
Abstract
Bartonella species are reemerging infectious agents that are transmitted by arthropod vectors among animals and/or humans. At least 13 of the 35 currently recognized Bartonella species are pathogenic for humans. Most of the pathogenic species, except Bartonella quintana and Bartonella bacilliformis, are zoonotic agents with animal reservoirs, including cats, dogs, coyotes, foxes, cattle, and rodents. In this study, a novel Bartonella species was isolated from the blood of a Crocidura suaveolens (Pallas, 1811) Lesser shrew that was captured in the Bartın region of Northwestern Turkey. The strain, RSKK 19006, was characterized using whole-genome sequencing and comparison, multilocus sequence typing (gltA, rpoB, ssrA, nuoG, and 16S rRNA) and internal transcribed spacer sequencing, electron microscopy scanning, biochemical tests, and MALDI-TOF MS (matrix assisted laser desorption ionization-time of flight mass spectrometry). This novel Bartonella is a Gram-negative, rod-shaped, microaerophilic bacterium and has neither flagella nor pilus. As a consequence, we propose to name this new species Bartonella refiksaydamii sp. nov. in Bartonella genus. The zoonotic potential of this novel Bartonella species is as yet unknown.
Introduction
The genus Bartonella includes Gram-negative, fastidious, facultative intracellular bacteria causing bacteremia in humans and/or mammals (Welch 2015). Some Bartonella species are considered reemerging infectious agents transmitted by arthropod vectors among animals and humans. Bartonella species are able to invade erythrocytes and are transmitted among their susceptible hosts by blood-sucking arthropod vectors (Buffet et al. 2013).
In the past 5 years, seven novel Bartonella species were descripted, including Bartonella ancashensis (Mullis et al. 2015), Bartonella apis (Kesnerova et al. 2016), Bartonella fuyuanensis, Bartonella hixiaziensis (Li et al. 2016), Bartonella kosoyii, Bartonella krasnovii (Gutierez et al. 2020), and Bartonella massiliensis (Medkour et al. 2019). In addition, one Candidatus Bartonella sp. as Candidatus “B. hemsudetiensis” (Lilley et al. 2015) was reported. Currently, the Bartonella genus includes 35 validly published species and 3 subspecies (
Depending on the causative species, Bartonella infections in humans may present as cat-scratch disease, bacillary angiomatosis, peliosis hepatis, bacteremia, endocarditis, and neurological and ophthalmic disorders (Celebi 2008, Angelakis et al. 2010).
In 2010, a Bartonella strain, RSKK 19006 ( = CSUR B1084 = RSKK 19006), was isolated from the blood of a Lesser shrew, Crocidura suaveolens (Pallas, 1811). The strain was predicted to be affiliated with members of the genus Bartonella, but distinct from species with a validly published name. In this study, using the polyphasic approach, taxonogenomics (Ramasamy et al. 2014), we propose to a novel Bartonella species that we named Bartonella refiksaydamii sp. nov., with strain RSKK 19006T as type strain.
Materials and Methods
Animal collection
In June 2009, during a field investigation, wild small mammals were collected in the Bartin region (41°41′8″N, 32°13′49″E) in Northwestern Turkey. This region's geographical structure is composed of 46% forest, 35% agricultural, and 7% grassland and pasture. It has an average annual temperature of 12.5°C, humidity 79%, and rainfall 871 mm. Small mammals were trapped using Sherman live-capture traps. Captured live rodents were collected each morning and transported to a biosafety level-2 mobile laboratory, where CO2 euthanasia was performed. They were necropsied on the same day.
The C. suaveolens was identified at the species level using morphological criteria (ventral and dorsal fur color, weight, size of body, head, tail, hind foot, and ear) as previously described (Tez 2000, Krystufek and Vohralik 2001). Animal blood specimens were collected by cardiac puncture, frozen in liquid nitrogen, and transferred to the laboratory, where they were stored at −80°C until further analysis. Permission to conduct this animal study was obtained from the Animal Ethics Committee of the Refik Saydam Hygiene Center under reference no. 06.04.2009/11-27.
Strain isolation and phenotypic description
Blood samples from small mammals were inoculated on 5% horse blood-enriched brain heart infusion agar for Bartonella sp. isolation. The inoculated plates were incubated for 10 days at 37°C in an incubator with 5% CO2. Suspected Bartonella isolates were identified using gltA gene PCR and sequencing. A Bartonella isolate from C. suaveolens blood was not identified at the species level using gltA sequencing (Celebi et al. 2015).
Using La Scola et al.'s (2003) proposal that new Bartonella isolates should be considered belonging to new species if they exhibit 96.0% sequence similarity over a 327-bp gltA fragment, we considered that our isolate, with a gltA sequence similarity lower than 96% with validly published Bartonella species, was a putative new species. To further characterize this novel Bartonella sp., biochemical tests, electron microscopy scanning, matrix assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF MS), multigene sequence analysis (MLST), and whole-genome sequencing were performed as detailed below.
MALDI-TOF MS protein analysis was performed using a Microflex spectrometer (Bruker Daltonics Bremen, Germany) (Seng et al. 2010). Spectra from strain RSSK 19006 were imported into the MALDI BioTyper software (version 2.0; Bruker) and analyzed by standard pattern matching (with default parameter settings). Interpretation of the scores was performed as previously described (Hadjadj et al. 2016). Multilocus sequence typing (MLST) was performed according to protocols from La Scola et al. (2003) gltA, rpoB, internal transcribed spacer (ITS), Diaz et al. (2012) ssrA, and Colborn et al. (2010) nuoG and 16S rRNA using the 27F and 1492R primers (Miller et al. 2013). For DNA sequence analyses, the ExoSAP-IT™ PCR Product Cleanup Reagent kit (ThermoFisher Scientific) was used to purify PCR amplificons according to the kit's procedures. Sequencing was obtained using the ABI 3730XL Sanger sequencer (Applied Biosystems, Foster City, CA) and BigDye Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems). DNA sequences were compared with GenBank using the Basic Local Alignment Search Tool (Blast version 2.0). A 16S rRNA based phylogenetic tree was inferred with MEGA7 using the maximum likelihood and Kimura 2-parameter methods with 1000 bootstrap replicates (Kumar et al. 2016). Different growth temperatures were tested for strain RSKK 19006 (28°C, 37°C, and 42°C) on 5% sheep blood-enriched Columbia agar (BioMérieux, Marcy l'Etoile, France) in a 5% CO2 atmosphere. Gram staining, catalase, and oxidase tests were performed. Biochemical properties such as enzymatic and carbohydrate activity were evaluated using the API ZYM, API 20 NE, and API 50CH strips (BioMérieux). For scanning electronic microscopy, a colony was collected from an agar plate and immersed into a 2.5% glutaraldehyde fixative solution. The slide was gently washed in water; air-dried, and examined on a TM 4000 plus microscope (Hitachi, Tokyo, Japan).
Genomic sequencing and assembly
Genomic DNA of novel B. refiksaydamii sp. strain RSKK 19006 was extracted in two steps: a mechanical treatment was first performed by acid-washed glass beads (G4649-500g; Sigma) using a FastPrep BIO 101 instrument (Qbiogene, Strasbourg, France) at maximum speed (6.5 m/s) for 90 s. Then, after a 2.5-h lysozyme incubation at 37°C, DNA was extracted using an EZ1 biorobot (Qiagen) with the EZ1 DNA Tissue kit. The elution volume was 50 μL. Genomic DNA of strain RSKK 19006 was sequenced using a Miseq sequencer (Illumina) as previously described (Anani et al. 2019b).
The 9,741,472 paired-end reads were filtered according to the read qualities. Reads were quality checked using FastQC and trimmed using Trimmomatic version 0.36.6 (Bogler et al. 2014). MiSeq reads were assembled using the SPAdes version 3.5.0 software (Bankevich et al. 2012). The option “careful” was used to reduce the number of mismatches and short indels. Default parameters were applied for k values, that is, k-mer values of 127, 99, 77, 55, 33, and 21. SSPACE (Boetzer et al. 2011) and GapFiller (Boetzer and Pirovano 2012) were used to combine contigs, using default parameters as previously described (Anani et al. 2019b).
Genome annotation and comparison
Eleven Bartonella genomes were annotated with Prokka 1.13.3 (Seemann 2014) that uses Prodigal 2.6 (Hyatt et al. 2010) for predicting coding sequences, ARAGORN 1.2 (Laslett and Canback 2004), to identify tRNA and tmRNA, whereas rRNA was identified using Barrnap 0.4. The resulting “gff” files were used with Roary (Rapid large-scale prokaryote pan-genome analysis) (Page et al. 2015) with an 80% identity to generate a pan-genome for Bartonella strains, as previously described (Anani et al. 2019a). A core genome-based phylogenetic tree was generated from the core genome alignment using MEGA7 with the maximum-likelihood method and 1000 bootstrap replicates.
The estimation of genomic similarity between the compared strains was obtained using the OrthoANI software (Lee et al. 2016). Furthermore, digital DNA–DNA hybridization (dDDH) values were calculated using Genome-to-Genome Distance Calculator (GGDC) 2.1 on the Type (Strain) Genome Server (TYGS), available at
Results
Strain identification and classification
A novel B. refiksaydamii sp. strain RSKK 19006 was isolated from C. suaveolens blood. The MALDI-TOF MS score obtained for this strain was lower than 1.7, preventing any identification in the Bruker and IHU databases. According to the 16S rRNA sequence-based phylogenetic analysis, this strain was classified within the Bartonella genus, most closely related to Bartonella grahamii with a 99.5% sequence similarity (Fig. 1). Using MLST-based phylogeny, strain RSKK 19006 was closely related to a cluster made of Bartonella elizabethae, Bartonella tribocorum, Bartonella henselae, and Bartonella vinsonii (Fig. 2). The highest gene sequence similarity values between strain RSKK 19006 and other Bartonella species were 95.8% with Bartonella taylorii for gltA, 91% with B. taylorii for ITS, 93.9% with B. taylorii for rpoB, 94.8% with Bartonella washoensis for nuoG, and 96.7% with B. taylorii for ssrA (Table 1).
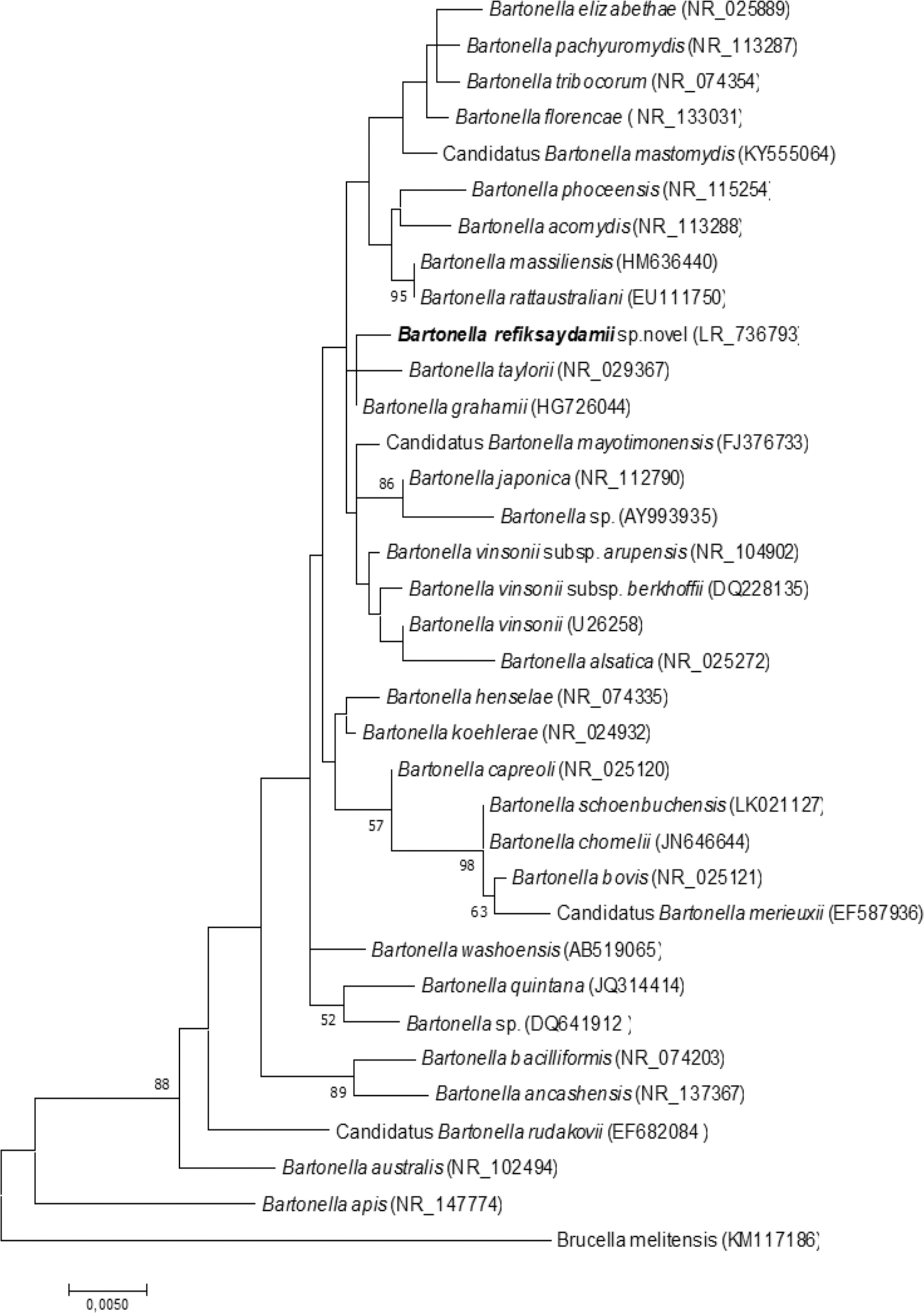
16S rRNA-based phylogenetic tree highlighting the position of Bartonella refiksaydamii sp., nov. (bold), relative to 33 other Bartonella strains. Sequences were aligned using ClustalW. The evolutionary history was inferred using the maximum likelihood method and the Kimura 2-parameter within the MEGA software version 7.0. Codon positions included were first+second+third+noncoding. There were a total of 1335 positions in the final dataset. Brucella melitensis was used as outgroup. Values at the nodes are boostrap replicates obtained by repeating the analysis 1000 times. The scale bar represents 0.5% sequence divergence. GenBank accession numbers are presented in parentheses.

MLST-based phylogenetic tree highlighting the position of Bartonella refiksaydamii sp., nov., (bold) strain RSKK 19006 relative to nine other closely related Bartonella species. Sequences from 16S, rpoB, gltA, nuoG, and ssrA genes and ITS spacer were concatenated and aligned using Muscle v3.8.31 with default parameters, and phylogenetic relationships inferred using the maximum likelihood method and the Kimura 2-parameter within the MEGA software version 7.0., with 1000 bootstrap replicates. There were a total of 3349 positions in the final dataset. ITS, internal transcribed spacer; MLST, multilocus sequence typing.
Highest Sequence Similarity Valued of 16S rRNA, gltA, Internal Transcribed Spacer, rpoB, nuoG, and ssrA Between Strain RSKK 19006 and Other Bartonella Species
Accession numbers for strain Bartonella refiksaydamii sp. novel RSKK 19006.
Accession numbers for each compared species.
Sources of the various compared Bartonella strain.
ITS, internal transcribed spacer.
Furthermore, for the gltA gene and ITS sequences, we observed similarities ranging from 98% to 100% between strain RSSK 19006 and uncharacterized Bartonella sp. reported in some insectivores from England, Sweden, Spain, and Germany (Table 1). These results confirmed the status of new Bartonella species by respecting the thresholds proposed by La Scola et al. (2003).
Phenotypic characteristics
Strain RSKK 19006 grew on 5% sheep blood-enriched Columbia agar at 37°C in microaerophilic conditions. On this medium, colonies exhibited a mix of smooth and rough texture, and were gray and opaque with a diameter of 0.5 to 2 mm. Bacterial cells were rod shaped with a mean length and width of 1.18 ± 0.17 and 0.42 ± 0.04 μm, respectively. Neither flagella nor pili were observed by electron microscopy (Fig. 3). The strain was Gram negative and had neither catalase nor oxidase activity. Using API NE and API 50CH strips, carbohydrate fermentation tests were negative. The API ZYM strip showed positive results for leucine arylamidase and naphthol-AS-BI-phosphohydrolase activities, but negative results for alkaline phosphatase, esterase, esterase lipase, lipase, valine arylamidase, cystine arylamidase, trypsin, α-chymotrypsin, acid phosphatase, α-galactosidase, β-galactosidase, β-glucuronidase, α-glucosidase, β-glucosidase, β glucosaminidase, α-mannosidase, and α-fucosidase activities.

Scanning electron microscopy of Bartonella refiksaydamii sp. novel strain using a Tabletop TM 4000 plus microscope (Hitachi, Tokyo, Japan). The scale bar represents 1 μm.
Genome sequencing and comparison with closely related bacterial strains
The genome size of strain RSKK 19006 was 1,924,864-bp long with a 39.86% G + C content. It was assembled into 113 contigs. Of the 1777 predicted genes, 1735 were protein-coding genes and 42 were RNAs. In silico resistome encompassed genes coding resistance for β-lactamases (Amps). Thirty-two virulence-associated factors were detected from the genome of strain RSKK 19006, of which 22 genes coded type IV secretion system like trwK, trwM, trwL3, trwL7, trwL8, trwG, trwF, trwE, trwD, trwI2, trwJ1, virD4, virB2, virB3, virB4, virB6, virB7, virB8, virB9, virB10, virB11, and others and the rest coded acyl carrier protein (acpXL), surface protein adhesin BadA like, effector protein BepD, and antigen protein virB7.
The genome of strain RSKK 19006 was compared to the available genomes of 10 closely related species. OrthoANI values between strain RSKK 19006 and other Bartonella species ranged from 79.64% to 91.76%. The highest value was obtained with B. taylorii, which is below the 95–96% threshold to delineate a new species (Fig. 4). dDDH values ranged from 21.7% to 43.5%, which is lower than the 70% limit. The highest value, 43.5%, was obtained with B. taylorii (Table 2). The GBDP-based phylogenetic tree also showed that strain RSKK 19006 unambiguously belonged to the Bartonella genus (Fig. 5). Furthermore, the core genome-based phylogenetic tree showed that strain RSKK 19006 was most closely related to B. taylorii (Fig. 6).

Heatmap generated with OrthoANI values calculated using the OAT software between Bartonella refiksaydamii sp. nov., and other closely related taxa with standing in nomenclature.

Phylogenetic tree inferred with FastME 2.1.6.1 from GBDP distances calculated from genome sequences. The branch lengths are scaled in terms of GBDP distance formula d5. The numbers above branches are GBDP pseudo-bootstrap support values >60% from 100 replications, with an average branch support of 54.0%. The tree was rooted at midpoint. GBDP, Genome BLAST Distance Phylogeny.

Core genome-based phylogenetic tree. The Core genome alignment was generated using the Roary software with 80% identity, the tree was inferred with MEGA7 using the maximum likelihood method with 1000 bootstrap replicates. The scale bar represents a divergence of 10%.
Digital DNA–DNA Hybridization Values (%) Resulting from the Pairwise Comparison of the 11 Compared Genomes Using the Genome-to-Genome Distance Calculator Software
dDDH values for novel Bartonella refiksaydamii sp. are shaded in gray.
1: Bartonella alsatica JH725020, 2: Bartonella clarridgeiae ATCC 51734, 3: Bartonella elizabethae F9251 4: Bartonella florencae strain R4, 5: Bartonella grahamii ATCC700132, 6: Bartonella henselae strain Houston 1, 7: Bartonella refiksaydamii sp. nov. RSKK 19006, 8: B. schoenbuchensis CP019789 9: Bartonella taylorii 8TBB, 10: Bartonella tribocorum AM260525, 11: Bartonella vinsonii subsp. berkhoffii CP003124.
Discussion
Bartonella species cause zoonotic diseases in humans, including depending on species cat-scratch disease, chronic bacteremia, Oroya fever, verruga peruana, bacillary angiomatosis, peliosis hepatis, endocarditis, meningoencephalitis, neuroretinitis, and uveitis (Celebi 2008, Welch 2015). They are transmitted among their sensitive hosts through blood-sucking arthropod vectors as flea, louse, and ticks. At least 13 zoonotic Bartonella sp, 6 of which are rodent borne, have been reported. These Bartonella sp. are transmitted to humans by means of infected animal biting, scratching, and their vectors biting (Buffet et al. 2013). The zoonotic properties and precise vectors of the novel B. refiksaydamii sp. that we describe are unknown, so further studies are required.
To date, over 30 validated species have been reported in mammal animals. At least 20 species have been identified in small mammals, notably rodents (Buffet et al. 2013). As Bartonella sp. are fastidious bacteria, growth on blood-based agar may take up to 21 days. Generally, biochemical tests are inert for Bartonella sp., so identification with biochemical tests is not very useful. For these reasons, a few Bartonella sp. were identified before 1990s. Molecular techniques like PCR and DNA sequencing allowed the identification of many novel Bartonella species and subspecies (Celebi 2008, Angelakis et al. 2010, Welch 2015).
In this study, a novel Bartonella species was isolated in the blood of C. suaveolens, which is classified Crocidura genus, Soricidae family in the order Eulipotyphla. C. suaveolens differs from rodents in morphological and karyological characteristics as well as feeding on invertebrates. It lives in close proximity (gardens, vineyards, and seashore) to the human population and is widely distributed from Europe to Asia and Siberia (Dubey et al. 2007). It is the most common Crocidura species in all regions of Turkey (Tez 2000). Although its role as zoonotic agent reservoir is not very well studied, the presence of Hantavirus, Borrelia sp., Listeria monocytogenes, Leptospira sp., Francisella tularensis, and Toxoplasma gondii has been reported in C. suaveolens (Hubálek and Rudolf 2011, Kalmár et al. 2019). However, there was no previous evidence of Bartonella species in this mammal, in contrast with other insectivores from the Soricidae family such as Sorex araneus, Sorex coronatus, Sorex vulgaris, Suncus murinus, Crocidura attenuate, and Crocidura olivieri (Engbank and Lawson 2004, Bai et al.2007, Lin et al. 2012, Halliday et al. 2015, Obiegala et al. 2019, Mediannikov et al. 2013). In France, Bartonella florencae has been isolated in S. araneus’ blood and some Bartonella sp. have been reported from S. araneus and S. coronatus in various European Countries (Holmberg et al. 2003, Bray et al. 2007, Gil et al. 2010, Obiegala et al. 2019, Mediannikov et al. 2013). The insectivores of the Soricidae family are common in Europe and Asia.
Fournier et al. (2009) reported that MALDI-TOF MS is an effective method for identifying Bartonella species. We also used this method for identification of this novel Bartonella strain. When MALDI-TOF MS data were compared with 28 Bartonella species registered in the database, the result of the evaluation was below 1.7 and this result supports the novelty of this Bartonella species.
We sequenced the genome and used it to compare this isolate to species with standing in nomenclature. Whole-genome sequence analysis by OrthoANI, dDDH, and pan-genomic analysis, as well as MLST analysis, demonstrated that strain RSKK 19006 was phylogenetically most closely related to B. taylorii, but belonged to a new species in the Bartonella genus. In contrast, using 16S rRNA-based phylogeny, this novel strain was closely related to B. grahamii (Fig. 1). However, as previously described by La Scola et al. (2003), 16S rRNA sequences lack discriminatory power for Bartonella species and are of limited taxonomic use for these bacteria.
Conclusion
We propose to name this new species B. refiksaydamii sp. nov., belonging to the genus Bartonella, family Bartonellaceae, order Rhizobiales, class Alphaproteobacteria, and phylum Proteobacteria.
Description of B. refiksaydamii sp. nov.
B. refiksaydamii (
B. refiksaydamii sp. nov., is a nonmotile, Gram-negative bacterium. Optimal growth is observed at 37°C in microaerophilic atmosphere. Colonies appear as a mix of smooth and rough, gray, and opaque with a diameter of 0.5 to 2 mm on 5% sheep blood-supplemented Columbia agar. Length and width are 1.18 ± 0.17 and 0.42 ± 0.04 μm, respectively. Cells are rod shaped and have neither flagella nor pili. B. refiksaydamii strain RSKK 19006 sp. nov. exhibited leucine arylamidase and naphthol-AS-BI-phosphohydrolase activities. The genome size and G + C contents are 1.92 Mb and 39.86 mol%, respectively. The type strain (RSKK 19006) was isolated from C. suaveolens, collected in Bartın rural area (41°41′8″N, 32°13′49″E) in Northwest of Turkey. On the basis of phenotypic, phylogenetic, and genomic analyses, we formally propose to name this new species B. refiksaydamii sp. nov. (RSKK 19006), which is the type strain. Genome and gene sequences were deposited in GenBank under the accession numbers, whole-genome CACRUZ010000000, 16S rRNA LR736793, gltA LR736794, ITS LR738423, rpoB LR738912, ssrA LR738913, and nuoG LR738915. This strain is deposited in RSKK and CSUR culture collections as RSKK 19006 and CSUR B1084 in two countries.
Footnotes
Acknowledgments
The authors thank Olivia Ardizzoni for sequencing the genome, Aurelia Caputo for submitting the genomic sequence to GenBank, Hitachi Company, Tokyo, Japan, for scanning electron microscopy, and Turkish Hantavirus field study group for identifying of C. suaveolens.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This study was funded by The Mediterranee Infection Foundation.