
Abstract
Japanese encephalitis virus (JEV) is recognized as a public health risk by the World Health Organization. In Asia, each year, ∼70,000 people become infected with JEV, which results in ∼10,000 deaths. Chikungunya virus (CHIKV) is an RNA virus, whose infection mainly causes fever, myalgia, and skin rash. Although the mortality rate is low, it seriously affects daily life. JEV and CHIKV infect humans through mosquitoes; therefore, a recombinant vaccinia virus coexpressing JEV E and CHIKV E1 proteins was constructed to prevent their concurrent infection. In this study, after mice first immunization, booster immunization was performed at 21 days postimmunization (dpi). At 35 dpi, mice were challenged with JEV and CHIKV. Specific antibodies significantly increased in the rVTT-CE1-JE-EGFP group, which were significantly (p < 0.01) higher than those of the control groups at 35 dpi. The plaque reduction neutralization tests (JEV) of rVTT-CE1-JE-EGFP group was 1:320 at 35 dpi. Furthermore, cytokine levels and the percentage of CD3+CD4+ and CD3+CD8+ T-lymphocytes in the rVTT-CE1-JE-EGFP group were significantly (p < 0.01) higher than those in the control groups at 35 dpi. After challenge, mice body weights in rVTT-CE1-JE-EGFP group were not significantly altered, and the survival rate was 100%. These results showed the rVTT-CE1-JE-EGFP group elicited significant humoral and cellular immune responses, thus indicating that the recombinant vaccine may serve as a candidate for effective prevention of CHIKV and JEV infection.
Introduction
Japanese encephalitis virus (JEV) is a positive-strand RNA virus of the family Flaviviridae (Vaughn and Hoke 1992, Unni et al. 2011), which is mainly transmitted by Culex tritaeniorhynchus mosquitoes. In 1935, JEV was first isolated from a human case in Japan, which was isolated and passaged in mice brain. JEV caused serious viral encephalitis in human throughout Asia with ∼70,000 human cases each year and ∼10,000 fatalities (Mansfield et al. 2017). Similarly, chikungunya virus (CHIKV) is an RNA virus (Alphavirus genus, Togaviridae family) first isolated in humans in Tanzania in 1953 (Ross 1956). It is mainly transmitted by Aedes aegypti and Aedes albopictus mosquitoes. Although mortality rates are rarely associated with CHIKV (∼1 of 1000 cases) (Borgherini et al. 2007, Lemant et al. 2008), the high incidence rate of acute debilitating polyarthralgia (∼ 95%) (Queyriaux et al. 2008, Sissoko et al. 2008, Weaver and Lecuit 2015) and subsequent chronic polyarthralgia (12–75%) (Brighton et al. 1983, Sissoko et al. 2009) lead to significant health and financial burdens (Yang et al. 2017).
Japanese encephalitis outbreaks were reported in South and Southeast Asia in recent years (Mansfield et al. 2017). In 2005–2006, an unprecedented chikungunya fever (Powers and Logue 2007, Schwartz and Albert 2010) occurred in the Indian Ocean, and outbreaks in India, Sri Lanka, and Southeast Asia from 2007 to 2010 (Laras et al. 2005, Kumarasamy et al. 2006, Ravi 2006, Watanaveeradej et al. 2006, Her et al. 2009, Senanayake et al. 2009) were reported. According to our epidemiological investigation, JEV and CHIKV were also detected in mosquitoes from Yunnan province in 2018 and 2019, China (unpublished observations). It is important to prevent Japanese encephalitis and chikungunya fever in South and Southeast Asia at the same time. Therefore, a bivalent genetic engineering vaccine is needed for JEV and CHIKV prevention (Chen et al. 2017).
JEV mainly encodes the structural proteins capsid (C), pre-membrane/membrane (pre-M/M), and envelope glycoprotein (E), and the nonstructural proteins NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5 (Mukhopadhyay et al. 2005). JEV E protein is the main flavivirus surface peptide, which is involved in viral attachment, fusion, penetration, and other activities. In addition, some studies suggested that E protein is a candidate for JEV subunit candidate vaccine (Chavez et al. 2010). CHIKV has two major envelope proteins, E1 and E2, which are components of the viral spikes, which facilitate viral attachment to cell surfaces and entry to the cells. The expressed E1 and E2 proteins both induce specific antibodies, and E1 was more immunogenic. However, in immunofluorescence assays in infected cells, E2 antibody showed significantly more specificity in detecting CHIKV (Abraham and Eswaran 2015).
Poxvirus is a large double-stranded DNA virus that replicates in cytoplasm (Zhang et al. 2013). Despite the global eradication of smallpox in 1979, vaccinia virus (VACV) has been used as a vaccine vector for decades. In addition, VACV TianTan (VTT) strain was isolated from skin lesions of a Chinese patient with smallpox in 1926 by Qi Changqing (Qin et al. 2013). Furthermore, VACV was used as a vector for the development of human immunodeficiency virus (HIV) vaccines (Liu et al. 2012). Previous reports showed E3L deletion mutations in Copenhagen and NYCBH VACV strains were highly attenuated and were more suitable for human vaccine vector use (Li et al. 2016, Vijaysri et al. 2008).
The aim of this study was to develop a vaccine that effectively prevents JEV and CHIKV infection. We used the E3L gene deletion mutant VACV strain TianTan as a vector to construct the recombinant vaccinia virus rVTT-JE-CE1 vaccine, which coexpresses E protein of JEV and E1 protein of CHIKV, respectively. In vivo immunogenicity of recombinant vaccine was tested in mice.
Materials and Methods
Experimental animals, viruses, cells, and plasmids
Six-week-old female BALB/c mice (Experimental Animal Center, Academy of Military Medical Science of PLA, Beijing) were used. The CHIKV virus (GenBank: KU561451.1) was provided by Yunnan Institute of Parasitic Diseases, whereas the JEV virus (GenBank: JQ086762.1) was isolated in our laboratory. The TE3L deletion mutant of recombinant VACV strain TianTan (rVTT) (Wang et al. 2012) and the shuttle vector pSTKE-EGFP plasmid (Du et al. 2012) were previously constructed. BHK-21 cells (Baby Hamster Syrian Kidney cells) were cultured in Dulbecco's modified Eagle's medium (DMEM) with 5% fetal bovine serum (FBS) and 1% penicillin–streptomycin at 37°C in a CO2 incubator.
Construction of shuttle plasmids and recombinant VACV
There were three expression cassettes MCS1, MCS2, and MCS3 in the pSTKE-EGFP, containing EGFP gene in MCS3. CHIKV E1 (GenBank: KU561451.1) and JEV whole E gene (GenBank: AF495589.1) were cloned, and respectively inserted into pSTKE-EGFP MCS1 and MCS2 to construct shuttle plasmid pSTKE-CE1-JE-EGFP (Fig. 1A).
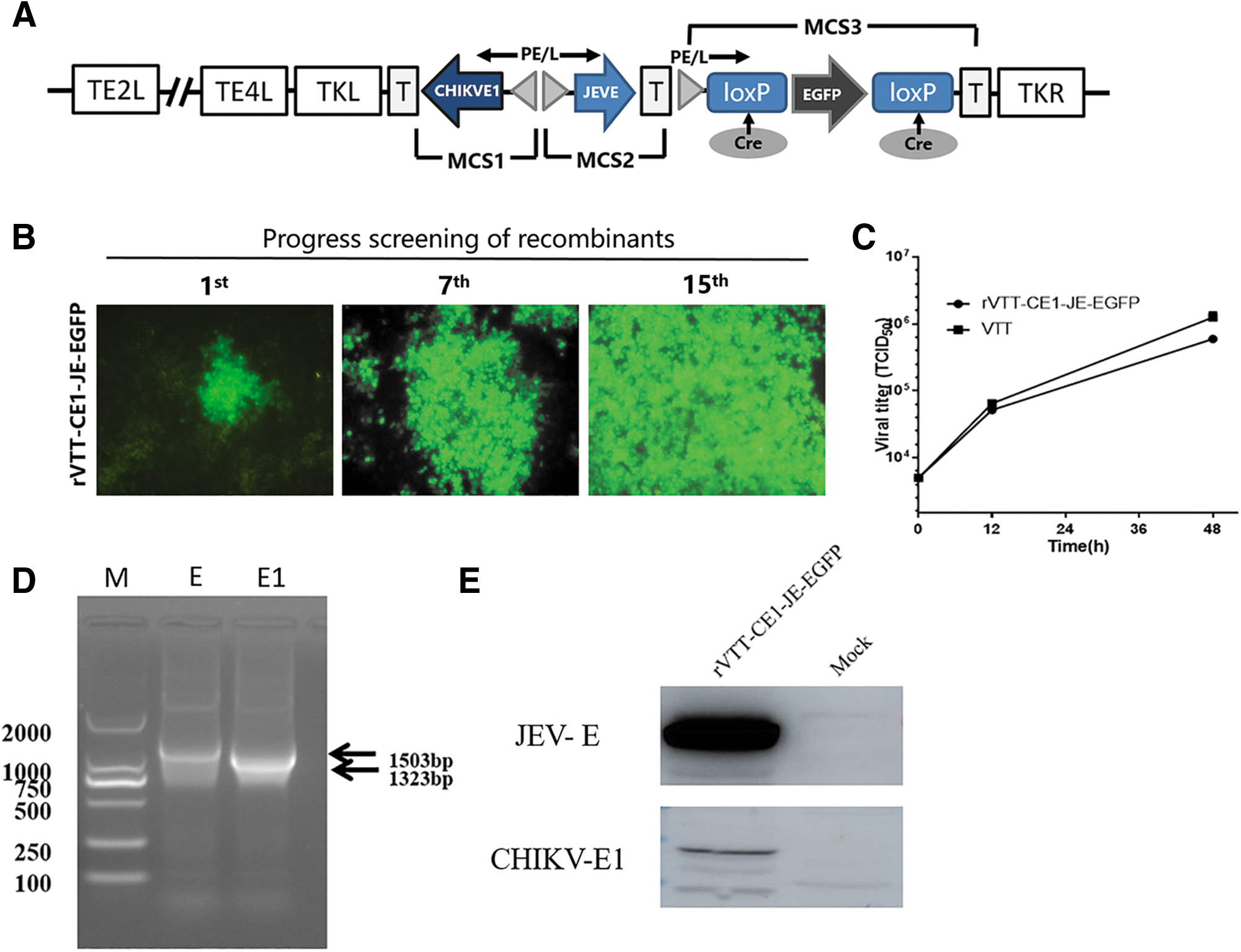
Construction and identification of the recombinant vaccinia virus.
The recombinant vaccinia virus rVTT-CE1-JE-EGFP was obtained by homologous recombination. BHK cells were infected with rVTT-EGFP at a multiplicity of infection (MOI) of 0.1 for 2–3 h; next, pSTKE-CE1-JE-EGFP plasmids were used to transfect BHK cells using Lipofectamine 3000 (Invitrogen). After 4–6 h, culture medium was aspirated and fresh medium added. Screening for fluorescent viral plaques was performed under a fluorescence microscope, as previously described (Du et al. 2012). After 15 rounds of plaque purification, rVTT-CE1-JE-EGFP was obtained.
PCR analysis of the recombinant virus
BHK cells were infected with rVTT-CE1-JE-EGFP at an MOI of 0.1 and harvested 48–60 h postinfection. RNA was extracted from recombinant viruses and subjected to reverse transcription to cDNA for use as PCR templates. Amplification of CHIKV E1 and JEV E gene sequences was performed in 30 cycles at 95°C for 30 s, 56°C for 30 s, and 72°C for 90 s, and a final extension at 72°C for 10 min with Ex-Taq (TAKARA) polymerase. Tm values of primer sequences are given in Table 1.
Sequences of Identification Primers and Corresponding Tm Values
JEV, Japanese encephalitis virus; CHIKV, chikungunya virus.
Western blotting analysis of recombinant viruses
Western blot analyses of proteins collected from BHK cells infected with rVTT-CE1-JE-EGFP at an MOI of 0.1 and harvested 36–48 h postinfection were performed. Reactions were developed with rabbit anti-CHIKV or anti-JEV polyclonal antibodies at a 1:1000 dilution as the primary antibody and horseradish peroxidase (HRP)-labeled goat anti-rabbit immunoglobulin G (IgG; Sigma-Aldrich, St. Louis, MO) at a 1:3000 dilution as the secondary antibody.
Animal immunogenicity test
Six-week-old BALB/c mice were randomly divided in 7 groups of 12 animals. Mice in groups 1 and 2 received intramuscular (IM) administration of 2 mL of 1 × 107 TCID50/mL rVTT-CE1-JE-EGFP, whereas mice in groups 3 and 4 were vaccinated IM with an identical dose of rVTT-EGFP, mice in groups 5 and 6 were IM injected with 2 mL of phosphate-buffered saline (PBS), and mice in group 7 were IM vaccinated with SA14-14-2, according to the manufacturer's instructions. Mice were boosted with the same dose at 3-week intervals after priming. Immunized mice were weekly bled from the retro-orbital sinus/plexus after priming vaccination.
Enzyme-linked immunosorbent assay
Serum samples (1:20 dilution) were tested for specific JEV and CHIKV IgG antibodies by enzyme-linked immunosorbent assay (ELISA), using as coating antigen JEV E and CHIKV E1 proteins in BL21 (DE3) and HRP-labeled rabbit anti-mouse IgG antibody (diluted 1:1000) for antibody detection at 450 nm, as previously reported (Liu et al. 2017).
Plaque reduction neutralization test
JEV neutralizing antibody titer was measured as previously reported (Tsai and Wu 2015). Potency was determined by estimating the JEV neutralizing antibody response with the plaque reduction neutralization test (PRNT) in BHK cells. Twofold serial dilutions of serum samples ranged from 1:10 to 1:320. CHIKV PRNT was performed using the method of Russell et al., with modifications as previously described (Chusri et al. 2014, Russell et al. 1967). Twofold serial dilutions of serum samples started at 1:2. PRNT data were expressed as the reciprocal of the serum dilution causing 50% of plaque reduction.
Cytokine analysis
Serum samples interleukin 2 (IL-2), IL-4, and interferon γ (IFN-γ) were determined by ELISA (eBioscience, San Diego, CA) at 14 and 35 days postimmunization (dpi), according to manufacturer's instructions.
Lymphocyte proliferation assay
Spleen lymphocytes were isolated using the mice lymphocyte isolation solution kit (Hao Yang Biological Manufacturers, Tian Jin, China), following supplier's instructions at 35 dpi. Lymphocyte concentration was adjusted to 1 × 105/well and proliferation was measured as previously reported (Ren et al. 2014). Each sample was stimulated with 50 μL of inactivated CHIKV or JEV antigen (10 μg/mL) in triplicate. Results were expressed as a stimulation index (SI), which is the ratio of stimulated sample/negative control sample.
Analysis of CD4+ and CD8+ T lymphocytes
Murine spleen cells were isolated using mice lymphocyte isolation solution kit (Hao Yang Biological Manufacturers, Tianjin, China), according to manufacturer's instructions at 35 dpi. Percentages of CD3+CD4+ and CD3+CD8+ T lymphocytes were measured as previously reported (Ren et al. 2014). PE anti-mouse CD8, PE/Cy5 anti-mouse CD3, and FITC anti-mouse CD4 (BioLegend, San Diego, CA) antibodies were used for each sample. The numbers of CD3+CD4+ and CD3+CD8+-positive cells in 10,000 cells were detected by flow cytometry.
CHIKV and JEV challenge in mice
At 35 dpi, mice from groups 1, 3, 5, and 7 were challenged with 1 × 104 TCID50 of JEV. The other groups were challenged with 1 × 104 TCID50 of CHIKV. Mice status and weight were daily monitored. After the experiment, all mice were killed.
Statistical analysis
Statistical analysis and comparisons between immunization groups were performed using the GraphPad Prism software 5.0 (San Diego, CA). The levels of cytokines, SI values, and T cell percentage in different groups were determined by applying one-way repeated-measures analysis of variance. p value was used to describe significant statistical differences. Specifically, p < 0.01, highly significant; p < 0.05, significant; p > 0.05, not significant. Data are presented as the mean ± standard deviation (SD).
Results
Confirmation of the presence of CHIKV E1 and JEV E genes by PCR amplification
rVTT-CE1-JE-EGFP was obtained after 15 rounds of plaque purification (Fig. 1B). BHK cells were infected with recombinant VACV for 48 h, and the virus titer was 6 × 105 TCID50/100 μL (Fig. 1C). To confirm the presence of CHIKV E1 and JEV E genes in rVTT-CE1-JE-EGFP, PCR amplification was carried out using the primers shown in Table 1. A 1323 bp fragment was amplified and was consistent with CHIKV E1 gene size, whereas a 1503 bp fragment was amplified to coincide with the JEV E gene. Results showed that CHIKV E1 and JEV E genes were correctly inserted into rVTT-CE1-JE-EGFP (Fig. 1D).
Analysis of rVTT-CE1-JE-EGFP expression by western blotting
Proteins were obtained from BHK cells infected with rVTT-CE1-JE-EGFP at 0.1 MOI and harvested 48 h postinfection. Western blot results showed one band of ∼60 kDa consistent with JEV E protein molecular weight. The other western blot result showed one band of ∼50 kDa consistent with CHIKV E1 protein molecular weight. These results indicated that CHIKV and JEV E proteins were properly expressed in BHK cells (Fig. 1E).
Specific antibody responses
CHIKV and JEV-specific antibodies by the rVTT-CE1-JE-EGFP were assessed by ELISA at 0, 7, 14, 21, 28, and 35 dpi. Mice in the rVTT-CE1-JE-EGFP group exhibited JEV and CHIKV-specific antibody levels at 7 dpi. CHIKV-specific antibody titers were significantly increased at 21 dpi in mice of the rVTT-CE1-JE-EGFP group, which further increased after boost immunization (21 dpi). At 35 dpi, CHIKV-specific antibody titers in mice of the rVTT-CE1-JE-EGFP group were significantly higher than those of the rVTT-EGFP and PBS control groups (p < 0.01) (Fig. 2A).

Quantifying the serum antigen-specific IgG titer in mice after immunization. CHIKV-E1
Similarly, JEV-specific antibody titers further increased after immunization. At 35 dpi, JEV-specific antibody titers in mice of the rVTT-CE1-JE-EGFP group were significantly higher than those of the rVTT-EGFP and PBS control groups (p < 0.01). Specific antibodies for JEV in rVTT-CE1-JE-EGFP were not significantly higher than those of the SA14-14-2 group at 35 dpi (Fig. 2B).
Neutralizing antibody responses (PRNT)
At 35 dpi, JEV PRNTs for mice in both rVTT-CE1-JE-EGFP and SA14-14-2 groups showed high neutralizing antibodies (1:267 and 1:320), significantly higher than those of mice in the rVTT-EGFP and PBS control groups (Table 2). In addition, CHIKV PRNTs for mice in the rVTT-CE1-JE-EGFP group also showed high neutralizing antibodies (1:64), significantly (p < 0.05) higher than those of mice in the VTT-EGFP and PBS control groups (Table 2).
Neutralizing Antibodies for Japanese Encephalitis Virus or Chikungunya Virus Indicated Day Postimmunization
PRNT, plaque reduction neutralization test.
Cytokine levels in immunized mice
IL-2, IL-4, and IFN-γ were determined to assess cytokine levels change in vaccinated mouse sera at 35 dpi. IFN-γ (Fig. 3A) and IL-4 levels (Fig. 3B) in the rVTT-CE1-JE-EGFP group were significantly (p < 0.01) higher than those of the rVTT-EGFP and PBS control groups at 35 dpi. Moreover, IL-2 levels (Fig. 3C) in the rVTT-CE1-JE-EGFP group were significantly (p < 0.01) higher than those of the rVTT-EGFP and PBS control groups at 35 dpi. At 35 dpi, IFN-γ, IL-4, and IL-2 levels in the rVTT-CE1-JE-EGFP group were, respectively, 1.14, 1.20, and 1.21 times not significantly higher than those in the SA14-14-2 group.

Serum cytokine levels. IFN-γ
T lymphoproliferation
Murine spleen lymphocytes were isolated to determine T lymphocyte proliferation levels at 35 dpi. Results showed significantly (p < 0.01) higher T lymphocyte proliferation levels in the rVTT-CE1-JE-EGFP group following stimulation by inactivated CHIKV antigen, as compared with those of the rVTT-EGFP and PBS control groups (Fig. 4A). T lymphocyte proliferation levels of the rVTT-CE1-JE-EGFP group after stimulation with inactivated JEV antigen were significantly (p < 0.01) higher than those of rVTT-EGFP and PBS control groups, whereas no significant difference was observed as compared with the SA14-14-2 group at 35 dpi (Fig. 4B).
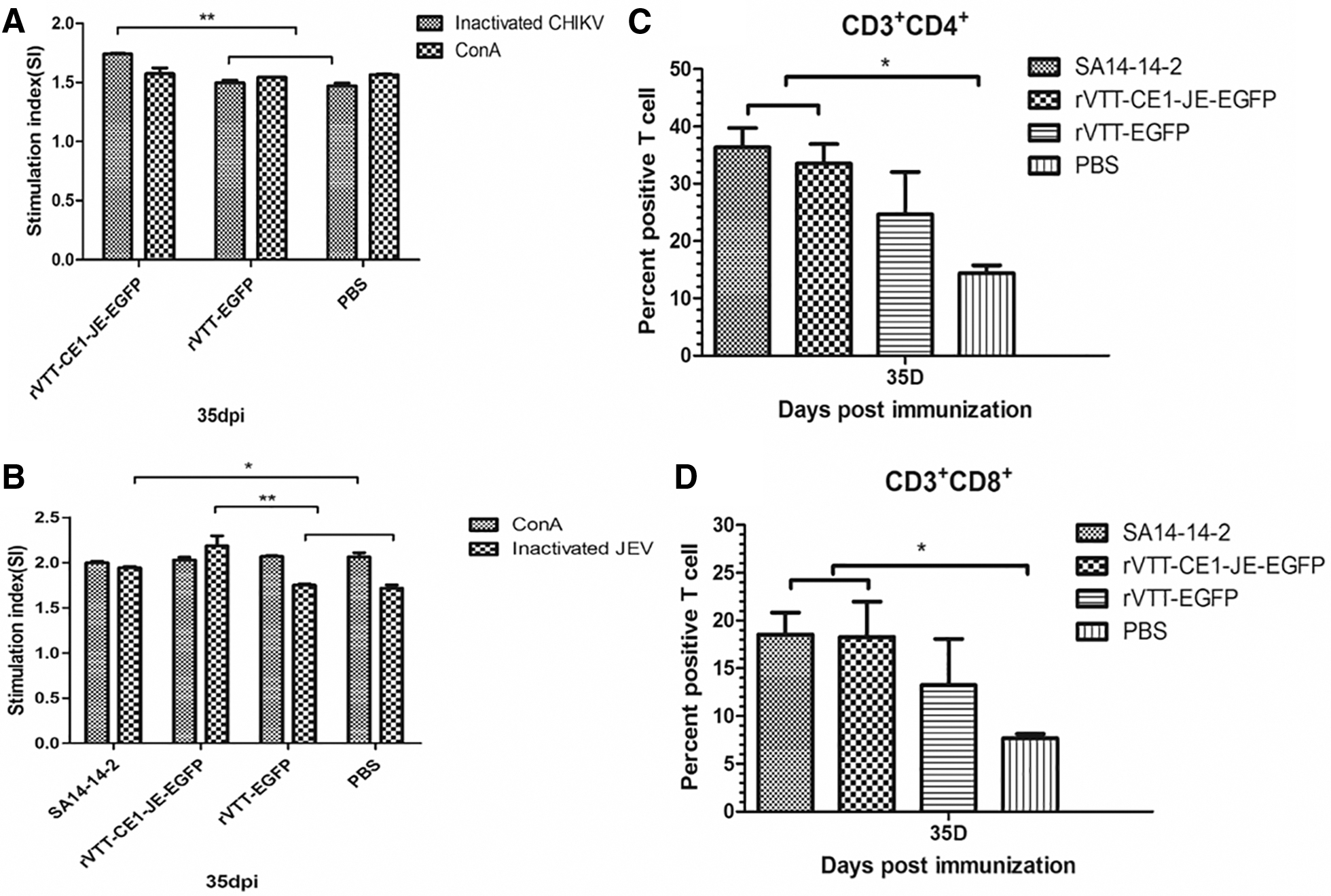
T lymphocyte proliferation and CD3+CD4+ and CD3+CD8+ T lymphocytes percentage in each group. Analysis of T lymphocyte proliferative responses in mice immunized with rVTT-CE1-JE-EGFP, rVTT-EGFP or PBS, and stimulated with inactivated CHIKV
CD4+ and CD8+ T lymphocytes responses
Results of T lymphocytes responses detected by flow cytometry showed that the percentage of CD3+CD4+ (Fig. 4C) and CD3+CD8+ (Fig. 4D) T lymphocytes in the rVTT-CE1-JE-EGFP group were significantly (p < 0.01) higher than those of the PBS control group at 35 dpi. CD3+CD4+ and CD3+CD8+ T lymphocytes percentage in the rVTT-CE1-JE-EGFP group was not significantly higher than that in the rVTT-EGFP control group at 35 dpi.
Body weight change and survival after challenge
To assess the protective effect of rVTT-CE1-JE-EGFP against JEV and CHIKV infection, the status and body weight of mice was daily monitored. Weight of mice challenged with JEV (Fig. 5A) or CHIKV (Fig. 5B) in the rVTT-CE1-JE-EGFP group was not altered. However, mice challenged with JEV or CHIKV in the rVTT-EGFP and PBS groups continuously lost weight. After JEV challenge, survival rate in the rVTT-CE1-JE-EGFP, SA14-14-2, rVTT-EGFP, and PBS groups were 100%, 100%, 33.3%, and 0% at 10 days postchallenge (dpc), respectively (Fig. 5C). Moreover, survival rate of mice challenged with CHIKV in the rVTT-CE1-JE-EGFP, rVTT-EGFP, and PBS groups was 100%, 80%, and 50% respectively, at 10 dpc (Fig. 5D). Results showed that rVTT-CE1-JE-EGFP had a protective effect after JEV and CHIKV challenge.
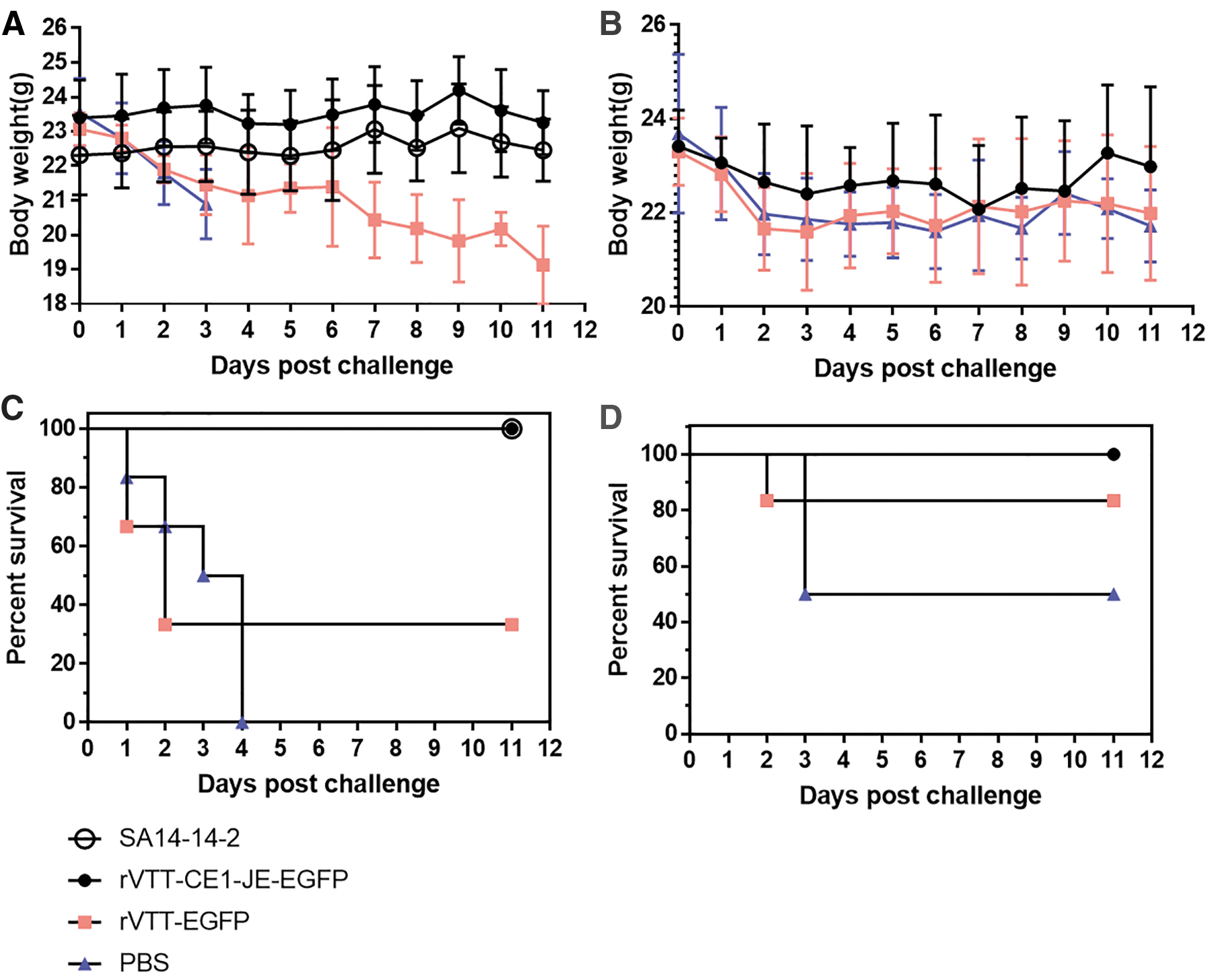
Vaccine protective effect.
Discussion
JEV is a zoonotic disease transmitted to human and swine by mosquitoes (Mansfield et al. 2017, Nickols et al. 2017). WHO 2015 report showed that JEV has spread to humans in 24 countries in the Western Pacific and Southeast Asia (Nickols et al. 2017). There are currently four types of vaccines for JEV, including inactivated mouse brain-derived JEV, inactivated Vero cells-produced JEV, live-attenuated JEV, and live-recombinant vaccines (Nickols et al. 2017). At present, SA14-14-2 live-attenuated vaccine developed by China is one of the most widely used vaccines.
In this study, we constructed a recombinant VACV that expresses JEV E and CHIKV E1 proteins with the VACV TianTan E3L deletion mutant, rVTT-CE1-JE-EGFP. With regard to JEV vaccine, specific antibodies significantly increased, as previously reported (Lee et al. 2016). At 35 dpi, JEV-specific antibodies in the rVTT-CE1-JE-EGFP group were significantly higher than those in the rVTT-EGFP and PBS control groups (p < 0.01), but there was no significant difference as compared with the SA14-14-2 group. At 35 dpi, rVTT-CE1-JE-EGFP PRNT was 1:267, and there was no significant difference compared with that of the SA14-14-2 group. Previous studies reported that immunized mice produced PRNTs similar to the results of this study (Nam et al. 1999), and suggested that PRNTs (1:160) are sufficient to completely protect mice against a JEV challenge (Lee et al. 2016). Sera cytokine levels and CD3+CD4+ and CD3+CD8+ T lymphocytes percentage in the rVTT-CE1-JE-EGFP group were significantly higher than those in the rVTT-EGFP (p < 0.01) and PBS (p < 0.01) control groups, but there was no significant difference compared with those of the SA14-14-2 group, thus indicating that rVTT-CE1-JE-EGFP effectively induced humoral and cellular responses. After murine JEV challenge, body weights in the rVTT-CE1-JE-EGFP group was not significantly altered, and the survival rate was 100%. These results indicate that rVTT-CE1-JE-EGFP was protective against JEV in mice.
At 35 dpi, CHIKV-specific antibodies in the rVTT-CE1-JE-EGFP group were significantly higher than those of the control group (p < 0.01), suggesting that mice produce E1-specific antibodies to provide a protective effect against CHIKV infection. Previous studies have shown recombinant MVA expressing the structural envelope of CHIKV-induced levels of neutralizing antibodies ranging from 40 to 160 at 63 dpi. Animals immunized with recombinant MVA vaccine and challenged with CHIKS27 had null or marginal levels of viral RNA detectable in different tissues (van den Doel et al. 2014). In our study, CHIKV PRNTs in the rVTT-CE1-JE-EGFP group was 1:64 at 35 dpi, which was similar to the recombinant MVA vaccine study.
Some studies have shown that T cells are important effector cells in viral infection. CD4+ and CD8+ play an important role in virus infection in mice (Ng 2017). To date, reports on the role of T cells in CHIKV patients are limited, indicating that CD8+ is predominantly present in the early stages of the disease and that CD4+ appears at later stages of the disease to help produce CHIKV-specific humoral immunity (Wauquier et al. 2011). Some studies have shown that CD4 plays a critical role in joint inflammation (Teo et al. 2015, Teo et al. 2013). In our study, CD3+CD4+ and CD3+CD8+ percentage in the rVTT-CE1-JE-EGFP group was significantly higher than that in the control group (p < 0.05) at 35 dpi. At 35 dpi, CHIKV neutralizing antibody titer was 1:64, which was significantly higher than that of the control group. Neutralizing antibody titers were similar that those of previous studies (Eldi et al. 2017). After CHIKV challenge in mice, weight of mice in rVTT-CE1-JE-EGFP group was not significantly altered, which indicated rVTT-CE1-JE-EGFP induced partial protection. Because BALB/c mice is not very susceptible to CHIKV (5/10 died in PBS group), immunodeficient AG129 mice will be chosen for future CHIKV challenge, which lack type I and II interferon receptor.
Conclusions
JEV and CHIKV may concurrently infect humans by mosquitoes, thus recombinant VACV coexpressing the JEV E and CHIKV E1 was constructed for JEV and CHIKV prevention. These results suggest that recombinant VACV induced humoral and cellular immune responses in mice, and rVTT-CE1-JE-EGFP prevented JEV and CHIKV infection in mice.
Statement of Ethics
All animal protocols were approved by the Animal Care and Use Committee of the Institute of Military Veterinary. Mice used in the experiment were humanely bred and killed at the end of the study.
Footnotes
Acknowledgment
The authors thank the native English-speaking scientists of EditSprings for editing our article.
Author Disclosure Statement
The research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding Information
This work was supported by the National Program on Key Research Project of China (no. 2016YFD0500401), and National Science and Technology Major Project (no. 2018ZX10101003 and no. 2018ZX10102).