
Abstract
Background:
Aedes albopictus is a major vector for transmission of many viral pathogens. Deltamethrin resistance was analyzed by catching A. albopictus in the field; the analysis effect is affected by many insecticides that often interact with A. albopictus in the field environment.
Materials and Methods:
This study examined the development of deltamethrin resistance in A. albopictus mosquitoes under controlled laboratory conditions, focusing on morphological changes, reproductive fitness, and mutation of the knockdown resistance (kdr) gene. Deltamethrin-resistant strains were selected up to the 20th generation. To determine the level of resistance, the lethal concentration 50 (LC50) of deltamethrin in the larvae was obtained, followed by the resistance ratio (RR), and in adult mosquitoes, mortality rates were calculated using the contact tube method.
Results:
An increase in the LC50 from 0.0070 to 0.0563 mg/L was observed in resistant versus sensitive strains, with an increase of 11.26 in the RR. Overall, the results of the larval resistance bioassay showed that resistant larvae had medium resistance; however, by the 20th generation, adult mosquitoes showed strong resistance. PCR amplification, cloning, and sequencing of sodium channel domain III gene fragments were subsequently carried out using selected resistant and sensitive female mosquitoes. As a result, a number of base mutations were observed in the kdr gene in the resistant strain; however, no amino acid sequence mutations were observed, suggesting that base sequence changes did not affect protein expression. Results of morphological changes between resistant and sensitive strains showed that significant differences in the body, foreleg, mid leg, and hind leg length, as well as wing length and width, antenna length, and proboscis length were observed between 18th-generation resistant and sensitive strains of A. albopictus. On analysis of reproductive fitness associated with deltamethrin resistance in selection of mosquitoes, observation results showed differences between resistant and sensitive strains; the female/male ratio of mosquitoes decreased after pupa hatching, with more females and fewer males.
Conclusions:
The model of deltamethrin-resistant selection of A. albopictus was successfully established in the laboratory. The morphological phenotypes of the deltamethrin-resistant population of A. albopictus mosquitoes had changed. The kdr gene of the 19th and 20th generations of deltamethrin-resistant A. albopictus mosquitoes had silent mutations at several sites. After deltamethrin resistance selection, the female/male ratio of mosquitoes increased after pupa hatching, with more females and fewer males, hinting at increased chances of more female mosquitoes transmitting diseases.
Introduction
Aedes albopictus originated in Southeast Asia, but has now spread to more than 70 countries worldwide, spanning tropical, subtropical, and temperate zones. As a result, it has become one of the 100 fastest spreading species in the world in the past 20 years and one of 100 malignant species listed by the Invasive Species Specialist Group (Yang et al. 2013). It is known as the Asian tiger mosquito because of the aggressive nature of the adults, and the eggs show excellent drought and cold resistance. A. albopictus is an important virus vector, transmitting a number of diseases such as dengue fever, chikungunya fever, and yellow fever (Liu et al. 2018). As a result of its high survival ability, A. albopictus has therefore become a major global health concern.
Deltamethrin is a highly efficient, broad-spectrum pyrethroid insecticide that has been widely used in agriculture and general pest control since the 1970s due to its long duration of efficacy, good level of safety, and overall stability. Pyrethroids are intensively used to control Aedes mosquitoes and dengue transmission (Marcombe et al. 2011); however, in many areas, A. albopictus has reportedly developed low to moderate deltamethrin resistance (Pang et al. 2018, Peng et al. 2018, Xing et al. 2018). Understanding the occurrence and development as well as the mechanism of deltamethrin resistance in A. albopictus is therefore an important subject of vector resistance research.
The two major causes of insecticide resistance are alterations in the target sites and increase in the rate of insecticide metabolism (Zheng and Hu 2006). While metabolic resistance is caused primarily by three enzyme families, cytochrome P450s, esterases, and glutathione S transferases, target site resistance is conferred by one or several mutations in the insecticide target site (Zheng and Hu 2006). Pyrethroids target the VGSC gene, also known as the voltage-gated sodium channel (VGSC) of insect neurons (Martinez-Torres et al. 1998, Hu et al. 2011, Kasai et al. 2011). The other known mechanism is reduced target site sensitivity resulting from nonsynonymous mutations in the VGSC gene, leading to single amino acid substitutions, which has been shown to be correlated with phenotypic resistance to pyrethroids (Zhong et al. 2013). Zhou et al. (2019) reported on knockdown resistance (kdr) mutations within 17 field populations of A. albopictus from Beijing, China: the first report of a novel V1016G mutation and evolutionary origins of kdr haplotypes (Zhou et al. 2019). Gao et al. (2018) reported on correlation between adult pyrethroid resistance and kdr mutations in A. albopictus (Diptera: Culicidae) field populations in China. However, deltamethrin resistance was analyzed by catching A. albopictus in the field, and the analysis effect is affected by many insecticides that interact with A. albopictus in the field environment. Thus, by studying deltamethrin resistance in the laboratory using a single deltamethrin-resistant (DLR) mosquito strain with a clear genetic background would aid our understanding of the molecular mechanism of resistance.
In this study, we therefore used a single deltamethrin insecticide to select resistance in 20 generations of A. albopictus mosquitoes under controlled laboratory conditions. The mosquito's clones carry a clear genetic background; its genome has been sequenced (the sequence's accepted number is NCBI IDJXUM0000000). To determine the level of resistance, morphological changes and reproductive fitness were observed along with the VGSC gene sequence and corresponding protein sequence analysis. Our findings provide a model for further molecular and proteomic studies on the causes and mechanisms of deltamethrin resistance in A. albopictus.
Materials and Methods
Ethics approval and consent to participate
Mice were used for feeding the mosquitoes in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health and guidelines of Southern Medical University on experimental use of mice. All of the animals were handled according to approved Institutional Animal Care and Use Committee (IACUC) protocols (#2017-005) of Southern Medical University.
Mosquito strain
The A. albopictus Foshan strain was bred in the Pathogenic Biology Laboratory of the School of Public Health, Southern Medical University, Guangdong Province, China. The Foshan strain of A. albopictus was obtained from the Center for Disease Control and Prevention of Guangdong Province, China, where it has been in culture since 1981. Mosquitoes were reared at 28°C and 70–80% relative humidity under 14-h light/10-h dark cycles. The larvae were fed with Inch Gold® turtle food every day at an average of about 0.1 g per 100 larvae per day. Adults were kept in 30-cm3 cages. Mosquitoes were provided with 10% glucose daily, and every 3 days, a mouse was placed in each cage for ∼4 h to blood-feed the mosquitoes. It had been confirmed as a sensitive strain (S-lab strain).
Pesticides and reagents
Deltamethrin, 98.37% purity, technical-grade pyrethroid chemicals were provided by Jiangsu Yangnong Chemical Group Co., Ltd., in China. Acetone (99.5% purity) and glucose were purchased from Guangzhou Chemical Reagent Factory (China Production). Contact canister and 0.05% deltamethrin (98.37% purity, 0.0178 g/m2) film were also purchased from the China Center for Disease Control and Prevention. Specificity of the contact canister and 0.05% deltamethrin is as follows: the mortality rate is more than 98% in the sensitive group (s), 80–97% in the primary group step resistant population (m), and <80% in the resistant population (R).
The Insect DNA Kit and Gel Extraction Kit were purchased from Omega Bio-Tek Company (USA), and the PMD18-T Vector Kit was purchased from TaKaRa Company (Japan).
Analysis of deltamethrin resistance in larvae of A. albopictus
Larval susceptibility testing was performed according to WHO guidelines (WHO 2011–2013). For each bioassay, 25 late third or early fourth instar larvae from each population were transferred to plastic cups with 99 mL of distilled water and 1 mL of deltamethrin at the desired concentration, diluted in ethanol, leading to 0–100% mortality.
An appropriate volume of dilution from the stock solution was added to the water in the cups to obtain the target dose, diluted in acetone, starting with the lowest concentration. Four replicates of 10 different concentrations between 0.004 and 0.15 ppm and acetone only as control were tested on 25 early fourth instar larvae.
The mortality rate was recorded after 24 h of exposure. Larvae that were unable to swim up to the surface were counted as dead, and the larvae that pupated were omitted from the final total. The lethal concentration that kills 50% of the tested samples (LC50) was calculated using probit analysis (SPSS Statistics 22 software).
Large-sample screening of resistance was subsequently carried out using ∼1500 4th instar larvae soaked in deltamethrin–acetone solution containing dechlorinated water overnight. The LC50 concentration was determined based on the above-mentioned small sample and each of the bioassay test results. Deltamethrin of LC50 was added to the water for population selection. The larvae were placed in a stainless steel dish with 5 L of dechlorinated water overnight (mean 16 h). After 24 h, the surviving larvae were selected and fed to adult mosquitoes. Each bioassay was conducted at 28°C and 70–80% relative humidity under 14-h light/10-h dark cycles. The larvae were fed with Inch Gold turtle food every day at an average of about 0.1 g per 100 larvae per day. Some of the selected generations were recorded as rate of development female and male from S1-R20 generations. Adult mosquitoes in 3–4 days after eclosion suck on Kunming rat blood. Three days after feeding blood, they were put into wet filter paper for laying eggs. The egg paper was dried in the mosquito room, marked with the date and algebra, placed for one week for the egg to mature, and then placed in dechlorinated water for incubation. The above steps were repeated to select mosquitoes up to the 20th generation.
The resistance ratio (RR) was then determined using the LC50 of each generation with that of the sensitive 1st-generation LC50. Those with an RR ≤2 were deemed sensitive (−), >2 ≤ 10 was deemed low resistance (+), >10 ≤ 20 was deemed medium resistance (++), and >20 was deemed high resistance (+++) (WHO 2011–2013, Xu et al. 2016, Xie et al. 2018).
Analysis of deltamethrin resistance in adult A. albopictus
Adult bioassays were carried out according to the WHO protocol (WHO 2013). Female mosquitoes that were 3–5 days old after emergence and not blood fed were tested for susceptibility to deltamethrin insecticides by the tube bioassay. The concentration of insecticide-impregnated test papers of deltamethrin is 0.05%. A. albopictus mosquitoes with 4 replicates of 25 mosquitoes per tube were used for the bioassay. Control tests were performed using silicone oil preimpregnated papers. The number of mosquitoes being knocked down was recorded every 10 min during the 60-min exposure period. Mortality was scored after a 24-h recovery period.
If the mortality rate of the control group was >20%, the experimental data were considered invalid, but if it was <5%, the mortality rate of the experimental group was subsequently calculated. If the mortality rate was between 5% and 20%, the corrected mortality rate was obtained. The corrected mortality rate was determined using Abbott's formula (WHO 2011–2013, 2016) as follows:
Mosquitoes with a mortality rate >98% were deemed sensitive (S), those with a mortality rate of 90–98% were deemed initially resistant (M), and those with a mortality rate <90% were deemed resistant (R).
Analysis of morphological changes in deltamethrin-resistant A. albopictus
Quantitative comparisons of morphological indices in sensitive and resistant adult A. albopictus were subsequently carried out in each genetic generation. Thirty females were selected from the resistant group and the sensitive group, respectively. The mosquitoes were female mosquitoes, 3–5 days old after emergence, and not blood fed. Measurements were made using a micrometer under a microscope. Indexes were as follows: sex, body, foreleg, mid leg, and hind leg length, wing length and width, antenna length, and proboscis length.
Analysis of reproductive fitness in deltamethrin-resistant A. albopictus
Four batches of 25 mosquitoes that were 1–2 days old after emergence (5 males and 20 females, another group of 20 males and 5 females, each group had 3 repeats) and deltamethrin sensitive (SS) and DLR (RR) were released, respectively, in each cage consisting of an 85-oz popcorn bucket (17.8 cm caliber, 14.5 cm bottom diameter, and 14.5 cm height).
Three days later, emerging adults were provided with a bloodmeal marking the beginning of the first gonotrophic cycle. After the first round of egg-laying, remaining females were counted, then after 24 h of starvation, a second bloodmeal was provided. The period from the beginning of one bloodmeal to the next was recorded as one gonotrophic cycle. To determine fecundity, eggs were collected and counted daily using a filter paper in a small cup kept in the cage. A fresh filter paper in a small cup was placed in the cage daily until no further eggs were laid for at least 48 h. The number of eggs was counted using a dissecting microscope at a magnification of × 40. Eggs counted after each daily bloodmeal were documented separately. During each gonotrophic cycle, the eggs on the filter paper were submerged in a tray filled with dechlorinated water and allowed to hatch. The hatched larvae were then counted, and the rate of hatching was calculated by determining the ratio of hatched larvae to the number of eggs laid. The pupation rate was also determined at the same time. The conditions for mosquito culture are the same as the above. Fecundity of the SS strain was then compared with that of the RR strain, and the fecundity of each strain during three cycles was also compared. The results were analyzed using the χ 2 test with statistical significance set at p ≤ 0.05. All analyses were conducted using SPSS 22.0 statistical software.
Analysis of the sodium channel gene (VGSC) in deltamethrin-resistant A. albopictus
The domain III (kdr) gene fragment of the VGSC gene was amplified by PCR using the following primers: aegSCF7 (gagaactcgccgatgaactt) and aegSCR7 (gacgacgaaatcgaacaggt) (Chen et al. 2016). The PCR mixture consisted of 1.00 μL of DNA template, 22.00 μL of GoTaq® Green Master Mix, 1 μL of each primer (F and R), and 9.50 μL of high-pressure sterilization ddH2O, adjusted to a final volume of 25.00 μL. The PCR conditions were as follows: 95°C for 5 min, followed by 30 cycles of amplification at 94°C for 30 s, 57°C for 45 s, and 72°C for 60 s, with a final extension step at 72°C for 8 min. PCR products were obtained by agarose gel electrophoresis, purified (Invitrogen DNA Purification Kit), and recovered, then linked to the pMD18-T vector (Guangzhou Ruizhen Biotechnology Co., Ltd.) before transformation into Escherichia coli DH5α competent cells. The positive plasmid was then screened using the colony PCR method, and positive plasmids were extracted and identified by double restriction enzyme digestion and sequenced by the biotechnology company, Shanghai Sangon Biotech Company.
Two generations of the sensitive and resistant A. Albopictus strains, the 19th and 20th generations, respectively (S-19 and S-20, R-19 and R-20), were subsequently tested for gene mutations. Five mosquitoes were selected as samples in each generation. DNA was extracted from a single mosquito (OMEGA Insect DNA Kit) and a single mosquito's DNA was used as the PCR template to amplify the VGSC domain III sequence in the target gene fragments. Each mosquito was sequenced at least three times (three different clones were selected and sequenced). Sequences of target gene fragments in resistant strains selected in the laboratory and sensitive strains reared in the laboratory were then compared using DNAMAN, BioEdit, and Clustal Omega software (Xu et al. 2016, Pwalia et al. 2019).
Statistical analysis
LC50 was calculated using the probit regression analysis and χ 2 test with SPSS 20.0 statistical software (Xu et al. 2016, Pwalia et al. 2019). DNAMAN, BioEdit, and Clustal Omega software were used to compare the sodium channel domain III bases and amino acid sequences and confirm any mutation sites.
Results
Resistance of A. albopictus larvae to deltamethrin
After resistance pressure screening, the LC50 increased from 0.0070 mg/L in S-1, 95% CI with 0.0063–0.0084, to 0.0563 mg/L in larvae from the resistant strain R-20, 95% CI with 0.0534–0.0605, resulting in an RR of 11.26. After 20 generations of screening, the resistance level LC50 in A. albopictus larvae to deltamethrin reached 0.0563 mg/L from 0.007mg/L and the RR was 11.26 (Table 1). The overall trend of the resistance level of A. albopictus to deltamethrin increased with time. The resistance level was stable after the ninth generation.
LC50 in Aedes albopictus Larvae of Deltamethrin (mg/L)
CI, confidence interval; RR, resistance ratio.
Resistance of adult A. albopictus to deltamethrin
Following the standard WHO guidelines, mosquitoes with a mortality rate >98% were deemed sensitive (S), those with a mortality rate of 90–98% were deemed initially resistant (M), and those with a mortality rate <90% were deemed resistant (R). Using the contact test with membrane filter paper, the mortality rate of the sensitive control group was calculated as 8%. After correcting this value with the mortality correction formula recommended by the WHO (see the Materials and Methods section), the mortality rate of the resistant 20th generation was calculated as 81.74%.
Morphological changes in deltamethrin-resistant A. albopictus
Significant differences in body, foreleg, mid leg, and hind leg length as well as wing length and width, antenna length, and proboscis length were observed between 18th-generation resistant and sensitive strains of A. albopictus (R-18 and S-18) (p < 0.05; Table 2 and Fig. 1). Average values of all eight indexes were also significantly different between the 1st- and 18th-generation resistant groups (R-1 and R-18) (p < 0.05; Fig. 2). The standard error of the morphological data of resistant strains in these two generations was less than 1.00, suggesting that the data were representative of and accurately reflected the morphological characteristics of these two groups. The morphological data were also compared between 1st- and 3rd-generation resistant strains (R-1 and R-3), revealing no significant differences in any of the eight indexes (p > 0.05; Fig. 3). In contrast, significant differences were observed between R-3 and R-18, except for the proboscis length (p < 0.05; Table 2 and Fig. 4). Overall, average values of all morphological indexes showed a decreasing trend (Fig. 5).
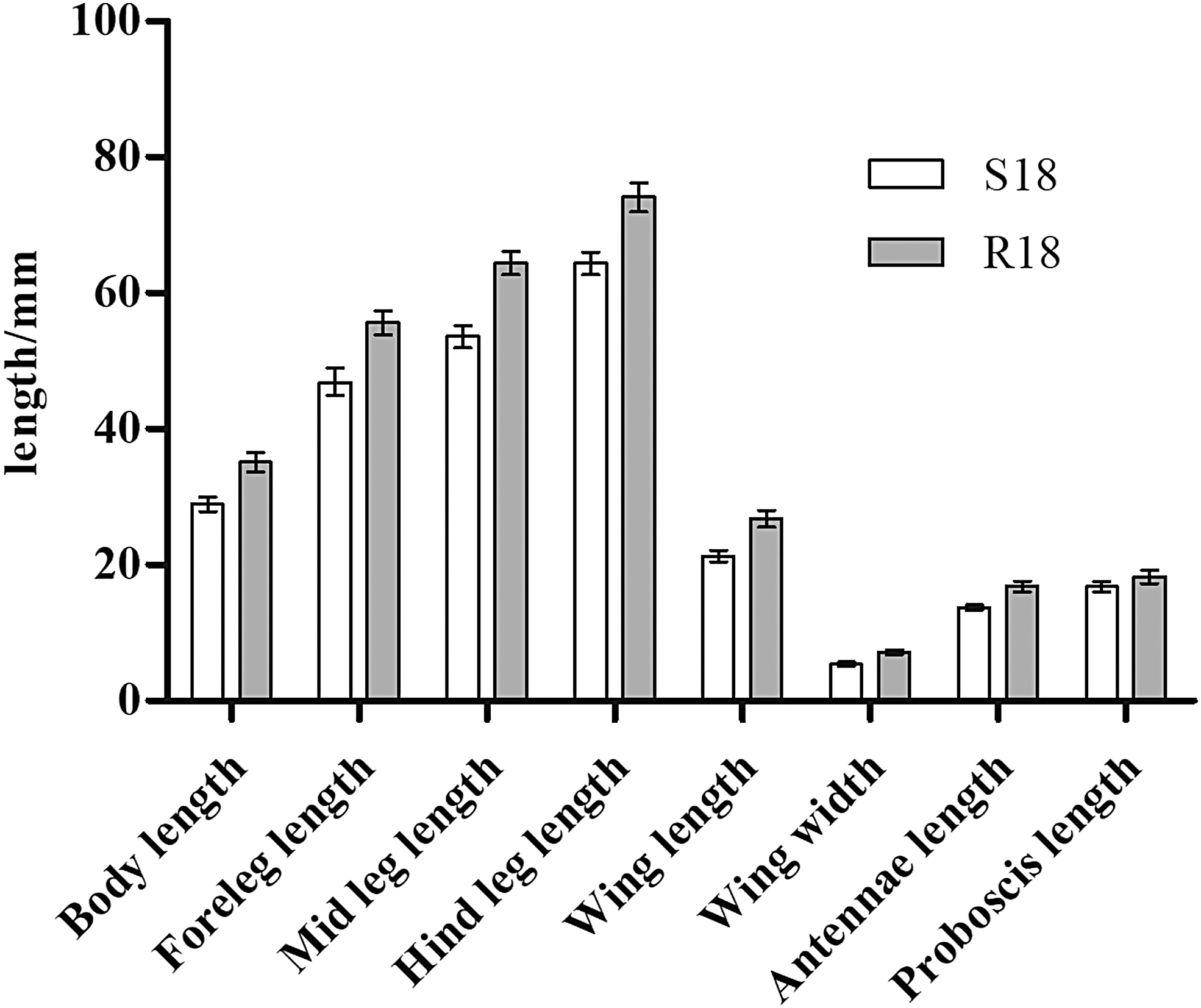
Comparisons of the mean morphological indexes in 18th-generation deltamethrin-resistant Aedes albopictus and the sensitive control.
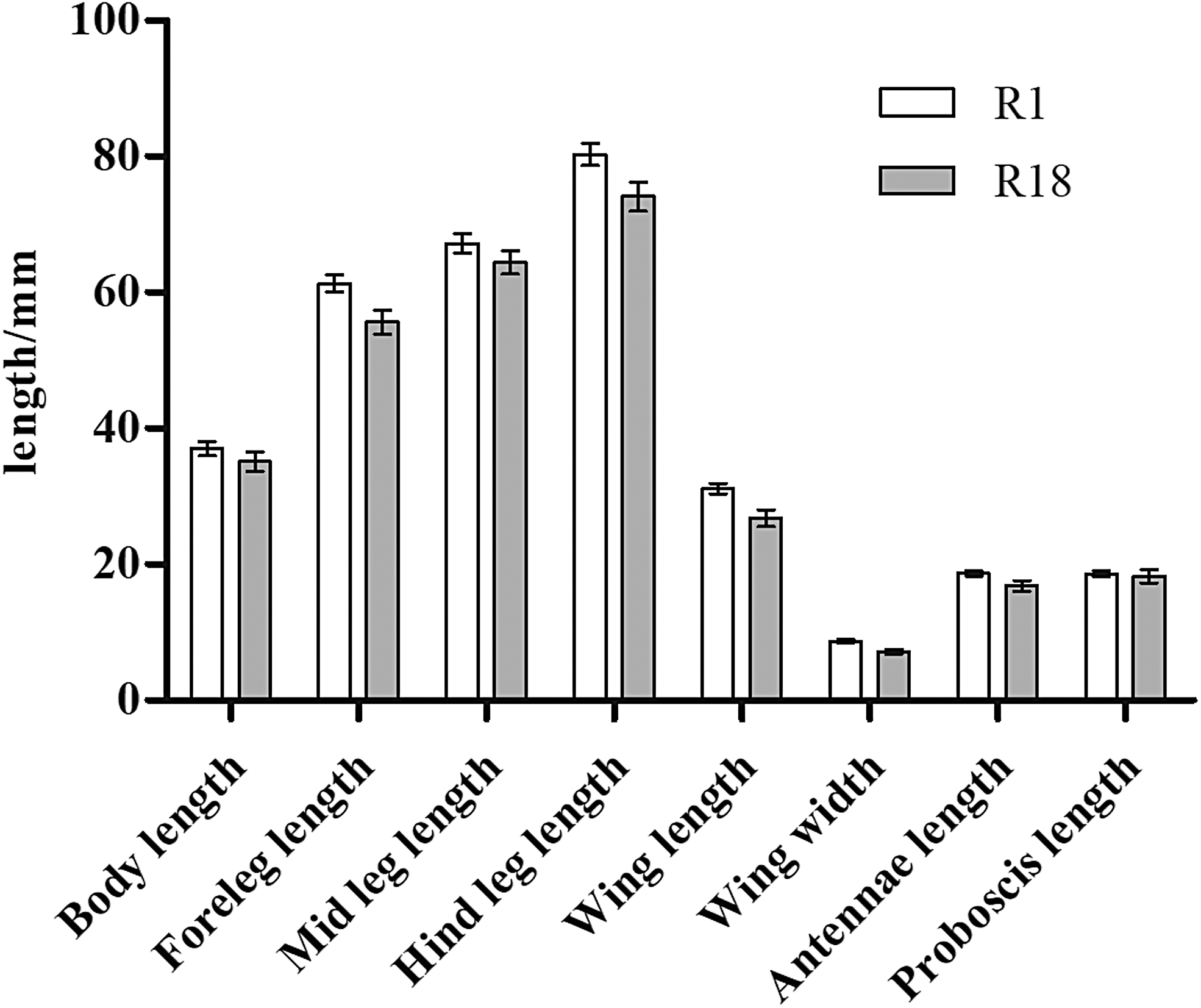
Comparison of mean morphological indexes between 1st- and 18th-generation resistant Aedes albopictus.
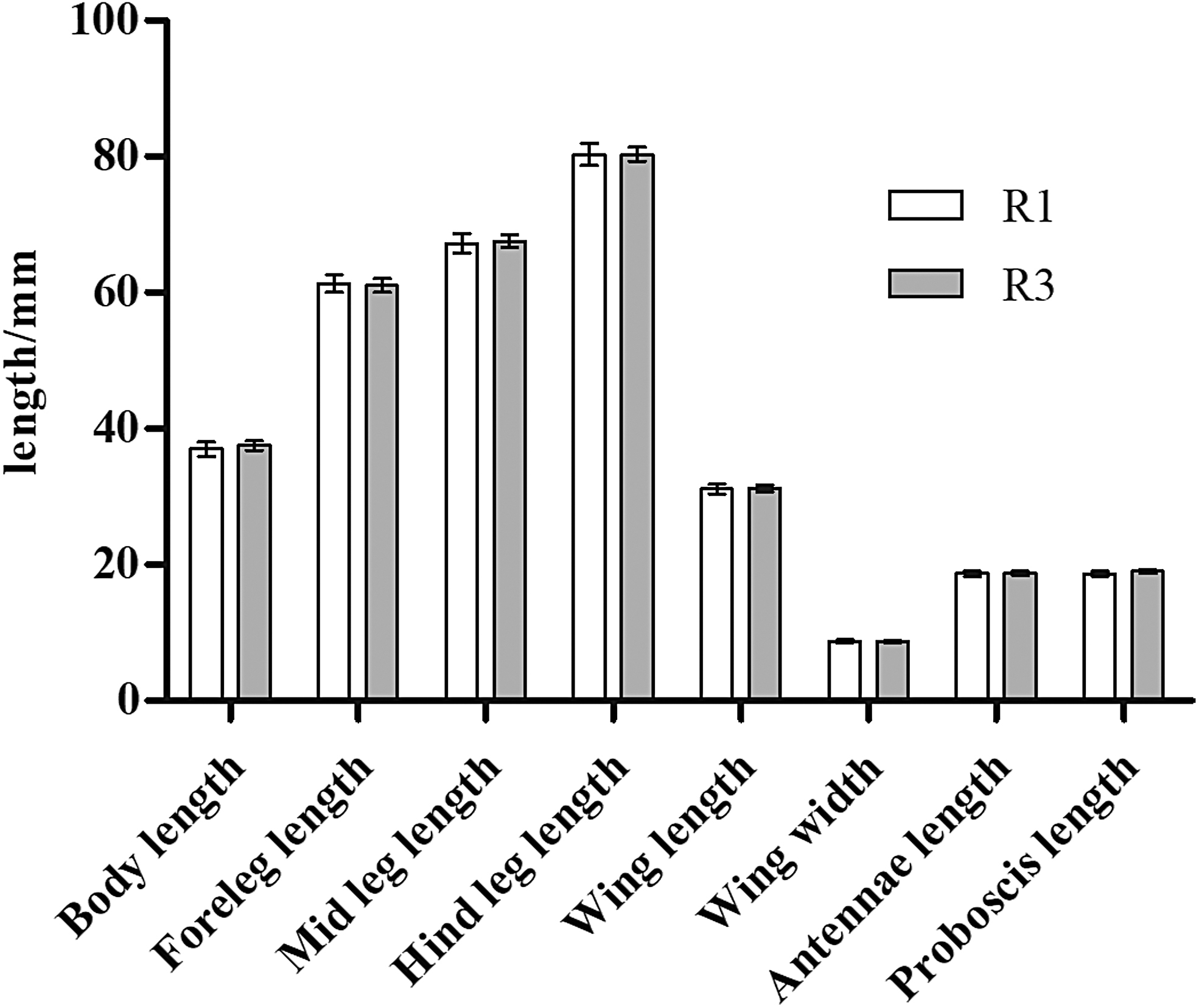
Comparison of mean morphological indexes between 1st- and 3rd-generation resistant Aedes albopictus.
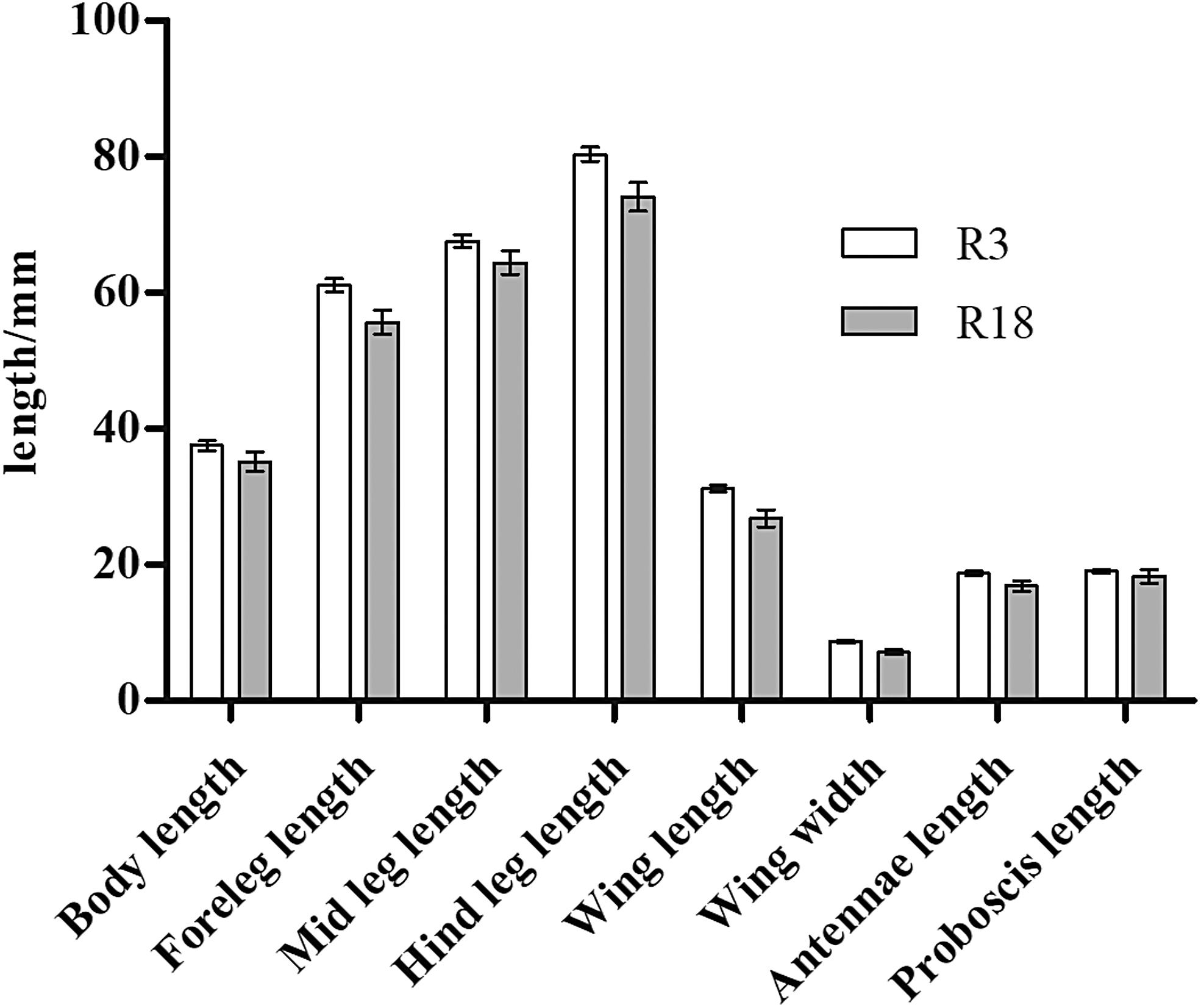
Comparison of mean morphological indexes between 3rd- and 18th-generation resistant Aedes albopictus.
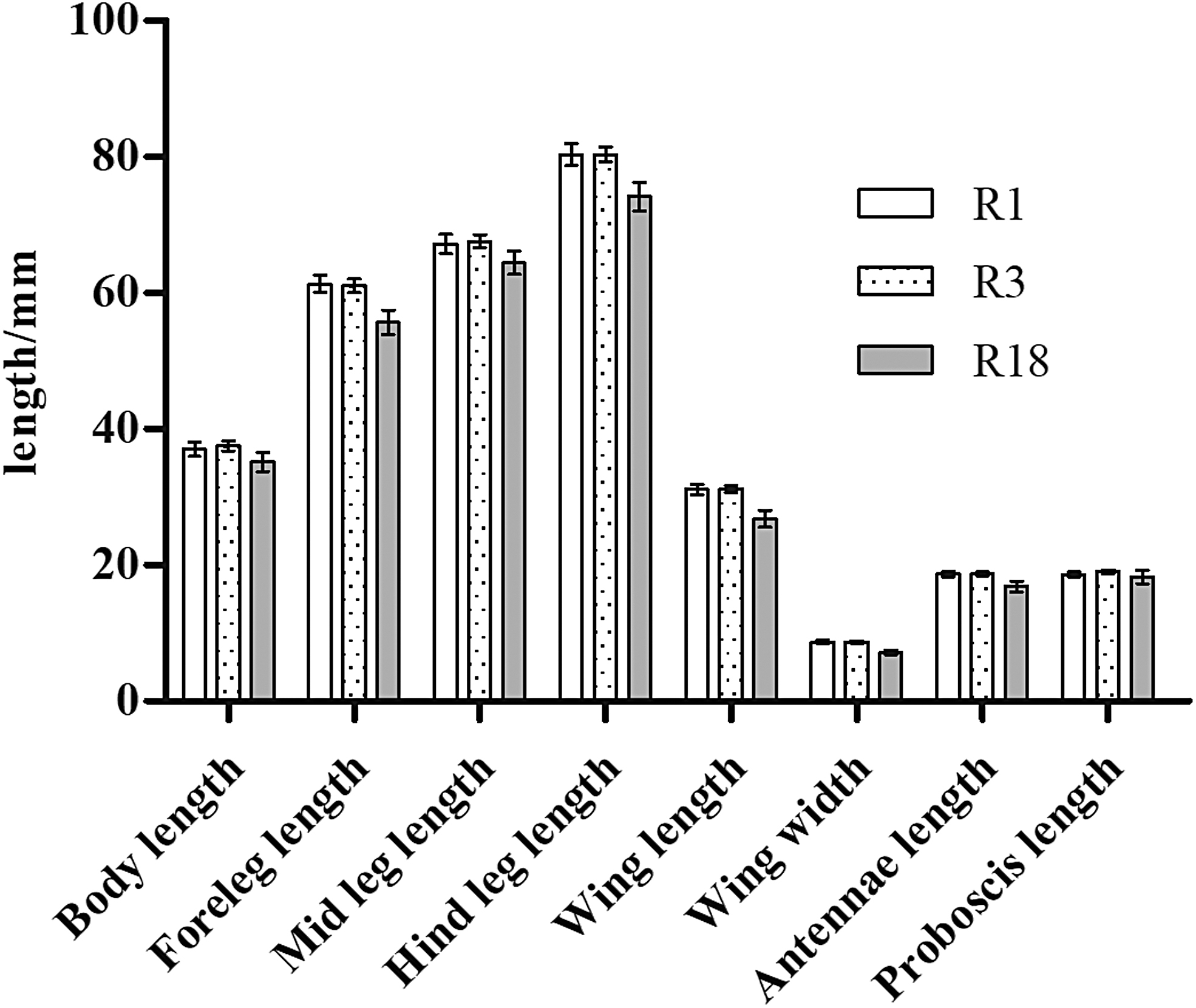
Comparison of mean morphological indexes among 1st-, 3rd-, and 18th-generation resistant Aedes albopictus.
Morphological Differences Between Generations of Resistant and Susceptible Strains of Aedes albopictus
S represents the deltamethrin insecticide-sensitive strain; R represents the deltamethrin insecticide-resistant strain. S-18 represents the deltamethrin insecticide-sensitive strain, 18th generation; R-3 represents the deltamethrin insecticide-resistant strain, 3rd generation; R-18 represents the deltamethrin insecticide-resistant strain, 18th generation; using the χ test group difference, p represents probability.
Reproductive potential
Reproductive fitness was subsequently compared between the SS strain and R-1 and R-3 DLR strains. As a result, significant differences in egg hatching rates were observed between the SS and DLR strains (χ 2 = 174.551, p < 0.001; Table 3 and Supplementary Table S1); there were also significant differences in pupation rates (χ 2 = 15.019, p < 0.001). The pupation rate was higher in the SS strain than both generations of DLR strains (Table 3; Supplementary Table S1). Before selection, the ratio of females to males of the susceptible strain S1 was 0.205714, with fewer females and more males. After deltamethrin resistance selection, the female/male ratio of mosquitoes decreased after pupa hatching, with more females and fewer males. For example, the female/male ratio of the R-5 generation without deltamethrin selection was 0.406015, and the female/male ratio increased to 1.960784 after deltamethrin selection. Before deltamethrin selection, the mean hatchability of pupae in different sensitive groups was high; in the S1A1-A3 group, mean hatchability of pupae is 72.33%. After deltamethrin selection, the hatchability of pupae in the resistant group decreased; in the R3A1-A3 group, mean hatchability of pupae is 42.66% (Table 4).
Analysis of Reproductive Fitness of Aedes albopictus Between the Sensitive Group and Deltamethrin-Resistant Breeding Group
χ = 174.551, p < 0.001. Rates of egg hatchability among groups were significantly different.
(1) Fecundity test results of the sensitive strain (s-1): A represents the combination of 25 female mosquitoes and 5 male mosquitoes, repeated 3 times. (2) B represents the combination of 25 male mosquitoes and 5 female mosquitoes, repeated 3 times. (3) Test results of fecundity of the R-1 generation, A represents the combination of 25 female mosquitoes and 5 male mosquitoes, repeated 3 times. (4) Test results of fecundity of the R-3 generation, A represents the combination of 25 female mosquitoes and 5 male mosquitoes, repeated 3 times.
Aedes albopictus Sex Ratios of S and R Groups Under Laboratory Conditions, Throughout Generations
S represents the deltamethrin insecticide-sensitive strain, the number and proportion of males and females in the S-1 control group. R represents the deltamethrin insecticide-resistant strain. Number and ratio of females and males in R-3 representative of the deltamethrin insecticide selection group after incubation. Number and ratio of females and males in R-4 representative of the deltamethrin insecticide selection group after incubation. R-5* represents the number and ratio of females and males after incubation in the selection group without deltamethrin; number and ratio of females and males in R-5 representative of the deltamethrin insecticide screening group after incubation. Number and ratio of females and males in R-6 representative of the deltamethrin insecticide selection group after incubation.
Detection of kdr mutations
No changes in the amino acid sequence of the kdr gene were observed between the resistant mosquitoes selected in the laboratory and sensitive strains reared in the laboratory; however, mutations in the nucleic acid sequence were observed in the resistant strain. Base mutation at the 96th locus of the kdr gene from C to T was observed in the resistant compared with the sensitive strain and at the 111th locus from A to G, the 132nd locus from C to T, the 204th locus from C to T, and the 534th locus from T to C.
Discussion
The continual usage of insecticides during the early 20th century in agriculture and public health has contributed to development of insecticide resistance in mosquito vectors in many countries. Determination of mosquito resistance status is essential because vector control using insecticides is the key strategy. There are many reports about collecting A. albopictus in the field to analyze the resistance to deltamethrin insecticides (Gao et al., 2018, Zhou et al. 2019). Deltamethrin resistance was analyzed by catching A. albopictus in the field; the analysis effect is affected by many insecticides that interact with A. albopictus in the field environment. Thus, studying deltamethrin resistance in the laboratory using a single DLR mosquito strain with a clear genetic background would aid our understanding of the molecular mechanism of resistance.
This study carried out successful laboratory selection of deltamethrin-resistant A. albopictus up to the 20th generation. After resistance pressure selection, the LC50 increased from 0.0070 mg/L in S-1 to 0.0563 mg/L in larvae from the resistant strain R-20, resulting in an RR of 11.26. Chen et al. (2008) selected dichlorvos dimethyl dichloroviny phosphate (DDVP) resistance in Culex pipiens pallens over 43 generations. As a result, they observed an increase in the LC50 from 0.00954 ppm in the sensitive group to 1.1607 ppm in the 43rd generation, suggesting an increase in resistance of 12.17. However, they also found that development of DDVP resistance was slow in the early stage of establishment up until the 20th generation, after which a rapid increase was observed and maintained up to 30 generations. From there on, an increase in resistance from 8.17 to more than 12 was observed up to the 43rd generation. Chen et al. reported results that are similar to our experimental results.
The resistance mechanism in A. albopictus mainly involves metabolic and target resistance. A large number of studies have shown that mutation at the kdr gene in the sodium channel in A. albopictus is related to insecticide resistance. For example, Xu et al. (2016) revealed a mutation of F1534S in the VGSC gene when studying field resistance in A. albopictus from different countries. Following laboratory-based selection of deltamethrin resistance in A. albopictus, we found multiple base mutations in the kdr gene in 19th-generation mosquitoes. In this study, we did not perform the metabolic resistance test and gene heterozygote observation, which is also a limitation of this study.
Furthermore, differences in the morphological data were revealed between the deltamethrin-resistant and -sensitive strains, with an overall decrease in all body size indexes. Moreover, as screening generations increased, the morphological phenotype of the resistant strains continued to change, suggesting that long-term selection results in increasing morphological variation. Moreover, Chaiphongpachara and Laojun (2019b) reported variation with time in wing size and shape in the coastal malaria vector Anopheles (Cellia) epiroticus Linton and Harbach (Diptera, Culicidae) in Samut Songkhram Province, Thailand. They further revealed that annual variation in weather conditions affected these adaptive and morphological changes. They also reported similar annual variation in wing morphology in Culex sitiens Wiedemann (Diptera, Culicidae) mosquito vectors in coastal Samut Songkhram (Chaiphongpachara and Laojun 2019a), with significant differences in wing size and shape between observation years. Differences in rainfall were found to affect these morphological changes, presumably reflecting environmental adaptation.
In this study, reproductive fitness was also compared between sensitive (SS) and R-1 and R-3 DLR strains of A. albopictus selected in the laboratory. No differences in egg hatching rates were observed between the SS and either DLR strain; however, pupation rates were significantly lower in the DLR strains (χ 2 = 15.019, p < 0.001). Before selection, the ratio of females to males of the susceptible strain S1 was 0.205714. After deltamethrin resistance selection, the female/male ratio of mosquito pupae increased after pupa hatching, with more females and fewer males. Before deltamethrin selection, the mean hatchability of pupae in different sensitive groups was high, but after deltamethrin selection, the hatchability of pupae in the resistant group decreased. With deltamethrin action and mosquito pressure increasing, mosquito reproductive fitness showed a difference. After deltamethrin resistance selection, the female/male ratio of mosquitoes increased after pupa hatching, with more females and fewer males, hinting at increasing chances of more female mosquitoes biting humans, increasing the risk of mosquito vector disease transmission. The results demonstrate the importance of epidemiology values.
Martins et al. also reported the effect of insecticide resistance on development, longevity, and reproduction in field and laboratory-bred A. aegypti populations. They examined larval development, the sex ratio, adult longevity, relative amounts of ingested blood, rates of ovipositing females, egg laying, and egg viability. In the field populations, the effects on all parameters were small, but directly proportional to resistance (Martins et al. 2012). The above report is similar to this study. Resistance has a significant adaptive value in the presence of insecticide treatment.
In conclusion, this study successfully screened deltamethrin-resistant strains of A. albopictus under laboratory conditions. Compared with experiments in the field, our laboratory conditions excluded the influence of confounding factors such as the external environment, resulting in a single influencing factor. As a preliminarily first step, we screened A. albopictus up to the 20th generation, revealing morphological variation and mutation of domain III of the sodium channel gene. These findings therefore provide model biological samples for further molecular and proteomic studies on the mechanisms of deltamethrin resistance in A. albopictus. The morphological phenotypes of the deltamethrin-resistant population of A. albopictus mosquitoes had changed. After deltamethrin resistance selection, the female/male ratio of mosquitoes increased after pupa hatching, with more females and fewer males, hinting at increased chances of more female mosquitoes transmitting diseases.
Footnotes
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (no. 31630011), the Science and Technology Plan Project of Guangzhou (no. 201804020084), and the Natural Science Foundation of Guangdong Province (no. 2017A030313625). The funders had no role in study design, data collection and analysis, the decision to publish, or preparation of the manuscript.
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.