the main reservoir hosts (Hesson et al., 2016) with migratory birds having been associated with the spread of SINV across the continents (Hubálek, 2004). In Kenya, migratory birds converge along the coastal beaches, reefs, mangroves trees, and lakes in the Rift Valley during their season migrations (Bennun et al., 2000).
Most blood meals identified from both study areas were from humans. Although the MMNR is comparatively less encroached by humans than the SHNR, two out of three MMNR blood-fed mosquitoes identified had blood from humans. This could be because the traps were set in areas with human activities in the reserve. Er. chrysogaster had a mixed blood meal from human and hippopotamus in the MMNR. This opportunistic blood-feeding behavior (multiple-host feeding) is typically demonstrated by mosquito vectors when suitable vertebrate hosts are unavailable (Lardeux et al., 2007) and is of medical importance since the mosquito could act as bridge vectors for arbovirus transmission between wildlife animals and humans.
Aedes aegypti and Cx. pipiens in locations near the SHNR demonstrated a high hematophagic tendency toward humans. Our finding that Cx. pipiens had many blood meals from humans concurs with those of Omondi et al. (2015), However, house crow, house sparrow, chicken, and dog were also identified as Cx. pipiens blood meal sources. This feeding pattern makes Cx. pipiens a potential vector for transmitting arboviruses from avian hosts to humans. Although most Ae. aegypti mosquitoes had fed on humans, the DENV-2 positive Ae. tarsalis mosquito sampled in the MMNR had a blood meal from a bushbuck. However, we cannot directly incriminate bushbuck as a reservoir for DENV-2. More serological studies need to be done from these wildlife species for reactivity against Flavivirus antigens.
Conclusions
Although we were not able to determine whether the zoophilic mosquitoes obtained the DENV-2 and SINV infections from wildlife, these findings highlight the potential importance of its sylvatic transmission among wildlife and zoophilic mosquitoes in maintaining the virus in enzootic circulation, which may be playing an important role in sporadic dengue outbreaks in Kenya. Eastwood et al. (2017) found evidence of DENV in olive baboons that were closer to human habitation in south coast, but not those baboons from the interior of SHNR. This suggests that spill-back from humans into wildlife may equally be occurring. Nonetheless, this study highlights that DENV-2 is circulating in an East African wildlife ecology with Ae. tarsalis and Ae. tricholabis as possible vectors and that SINV may be maintained in zoophilic mosquitoes by vertical or environmental transmission.
Footnotes
Acknowledgments
We acknowledge Enock Mararo (icipe) for technical laboratory support and Jackson Kimani for help in preparing the map figure. We are also thankful to Edwin Ogola for provision of blood meal and arbovirus-positive controls used in the study. We would like to thank Francis Maluki Mutuku and Peter Siema Musunzaji of Msambweni Division of Vector Borne Diseases (DVBD) for providing the traps and helping in mosquito collection in locations near the SHNR.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
We gratefully acknowledge the financial support for this research by the following organizations and agencies: United States Agency for International Development (USAID) and Partnerships for Enhanced Engagement in Research (USAID-PEER) cycle 4 awarded to L.W., under the USAID grant no. AID-OAA-A-11-00012 subawarded by the American National Academy of Sciences (NAS) under agreement no. 2000006204. Additional support was obtained from icipe institutional funding from the UK's Department for International Development (DFID), the Swedish International Development Cooperation Agency (SIDA), the Swiss Agency for Development and Cooperation (SDC), and the Kenyan Government.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.