
Abstract
Chikungunya virus (family Togavirdae and genus Alphavirus) is an emerging and reemerging virus of public health importance both regionally and globally. In Kenya, about 50–60% of the suspected measles cases remain undiagnosed once measles and rubella is ruled out by immunoglobulin M (IgM) ELISA thus prompted the need to do differential diagnosis on the measles/rubella negative samples. Nothing is known about the role played by chikungunya infection among these suspected measles cases. Febrile rash illness is a common clinical presentation of arboviruses, including chikungunya. In this study, we conducted a serosurvey to explore the possible role of chikungunya infections among suspected measles cases in Kenya that had tested negative for measles and rubella. Sera were tested by commercially available ELISA for the presence of IgG and IgM antibodies against the chikungunya virus. All positive samples for chikungunya by ELISA were confirmed by plaque reduction neutralization test (PRNT), and to rule out cross-reactivity with other alphaviruses a panel of viruses was used, namely o' nyong' nyong, Semliki Forest, and Sindbis viruses. Of the 392 serum samples screened, 0.3% (n = 1) tested positive for IgM antibodies, while 4.6% (n = 18) tested positive for IgG antibodies against the chikungunya virus. PRNT results indicated 2 (11%) chikungunya positives and 7 (38.9%) o' nyong' nyong positives. We recommend awareness among health care providers and improved surveillance for these arboviruses by both serology and molecular testing. Testing for other pathogens should also be done to improve disease detection and diagnosis.
Introduction
Measles infection is one of the causes of high morbidity and mortality among children in developing countries (Perry et al. 2004, Nandy et al. 2006). Clinical symptoms caused by measles infection include cough, coryza, and conjunctivitis, followed by a maculopapular rash and high fever (39–40°C) (WHO 2015). In Kenya, all suspected measles cases, are routinely submitted to the measles laboratory at the Kenya Medical Research Institute's (KEMRI), Centre for Virus Research (CVR). At the CVR, the samples are tested for measles. If negative, cases are tested for rubella IgM antibodies in support of the national measles/rubella case-based surveillance program. Over the years, it has been observed that ∼50–60% of these suspected measles cases usually test negative for both rubella and measles using enzyme-linked immunosorbent assay (ELISA).
Chikungunya (CHIKV) and other arboviruses such as o' nyong' nyong (ONNV), Sindbis (SINV), and Semliki Forest virus (SFV) are among arboviruses causing emerging and reemerging infections of public health importance regionally and globally (Paixao et al. 2018). It has been shown that CHIKV is endemic in Coastal and Northern parts of Kenya (Sergon et al. 2004, Konongoi et al. 2018). It has caused frequent outbreaks in the country (WHO 2016, 2018, Konongoi et al. 2018). Even though CHIKV, ONNV, SFV, and SINV are known to be circulating in Kenya (Morrill et al. 1991, Sergon et al. 2008, Mease et al. 2011, Sutherland et al. 2011, LaBeaud et al. 2015, Ochieng et al. 2015, Tigoi et al. 2015, Grossi-Soyster et al. 2017), no study has been conducted to test these arboviruses on non-measles cases that present as febrile rash illness. In addition, the disease burden of these arboviral diseases in Kenya is not well defined. To address these gaps, this study set out to determine the exposure of chikungunya virus among suspected measles cases collected in the measles case-based surveillance program in Kenya. This was to assess the possible contribution of chikungunya infections to cause measles-like illness.
Materials and Methods
Study design
We conducted a laboratory-based, retrospective study using 392 randomly selected archived human sera. These samples were collected between January 2008 and December 2014 from health facilities performing measles case-based surveillance in 10 counties from 3 regions in Kenya. These include Nairobi County, Coastal region (Lamu, Taita-Taveta, Mombasa, Tana River, Kwale, and Kilifi), and North-Eastern region (Mandera, Wajir, and Garissa) as shown in (Fig. 1). These counties were preselected because of past or recent reports of CHIKV outbreaks (Coastal and North Eastern regions) or because they are a major travel hub with a high risk of introduction and transmission (Nairobi County).
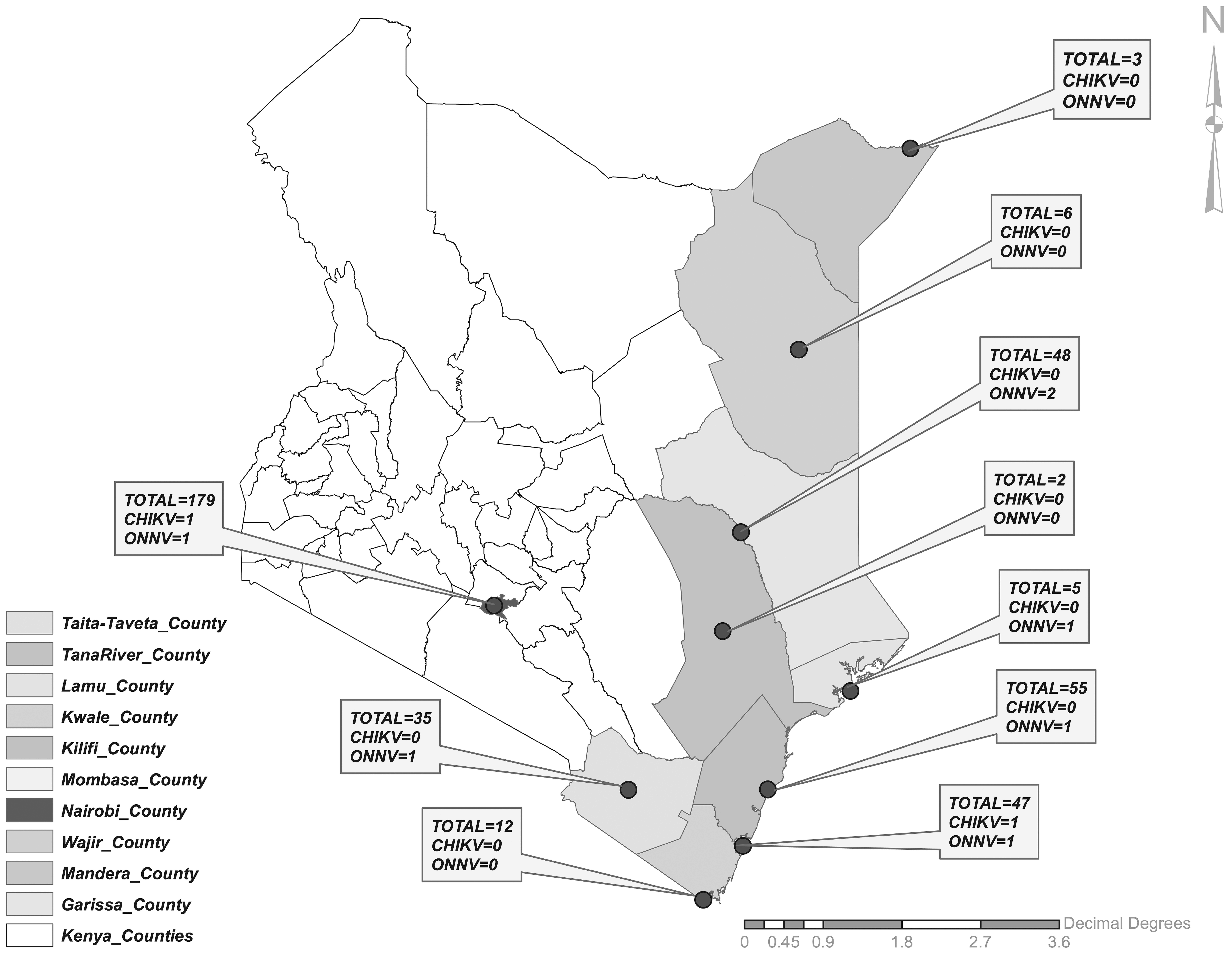
Map of Kenya showing the study sites and total numbers of samples used per site: Nairobi Region, Coast Region (Mombasa, Kwale, Kilifi, Tana-River, Taita-Taveta, and Lamu), and North-Eastern Region (Wajir, Mandera, and Garissa) and number of CHIKV and ONNV positives in each county.
The 392 archived serum samples used in this study were selected based on probability proportional to size from the total number of samples collected from each of the three regions: Nairobi n = 179 (46%), North-Eastern n = 57 (14%) and Coast n = 156 (40%).
This study was approved by Kenya Medical Research Institute's Scientific and Ethics Review Unit (SERU no.3122).
Serum specimens' collection, processing, storage, and transportation
Samples were collected from all counties in Kenya. Measles surveillance in Kenya follows World Health Organization guidelines, in which a suspected measles case is defined by a patient having fever and one of the following symptoms: rash, cough, coryza, or conjunctivitis (WHO 2015). Five milliliters whole blood was collected from all suspected cases, and serum was separated at the various health facilities then shipped to KEMRI's reference laboratory for testing by ELISA method. In addition, from each suspected patient, a case investigation form was filled and the information recorded included; region, health facility, gender, age, date of birth, residence, date of onset of rash, presence of fever, type of rash, measles vaccination history, status of the patient, tracer information, date of specimen collection, and type of specimen. The form was sent together with the serum to the laboratory. The serum was stored in a biobank at −20°C.
Laboratory testing
Enzyme-linked immunosorbent assay
CHIKV IgM and IgG antibodies were measured to assess serological evidence of recent exposure (IgM) and prior exposure (IgG) to CHIKV.
CHIKV IgG and IgM ELISA
All the 392 serum samples were heat-inactivated at 56°C for 30 min before testing. We screened the specimens for anti-CHIKV antibodies using commercial IgG and IgM capture ELISA kits (Novatec Immunodiagnostica GmbhH) according to the manufacturer's instructions. Briefly, CHIKV IgG microplates were coated with anti-human IgG antibodies. The diluted samples were added and incubated at 37°C for 1 h. After 1 h, the wells were washed to remove all unbound antibodies, the antigen was added, and the plates incubated at room temperature for 30 min. Next, the plates were washed, and the detector antibody was added into all the wells. The plates were incubated at room temperature for 30 min. After washing, the horseradish peroxidase-labeled streptavidin was added to bind to the captured specific immune complex during incubation at room temperature for 30 min and washed. The immune complexes were visualized by adding Tetramethylbenzidine (TMB) substrate, which gave a blue color. Stop solution (0.2 M sulfuric acid solution) was added and incubated for 15 min to stop the reaction that produced a yellow color. The absorbance was measured at 450 nm using ELISA plate reader ELx800 (Biotek).
CHIKV IgM antibodies were detected using microplates that were coated with anti-human IgM antibodies to bind to the corresponding antibodies of the diluted samples. All the other steps were followed as described for CHIKV IgG and the absorbance was measured at 450 nm using ELISA plate reader ELx800 (Biotek).
Plaque reduction neutralization test
Plaque reduction neutralization test (PRNT) assays were undertaken to confirm the CHIKV IgG and IgM ELISA positive results and to rule out cross-reactivity with other closely related alphaviruses. All the alphavirus isolates used in this study were obtained from KEMRI Viral Haemorraghic Fever (VHF) laboratory; including CHIKV Lamu 33 (Gen Bank accession no. HQ456255), (Kariuki Njenga et al. 2008), SFV ATH00510 (Gen Bank accession no. KF283988), (Lwande et al. 2013), SINV NVS/305 (Gen bank accession no KY616988), (Sigei et al. 2018), and ONNV SG 650 (Gen bank accession no AF079456), (Lanciotti et al. 1998). In brief, all the viruses were pretitred using plaque assay, then diluted in maintenance media (Minimum Essential medium Sigma with Earle's salts with reduced NaHCO3(Sigma), supplemented with 2% heat-inactivated fetal bovine serum (FBS) (Gibco), 2% L-glutamine (Sigma), and 2% penicillin/amphotericin B (Sigma) to a standard concentration that produced ∼20–50 plaques per 100 μL. Each serum sample was serially diluted from 1:20 to 1:320 using maintenance media in sterile Eppendorf tubes. A constant volume of the diluted virus with the required concentration was added into each 1.5 mL tube (Eppendorf) containing the diluted serum from 1:20 to 1:320 and incubated for 1 h at 37°C. The virus-serum mixture (30 μL of the diluted virus with 30 μL of the diluted serum) was then transferred to 24-well plates (Corning) with confluent Vero cell monolayers (African green monkey cells ATCC) and incubated at 37°C, 5% CO2 for 1 h for adsorption process, after which an overlay of 2.5% methylcellulose (Sigma) was added. After 3–5 days, depending on the virus tested, the plates were fixed with 3% formalin (Sigma) and stained with 0.5% crystal violet (Sigma) prepared in absolute ethanol (Sigma). For etiological diagnosis, the PRNT90 antibody titer for each virus was required to be four-fold greater than the other viruses in the same family to arrive at an etiological diagnosis (Tigoi et al. 2015, Chepkorir et al. 2019). A sample was considered indeterminate if it neutralized two viruses and did not meet the four-fold greater threshold.
Data analysis
Data analysis was performed using SPSS version 22. The IgM and IgG titers for CHIKV were dichotomized as either positive or negative based on optical density (OD) values. PRNT results were stratified by the age of the patients, gender and county of occurrence, and by region (North-Eastern, Coast, and Nairobi). Association of seropositivity to CHIKV and ONNV with age, gender, and site of collection was tested using chi-square. The proportion of positive sample for each virus was determined.
Results
Patient demographic characteristics
A total of 392 archived human serum samples were analyzed (Nairobi (n = 179) (46%), North-Eastern (n = 57) (14%), and Coast (n = 156) (40%)), the median age of the suspected measles cases was 4.7 years and interquartile range (IQR) was 2.8–6.7 years. Of the 392 samples, 52.8% were <5 years, 34.4% were 5–9 years, 6.4% were 10–14 years, and 6.4% were >14 years of age. One hundred and fifty-eight (40%) were female and 234 (60%) were male (Table 1).
Patient Demographic Characteristics Collected from Coast, North-Eastern, and Nairobi Regions in Kenya (2008–2014)
Serological detection of ONNV and CHIKV
Out of the 392 samples tested, 1 (0.3%) sample tested positive for both anti-CHIKV IgM/IgG and 18 (4.6%) tested positive for anti-CHIKV IgG. Out of the 18 samples that were positive by ELISA, 9 (50%) were positive by PRNT. Only 2 (11%) had neutralizing antibodies against CHIKV while 7 (39%) for ONNV (Fig. 1). The other 9 samples (50%) had neutralizing antibodies for both CHIKV and ONNV, which did not meet the four-fold greater threshold hence considered indeterminate (Table 2). None of the samples had neutralizing antibodies against SINV and SFV. CHIKV was detected from a sample collected in Mombasa County in 2014 while the other one was collected from Nairobi County in 2009. ONNV was detected in the years 2009 (3/7), 2010 (2/7), 2011 (1/7) and 2014 (1/7). The counties in which ONNV was detected were Nairobi (1/7), Garissa (2/7), Mombasa (1/7), Kilifi (1/7), Taita-Taveta (1/7), and Lamu (1/7) (Table 2).
Chikungunya and O'nyong-Nyong Virus Plaque Reduction Neutralization Test Positivity by Endpoint Titers for Archived Serum Samples from Coast, North-Eastern, and Nairobi Regions in Kenya (2008–2014)
PRNT, plaque reduction neutralization test; M, male; F, female; CHIKV, chikungunya virus; ONNV, o 'nyong' nyong virus; SINV, sindbis virus; SFV, semliki forest virus; (—), titer <1:20 and IND-indeterminate.
There was no significant difference in seropositivity of CHIKV and ONNV with age, gender, and site of collection (Table 3).
Association Between Seropositivity of Chikungunya and O 'nyong' nyong Viruses with Age, Gender, and Site
Discussion
This study found two cases of chikungunya infection among samples that were negative for acute measles in selected areas in Kenya during the 2008–2014 period. This finding of CHIKV in negative measles cases is an indication of how such co-circulation of viruses with similar clinical presentation could complicate measles diagnosis. The low prevalence rate of CHIKV observed in this study could be attributed to the early timing of sample collection. It is known that CHIKV IgM antibodies are highly detectable 4–20 days after the onset of illness (Natrajan et al. 2019) and may persist for about 2 months. Therefore, samples collected during the first week after onset of illness should have preferably been tested by both serological and virological methods (RT-PCR). This was not done due to resource limitation.
In this study, CHIKV IgM antibodies were detected in a sample collected from a 6 year old who reported to a health facility with febrile rash illness. This finding suggests the circulation of CHIKV virus in Mombasa County, Coastal region, as early as 2014. The Coastal region experienced the largest outbreak of CHIKV in Lamu County in 2004 (Sergon et al. 2008). Therefore, this indicates continued circulation of CHIKV outside of the outbreaks and may also suggest it had been circulating at low level even before the reported outbreak, which occurred in Mombasa in 2018 (WHO 2018). This is likely to happen because only a few cases present with a maculopapular rash, and they easily go undetected.
The Coastal region also experienced a large dengue outbreak in 2013 (Ellis et al. 2015), 2013–2014 (Lutomiah et al. 2016), which could have led to underreporting of CHIKV because of its similar clinical symptoms (WHO 2009). The Coastal region has competent vectors for CHIKV as shown by Mulwa et al. (2018), Agha et al. (2017), and Mbaika et al. (2016), and this is attributed to the exacerbation of the intensity of the CHIKV transmission and outbreaks.
CHIKV neutralizing antibodies were also detected in a sample collected from a 5 year old in Nairobi County in the year 2009, indicative of past exposure to the virus. Nairobi County has never experienced any arboviral outbreak, when compared with Mombasa County.
O'nyong-nyong virus neutralizing antibodies were detected in Lamu, Taita-Taveta, Kilifi, Mombasa, Garissa, and Nairobi, indicating past exposure to the virus in all the three regions from samples collected between 2009 and 2014. A large number of samples neutralized with ONNV were observed in Coastal region, a finding consistent with the findings from a previous study by LaBeaud et al. 2015 that found ONNV exposure to be common in Coastal Kenya between the years 2014 to 2015. The last documented ONNV outbreak in Kenya occurred in 1961; the outbreak spread to neighboring countries, eventually affecting more than 2 million individuals but without any recorded fatalities (Rezza et al. 2017). We did not detect any neutralizing antibodies against SINV or SFV in the measles negative samples but this doesn't rule out the possibility of these viruses circulating among undiagnosed measles cases. This is because these viruses have been detected in serological surveys carried out in Kenya (Tigoi et al. 2015, Ofula et al. 2016) and also during vector surveillance carried out by Ochieng et al. 2013.
The results of this study agree with the findings by Antonio et al. 2019, where CHIKV IgM and IgG were detected in negative measles cases from 2009 to 2015 in Mozambique. Other studies have also been carried out to determine the etiologies of fever and maculopapular rash, and these have yielded results indicating that some of the etiological agents were overlooked or left out. First, in Brazil by De Moraes et al. 2011, where dengue virus was detected in 22 cases. Two of these 22 cases met Brazil's clinical case definition for rubella, which would have been reported to be rubella, but these 2 cases were diagnosed as dengue infection. Second, in Belarus, a study by Yermalovich et al. 2014 found antibodies against enteroviruses, human herpes virus, adenovirus, and human parvovirus B19 in measles or rubella suspected cases among children. Lastly, in Rio de Janeiro, Brazil a study by Oliveira et al. 2001, showed that dengue fever, rubella, parvovirus B19, measles, and human herpes virus 6 were among the causes of rash between 1994 and 1998. Possibly, the main contributor to these occurrences could be the fact that arboviruses are overlooked because of unavailability of resources in terms of equipment for diagnostics assays and specialized laboratories.
In this study, cross-reactivity was observed in samples that had tested positive for anti-CHIKV IgG on ELISA neutralizing with ONNV on PRNT assay. This finding is consistent with other studies reported in Kenya and Cameroon, where cross-reactivity was observed between CHIKV and ONNV (Kuniholm et al. 2006, LaBeaud et al. 2015, Tigoi et al. 2015). This could be due to the fact that both viruses may have been co-circulating in the region and continued evolution of ONNV has made it antigenically more similar to CHIKV than previously reported (Chanas et al. 1979).
Conclusion
This study found two cases of chikungunya and seven cases of o 'nyong' nyong viruses in suspected measles cases from various counties in Kenya that had tested negative for measles. There is need for continuous awareness among clinicians and the need to improve surveillance of these epidemic-prone arboviruses for better detection, outbreak detection, and clinical management.
Our findings, indicate clinical diagnosis is not sufficient to detect febrile rash cases and laboratory confirmation is essential to determine other causes of febrile rash illnesses in Kenya.
Arboviruses are overlooked because diagnostic assays are not typically available at the county and sub-county levels. The tests are often performed in very few specialized laboratories at national level. A further reason is the lack of ELISA kits for other alphaviruses, which makes it difficult to detect these viruses causing illness. We strongly recommend molecular testing of arboviruses to detect these arboviral infections in Kenya and testing for additional pathogens should also be considered in measles/rubella control.
We consider this study a success; but at the same time, we acknowledge existing limitations. First, over 90% of the study participants are children, given the surveillance of measles and rubella is focused on children. Second, molecular testing of alphaviruses was not possible due to limited resources. Third, logistically speaking it would be difficult to obtain convalescent samples having worked on archived samples. Fourth, we were also not able to test for other pathogens due to limited testing panels, and finally, the travel history of the patients was not captured during sample collection.
Footnotes
Acknowledgments
We acknowledge the Expanded Programme on Immunization (EPI) laboratory at KEMRI for providing archived measles surveillance samples under World Health Organization/Ministry of Health surveillance programme. We thank Arbovirology/Viral Haemorrhagic Fever Laboratory and staff in the Centre for Virus Research (CVR) at KEMRI for providing an excellent environment to carry out the study.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was obtained to carry out this study.