
Abstract
The extended distribution and potential introduction of exotic ticks and associated tick-borne pathogens along the northern and southern routes of migratory birds pose zoonotic tick-borne disease risks to wild and domestic animals and incidentally to humans. A knowledge of bird migratory patterns, species of attached ticks, and associated pathogens during their migrations to and from their feeding and nesting grounds is central to understanding associated tick-borne disease risks. Tick-borne disease surveillance was conducted from 2010 to 2011 and 2016 at Hong-do (do = island), Heuksan-do, and Nan-do, major stopovers for migratory birds in Republic of Korea (ROK), as part of the Migratory Birds Research Center bird-banding program for studying bird migration patterns in the ROK. A total of 877 ticks belonging to three genera and nine species were collected, Ixodes turdus (576, 65.7%), Haemaphysalis flava (134, 15.3%), H. longicornis (91, 10.4%), I. nipponensis (56, 6.4%), H. formosensis (7, 0.8%), H. ornithophila (6, 0.7%), H. phasiana (5, 0.6%), H. concinna (1, 0.1%), and Amblyomma testudinarium (1, 0.1%) were collected from 274 birds belonging to 20 genera and 41 species. A total of 15/380 pools (3.95%) were positive for Borrelia species (14 pools of I. turdus and 1 pool of H. flava), while only 1/380 pools (0.26%) was positive for Anaplasma phagocytophilum (1 pool of I. nipponensis). Our findings support the role of migratory birds as possible vectors for the introduction of tick-borne pathogens, which requires continuous monitoring for the potential introduction of ticks and their associated tick-borne pathogens.
Introduction
Hong-do (do = island) and Heuksan-do in Korea are islands near the mainland coastal area of the Republic of Korea (ROK) where many migratory birds stopover on their way to their summer feeding and breeding grounds or remain during their breeding season. Recent surveillance of migratory birds that identified the introduction of associated ectoparasites, for example, mites and ticks, demonstrated the potential for the introduction of not only exotic tick species but also the potential introduction of associated tick-borne pathogens. Various tick-borne pathogens associated with migratory birds (e.g., Anaplasma phagocytophilum, A. bovis, Babesia spp., Borrelia burgdorferi sensu lato, and other unidentified Borrelia spp., Bartonella grahamii, Rickettsia monacensis, R. helvetica, R. sibirica, and Rickettsia spp. [including Candidatus Rickettsia vini]) have been identified (Smith et al. 1996, Ogden et al. 2008, Paulauskas et al. 2009, Hildebrandt et al. 2010, Palomar et al. 2012, Kang et al. 2013).
Anaplasma phagocytophilum, A. bovis, B. turdi, Borrelia spp., and Bartonella grahamii were reported from Haemaphysalis flava, H. longicornis, H. ornithophila, Ixodes turdus, and I. nipponensis, collected from migratory birds transiting Hong-do from 2008 to 2009 (Kang et al. 2013). The Animal and Plant Quarantine Agency (APQA) and Migratory Birds Research Center, in association with the Entomology Section, 65th Medical Brigade, examined ticks collected from migratory birds as part of the migratory bird-banding program in the ROK to understand bird migratory patterns and identify the potential for the introduction of exotic ticks and associated tick-borne pathogens of veterinary and medical importance, and additionally to monitor tick species and associated tick-borne pathogens of migratory birds compared with the previous studies in Hong-do.
Materials and Methods
Survey areas
Hong-do (34°41′ N, 125°11′ E) and Heuksan-do (34°41′ N, 125°25′ E) are remote, protected islands with a total area of 6.4 and 19.7 km2, respectively, and are located in Heuksan-myeon, Sinan-gun, Jeonnam province, 115 and 93 km west of Mokpo, a mainland port city. Hong-do was designated as a National Monument (No. 170) in April 1965. Nan-do (Gauido-ri, Geunheung-myeon, Taean-gun, Chungnam Province; 36°41′ N, 126°6′ E) is a small (0.0476 km2) remote, uninhabited island located 30 km west of Mohang, a major mainland port city, and was designated as a protected National Monument (No. 334) on November 16, 1982 (Fig. 1).
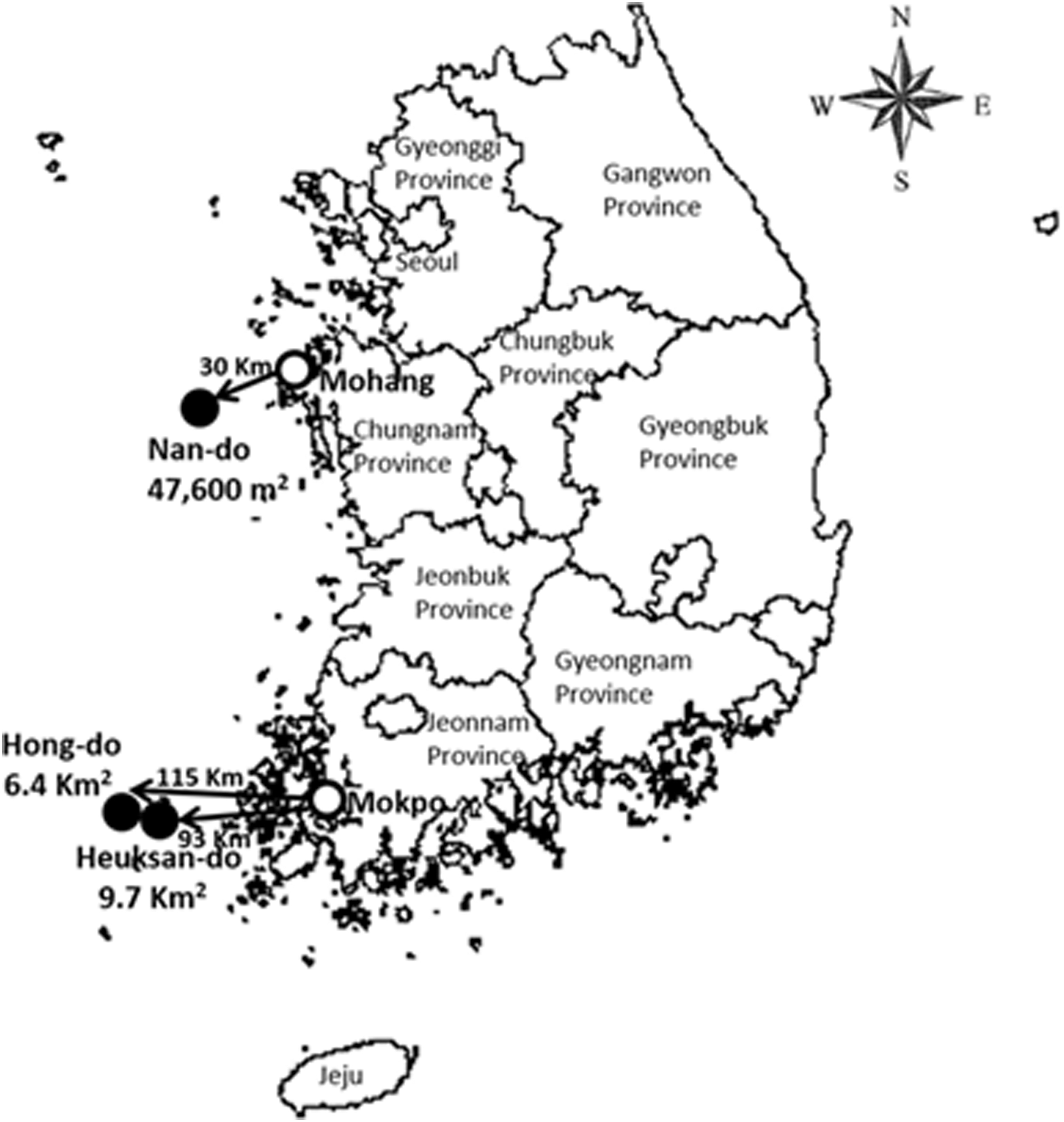
Collection sites (●) of ticks collected from migratory birds during the Migratory Birds Research Center migratory bird-banding surveys during 2010, 2011 and 2016 [mainland port cities (
Bird and tick collections
Migratory birds were captured using 36 mm nylon mist nets (12 meters in length, 2.5 meters high) placed at ground level. Trapped birds that flew into the mist nets were carefully removed to prevent injury and then identified to species, sex, and relative age determined, and then banded with a unique identification number. Before release, the banded birds were closely examined, particularly around the head and neck, for ectoparasites. Ticks were carefully removed by grasping their mouthparts with a fine forceps and gently pulling backward so as not to injure the host. The ticks were then placed individually in cryovials containing 70% ethyl alcohol that was then labeled with a unique identification number that corresponded to the migratory bird collection data. Ticks were sent to the Entomology Section, Force Health Protection and Preventive Medicine, Medical Department Activity-Korea/65th Medical Brigade, Camp Humphreys, Pyeongtaek, ROK, where they were microscopically examined and identified stage of development and species using morphological taxonomic identification keys (Hoogstraal et al. 1968, Yamaguti 1971, Hoogstraal and Wassef 1973). An electronic data sheet that included the unique bird identification number, species, sex, and other pertinent information was prepared, after which most tick specimens were transferred to the Bacterial Disease Division, APQA, Gimcheon, ROK for analysis of selected bacterial pathogens. Bird and tick surveys were conducted as part of the normal work of the Migratory Birds Research Center under the National Park Research Institute on an uninhabited island with access only by government and wildlife capture permits.
DNA extraction and PCR amplification
Ticks were pooled by collection dates, location, host species, and tick species, stage of development, and sex. Adult ticks were tested individually, while nymphs and larvae were pooled in groups of 1–6 and 1–9, respectively, based on collection number. The pooled ticks were homogenized in a tissue grinding tube (SNC, Hanam, South Korea) containing 1 mL phosphate-buffered saline and 2.3 mm stainless-steel beads. Total DNA products were extracted from the homogenized ticks using Maxwell® RSC Viral Total Nucleic Acid Purification Kits (Promega, Madison, WI) according to the manufacturer's instructions and then stored at −80°C until used.
PCR was performed using specific primers for two specific tick-borne pathogens (Anaplasma phagocytophilum and Ehrlichia chaffeensis) and two genera (Borrelia and Bartonella spp.) (Table 1). Target DNAs were assayed by PCR for the 16S rRNA (A. phagocytophilum, E. chaffeensis, and Borrelia spp.), groEL gene (A. phagocytophilum and Borrelia spp.), DNA gyrase subunit B gene (Borrelia spp.), and Internal Transcribed spacer region (Bartonella spp.), as described previously (Anderson et al. 1992, Barlough et al. 1996, Lotric-Furlan et al. 1998, Hancock et al. 2001, Park et al. 2004, Schwan et al. 2005, Seki et al. 2006, Zhai et al. 2017). Target DNAs were amplified in 20 μL of AccuPower PCR PreMix (Bioneer, Daejeon, South Korea) containing 2 μL of template DNA and 1 μL of 10 pmol of each primer added to the PreMix tube, and adjusted to a volume of 20 μL with distilled water. PCR reactions were performed at 35 cycles (30 s at 95°C, 30 s at annealing temperature, 30 s at 72°C), followed by 5 min extension at 72°C (Table 1).
Primer Sequences And PCR Conditions Used For The Detection Of Selected Tick-Borne Pathogens
Primers and probes that targeted the 23S rRNA gene of Borrelia burgdorferi were used as reported previously (Hersh et al. 2014) (Table 1). The 20 μL PCR mixture consisted of 10 μL of iQ Supermix (Bio-Rad), 300 nM each of the primer and reverse primer, 200 nM probe, 7 μL of distilled water, and 2 μL of extracted DNA. Amplification and detection were performed with a Bio-Rad CFX connect Real-Time system according to the following protocol: 95°C for 3 min, followed by 40 cycles of 95°C for 15 s and 55°C for 30 s.
Sequencing and phylogenetic analysis
PCR-positive samples of tick-borne pathogens were purified using a QIA Quick Purification Kit (Qiagen). Sequencing of PCR products was conducted by Macrogen (Seoul, ROK). The similarities of obtained sequenced DNAs were analyzed using BLASTn tool of National Center for Biotechnology Information GenBank database. The sequences aligned using the Clustal W method in MegAlign version 7.1 (DNA-STAR, Madison, WI) and phylogenetic trees were generated using neighbor-joining algorithms and the Jukes and Cantor matrix. Support for topology was calculated using 1000 bootstrap replications.
Results
A total of 13,793 birds representing >150 species were banded and examined for ectoparasites during 2010, 2011, and 2016. A total of 877 ticks belonging to 9 species and 3 genera were collected from 274 (2.0%) birds belonging to 41 species and 20 genera (Tables 2 and 3). Emberiza spodocephala (24.2%), Tarsiger cyanurus (8.9%), Emberiza elegans (7.9%), Turdus pallidus (7.0%), and Emberiza rutile (6.6%) were the most frequently captured migratory birds (Table 2). The majority of the 877 ticks were collected from the Yellow-throated Bunting (E. elegans) (180, 20.5%), followed by the Pale Thrush (T. pallidus) (158, 18.0%), the Tristram's Bunting (E. tristrami) (136, 15.5%), the White's Thrush (Zoothera aurea) (88, 10.0%), the Black-faced Bunting (E. spodocephala) (63, 7.2%), the Yellow-browed Bunting (E. chrysophrys) (33, 3.8%), the Chestnut Bunting (E. rutile) (28, 3.2%), the Dusky Thrush (T. eunomus) (23, 2.6%), the Mugimaki Flycatcher (Ficedula mugimaki) (21, 2.4%), and the Red-flanked Bluetail (Tarsiger cyanurus) (21, 2.4%). The remaining 31 species accounted for 14.4% (ranging 1–16 ticks, 0.1–1.8%) of the 877 ticks collected (Table 2). The highest migratory bird tick infestation rates were observed for Carpodacus sibiricus (50.0%), Locustella davidi (50.0%), Phylloscopus collybita (50.0%), L. pleskei (16.7%), Z. aurea (15.9%), T. cardis (11.1%), Pycnocotus sinensis (10.0%), Calliope calliope (8.6%), Luscinia svecica (8.3%), and Geokichla sibirica (7.7%).
Detection of Borrelia and Anaplasma spp. from Ticks Collected from Migratory Birds During Bird-Banding Surveys on Hong-Do and Heuksan-Do, Jeollanam Province, and Nan-Do, Chungnam Province, Republic of Korea, During 2010–2011, and 2016
Not tested: Haemaphysalis ornithophila (six ticks) and H. concinna (one tick) were mounted on slides for identification.
Number of Ticks, by Species, Collected from Migratory Birds, by Species, During 2010–2011, and 2016 Migratory Bird-Banding Surveys Conducted by the Migratory Birds Research Center on Hong-Do and Heuksan-Do, Jeollanam Province, and Nan-Do, Chungnam Province, Republic of Korea
Number of birds captured.
Number of birds with ticks.
IT, Ixodes turdus; IN, Ixodes nipponensis; HFL, Haemaphysalis flava; HL, Haemaphysalis longicornis; HFO, Haemaphysalis formosensis; HO, Haemaphysalis ornithophila; HC, Haemaphysalis concinna; HP, Haemaphysalis phasiana, AT, Amblyomma testudinarium; L, larva; N, nymph; M, male; F, female.
A total of nine species (larvae, nymphs, and/or adults) belonging to three genera, were identified. Ixodes turdus (576, 65.7%) was the most commonly collected tick, followed by H. flava (134, 15.3%), H. longicornis (91, 10.4%), I. nipponensis (56, 6.4%), H. formosensis (7, 0.8%), H. ornithophila (6, 0.7%), H. phasiana (5, 0.6%), H. concinna (1, 0.1%), and A. testudinarium (1, 0.1%) (Table 3). Two species, H. ornithophila (six ticks) and H. concinna (one tick) were not tested for pathogens since they were mounted on slides for identification and preservation.
A total of 15/380 pools (3.95%) were positive for Borrelia species (14 pools of I. turdus, and 1 pool of H. flava), while only 1/380 pool (0.26%) of I. nipponensis was positive for A. phagocytophilum. All positive samples of Borrelia spp. and Anaplasma phagocytophilum were detected by PCR for 16S rRNA and groEL gene. Neither E. chaffeensis nor Bartonella spp. was detected. Anaplasma phagocytophilum, Borrelia spp., E. chaffeensis, and Bartonella spp. were not detected in pools of H. formosensis, H. longicornis, H. phasiana, H. concinna, H. ornithophila, or Amblyomma testudinarium collected from migratory birds.
Borrelia spp. were detected in ticks collected from E. elegans, E. spodocephala, E. tristrami, T. pallidus, Horornis borealis, Z. aurea, and Phylloscopus collybita. Among the 380 tick pools, 2.1% of the larvae, 1.6% of the nymphs, and 0.3% of adult female were positive for Borrelia spp. A total of 12/15 Borrelia-positive pools (80.0%) were from ticks collected from birds captured at Heuksan-do (HS) (3 pools from E. elegans, 5 pools from T. pallidus, 1 pool from Phylloscopus collybita, 1 pool from E. spodocephala, 1 pool from E. tristrami, and 1 pool from Cettia diphone borealis), while the remaining 3 Borrelia-positive pools (20.0%) were from birds captured at Hong-do (H) (2 pools from Z. aurea and 1 pool from T. pallidus). Anaplasma phagocytophilum was only collected from one I. nipponensis larval tick (0.4% of all larval ticks) collected from E. rutile captured at Heuksan-do.
Analyses of tick-borne pathogen DNA sequences
16S rRNA PCR products were used for phylogenetic analysis of A. phagocytophilum collected from migratory bird ticks, and was 100% identical to previous isolates reported in the ROK and other countries, for example, Russia, Slovenia and China, indicating the same genetic lineage (Fig. 2A). Anaplasma phagocytophilum groEL PCR products in migratory bird ticks were 100% identical to previous isolates reported in the ROK and other countries, for example, Germany and Sweden, indicating the same genetic lineage (Fig. 2B).
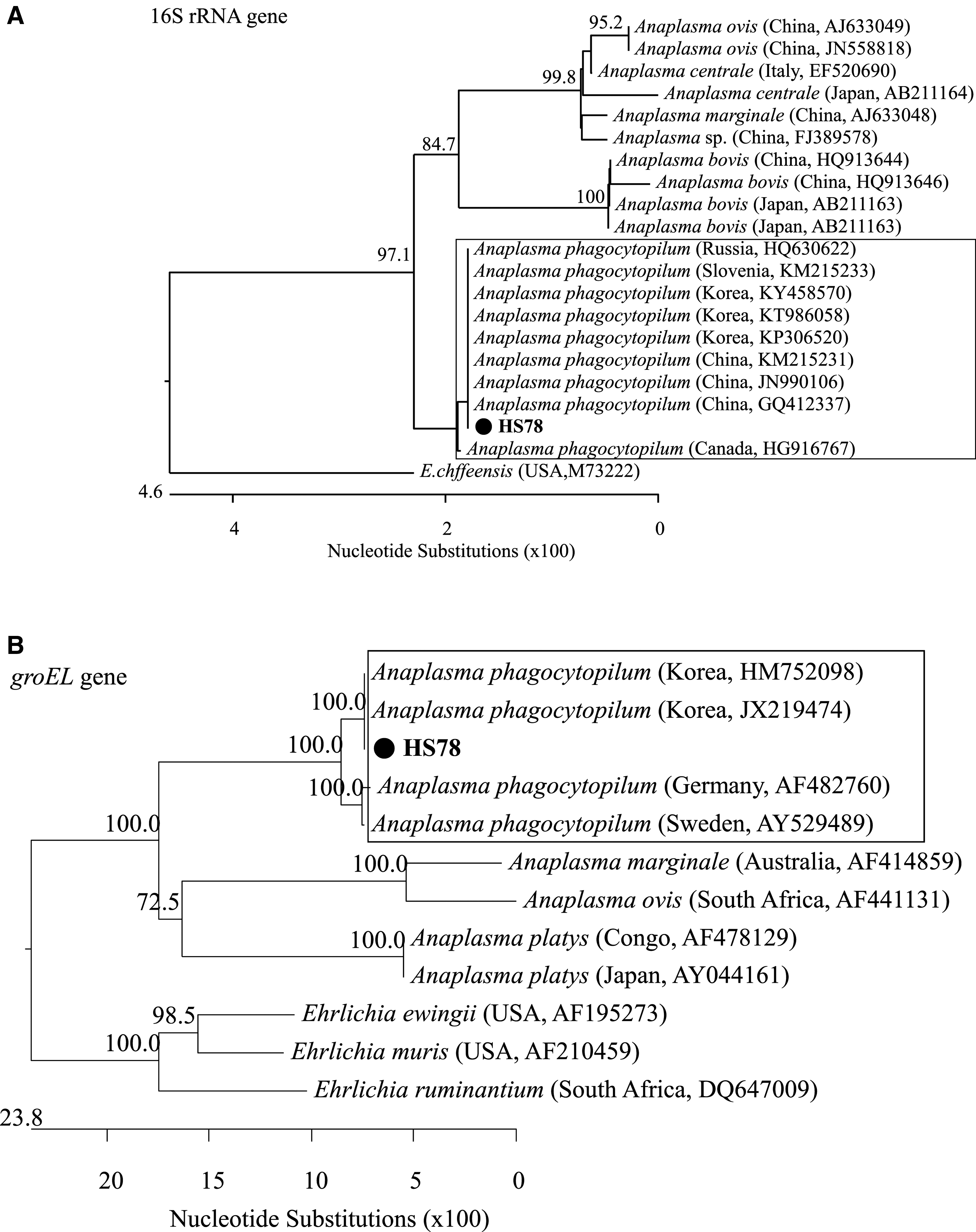
Phylogenetic tree showing relationship between Anaplasma phagocytophilum detected in ticks collected from migratory birds in the Republic of Korea and Anaplasma and Ehrlichia species based on partial sequences of the 511 bp 16S rRNA
Phylogenetic analysis of Borrelia spp., based on sequence analysis of 16S rRNA PCR products detected in migratory bird ticks, was divided into three groups (Fig. 3A). Group I (H100, HS25, HS22, HS183, HS49, HS24, and HS23) demonstrated a high degree of genetic similarity between B. bavariensis reported from Germany, B. burgdorferi, B. kurtenbachii, B. californiensis, B. americana, and B. mayonii reported from United States of America, B. tanukii and B. garinii reported from Japan, B. lusitaniae reported from Portugal, and B. sinica reported from China. Group II (HS90, H45, HS98, HS2, HS173, HS18, and H51) demonstrated a high genetic similarity with B. turdi derived from Japan, and B. lanei derived from United States of America. Group III (H136) did not have a high genetic match, but was most closely related to B. theileri reported from France, B. miyamotoi reported from Japan, B. coriaceae and B. anserine derived from United States of America, and B. hispanica derived from Moroco. All three genotypes of Borrelia spp. were identified from ticks collected from migratory birds captured at Heuksan-do, while only two genotypes of Borrelia spp. was identified from ticks from Hong-do. Borrelia spp. belonging to Group III (H136) were detected in H. flava, which was different from Borrelia spp. classified as Group I and II detected in I. turdus. Borrelia spp. detected in ticks collected in March and November corresponded to Group II, while Borrelia spp. detected in ticks collected in April corresponded to Group I. However, ticks collected from migratory birds at Hong-do were positive for both Groups I and II genotypes during April. The Group III (H136) genotype was only detected in a tick collected from a migratory bird (Emberiza elegans) in November at Heuksan-do.
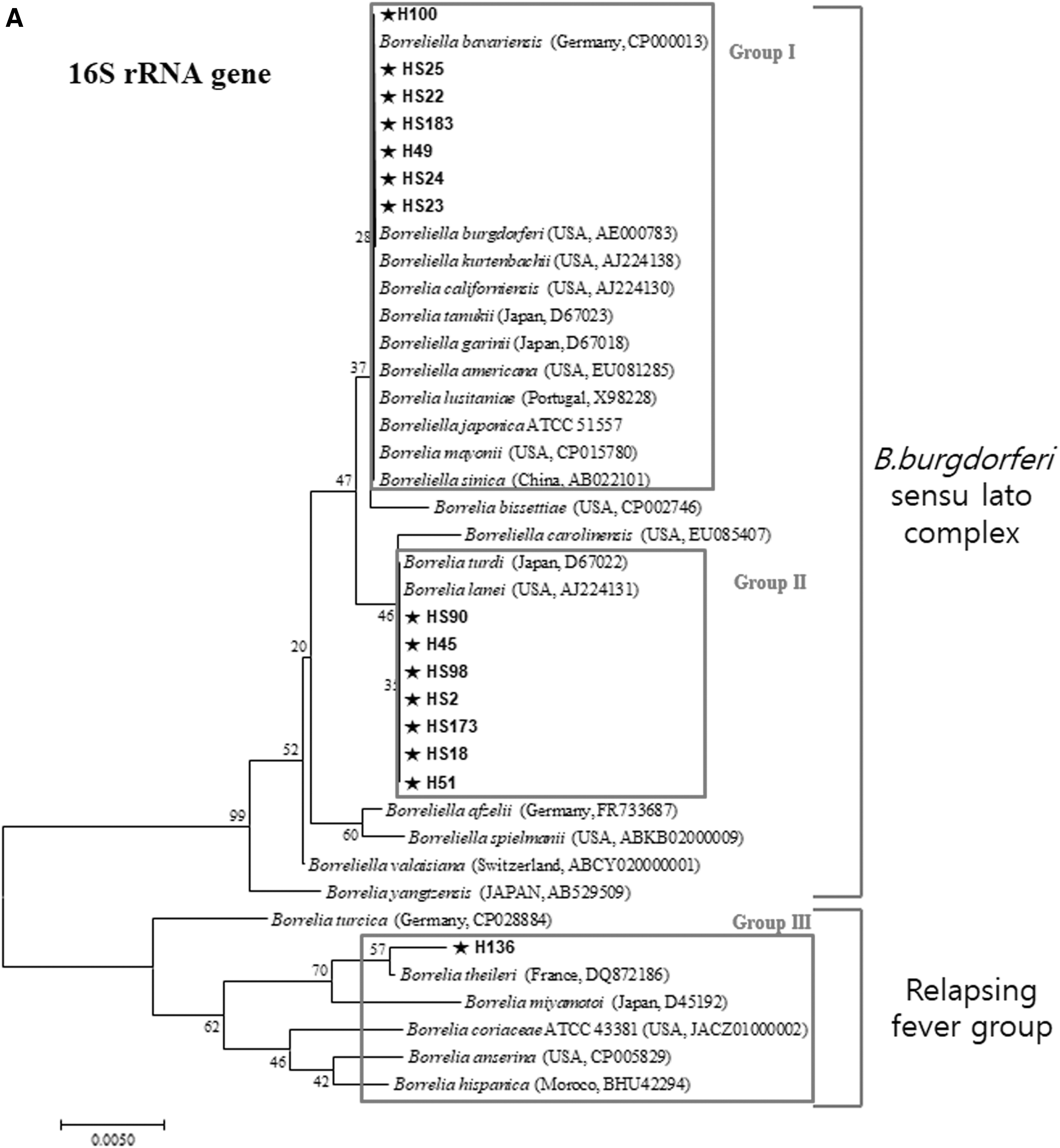
Phylogenetic tree showing relationship between Borrelia spp. detected in ticks collected from migratory birds in the Republic of Korea and Borrelia spp. based on partial sequences of the 622 bp (or 714) 16S rRNA
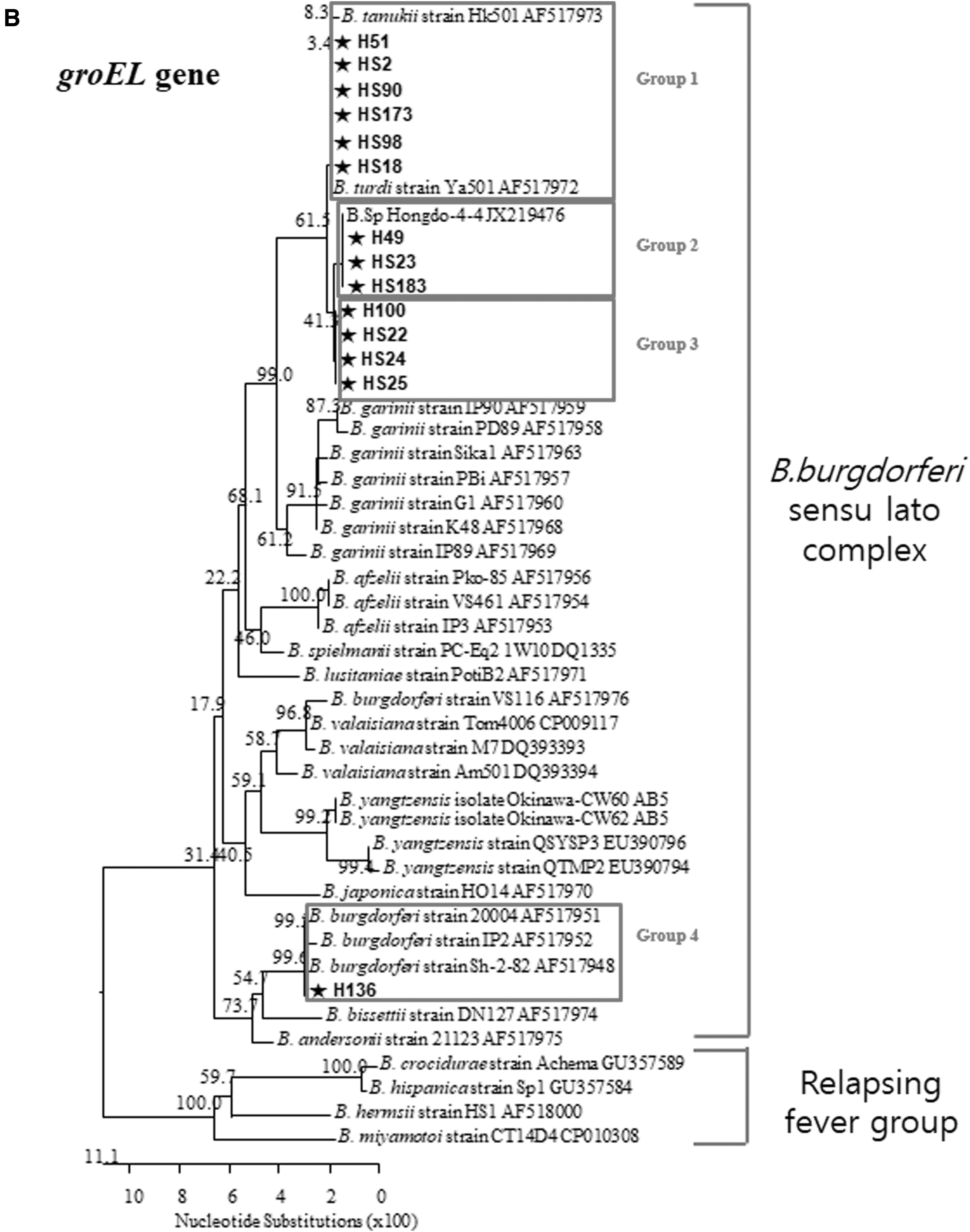
Based on the phylogenetic analysis of groEL PCR products, Borrelia spp. were divided into four groups (Fig. 3B). While pool H45 was positive for the 16S rRNA gene fragment, it was not positive using the groEL gene fragments. Group I 16S rRNA-positive ticks were subdivided into two subtypes using the groEL gene. Group II positive ticks using the 16S rRNA gene fragments (H51, HS2, HS90, HS173, HS98, and HS18) were placed in the same Group using the groEL gene. Group I 16S rRNA gene fragments were subdivided into subgroups 2 (H49, HS23, and HS183) and 3 (H100, HS22, HS24, and HS25) using groEL gene fragments. Phylogenetic analysis of 16S rRNA gene placed pool H136 in Group III, while it was classified Group 4 genotype using the groEL gene. Interestingly, H136 grouped with the relapsing fever group when analyzed using the 16S rRNA gene, but in the groEL gene analysis it belonged to B. burgdorferi sensu lato complex, similar to other Borrelia spp. observed in other migratory bird ticks.
Discussion
Migratory birds transport ticks during their annual migrations across international borders and continents. Therefore, intrinsic pathogen screening of migratory bird ticks in the ROK is important for preemptive detection of tick-borne diseases. In this study, ticks were collected from migratory birds captured in transit to Heuksan-do, Hong-do, and Nan-do that are migratory bird mid-resting areas. Summer migratory birds transit Korea during the spring from their feeding grounds in Southeast Asia to their breeding grounds in northern China and Russia, and again transiting Korea on their return to their feeding grounds in Southeast Asia. However, some migratory birds stay in Korea during the entire summer, while others may remain the entire year. Two agents (A. phagocytophilum and E. chaffeensis) detected in migratory bird ticks are known human tick-borne pathogens, while some species of Borrelia and Bartonella detected in migratory bird ticks may be of both veterinary and medical importance.
Species of migratory birds vary greatly by country and season. There were no common migratory bird species transiting the ROK associated with those captured in Canada and Germany (Ogden et al. 2008, Hildebrandt et al. 2010). Only 1/15 (6.7%) species of birds captured in Lithuania and 3/27 (11.1%) species captured in Norway were commonly collected in the ROK (Paulauskas et al. 2009). Of the four species of migratory birds captured in Hokkaido, Japan, during 1995, only two were commonly collected in the ROK during 2010, 2011, and 2016. Due to the proximity of Japan, there are many similarities in relative proportions of migratory bird species captured in the ROK (Miyamoto et al. 1997). A total of 21 species of migratory birds were captured at Hong-do from 2008 to 2009, of which 12 species (57.1%) were commonly collected (Kang et al. 2013). An additional 20 species of migratory bird species were captured, of which 17/41 (41.5%) species were commonly collected during the 2010, 2011, and 2016 surveys. The reasons for these differences are uncertain, but are likely due to the wider area collected, three islands versus one island.
Tick infestation rates of migratory birds in this study (2.0%) were similar to that reported in 2008–2009 (3.0%). The highest infestation rates (50.0%) were shown in the Long-tailed Rosenfinch (Carpodacus sibiricus), Baikal bush Warbler (Locustella davidi), and Common Chiffchaff (Phylloscopus collybita), while the lowest infestation rates (0.2%) were observed in the Eurasian Siskin (Spinus spinus) and Stejneger's Stonechat (Saxicola stejnegeri) (Table 2). When comparing migratory species with high tick infestation rates in each country, such as Canada, Lithuania, and Norway (Ogden et al. 2008, Paulauskas et al. 2009), common tick species were not identified, and there were no specific migratory species with high rates of tick-associated infections. Interestingly, only one Gray-backed Thrush (T. hortulorum) showed consistent high rates of tick infestation compared to migratory bird tick infestation rates in previous Hong-do migratory bird surveillance (Kang et al. 2013).
In this study, the collection rate of H. longicornis from migratory birds was about twice as high (10.4%) as previously reported (5.7%), suggesting that the risk of spreading associated tick-borne pathogens by this tick is potentially increasing (Kang et al. 2013). Black-tailed Gull (Larus crassirostris), which is usually distributed at Southeast China, Korea, Japan, and Far East Russia, was ever found in West Alaska and Mexico (Heinl 1997, Garrett et al. 1998, Kim et al. 2018). Recently, H. longicornis was first detected in United States of America on a sheep in 2017 (Beard et al. 2018, Rainey et al. 2018, Egizi et al. 2019, Saleh et al. 2019, Duncan et al. 2020). Based on Black-tailed Gull migration routes, it could play a role to introduce H. longicornis to United States of America from Asia. Therefore, One Health approach will be needed to prevent the spread of ticks and tick-borne pathogens internationally. Haemaphysalis formosensis, H. phasiana, H. concinna, and A. testudinarium were not collected in previous migratory tick surveillance in the ROK (Kang et al. 2013). While the numbers of these species were relatively low, there is an increased risk for the introduction of species-specific associated pathogens, as well as the introduction of these ticks to islands and the mainland of the ROK, where migratory birds stopover during their transit to their breeding grounds. The ticks collected from migratory birds on the three islands were distinct species from those of Japan, Spain, Germany, Lithuania, Norway, and Canada (Miyamoto et al. 1993, 1997, Ogden et al. 2008, Hasle et al. 2009, Paulauskas et al. 2009, Hildebrandt et al. 2010, Palomar et al. 2012). For example, while I. ricinus is pandemic in Europe, it was not collected from migratory birds transiting the ROK. In addition, Haemaphysalis spp. was collected from migratory birds in the ROK, but was not commonly collected in Europe.
Among the Korean migratory bird ticks, the tick species that was positive for A. phagocytophilum was I. nipponensis, while in Europe, A. phagocytophilum was detected in I. ricinus (Paulauskas et al. 2009, Hildebrandt et al. 2010, Palomar et al. 2012), and in Canada, it was detected in I. scapularis and I. dentatus (Ogden et al. 2008). The rates of A. phagocytophilum infection in migratory bird ticks in all countries were relatively low (0.3%), except Norway where rates were 2.6% (Hasle et al. 2009).
The geographical distribution of Borrelia spp. detected in ticks varied among species collected in each of the countries, for example, I. nipponensis, and I. turdus in the ROK (Kang et al. 2013), I. ricinus, I. frontalis, and H. punctate in Spain (Palomar et al. 2012), I. persulcatus in Japan (Miyamoto et al. 1997, Iwabu-Itoh et al. 2017), and I. scapularis and I. dentatus in Canada (Ogden et al. 2008). The proportion of ticks positive for B. burgdorferi that were collected from birds in Canada was very high in I. scapularis (12.2%), while in other countries, the infection rate was relatively low ranging from 0.3% to 3.7%. In addition, Borrelia spp. was detected for the first time in H. flava collected from migratory birds in the ROK in this study. Bartonella spp. was detected in I. turdus (0.9%, 1/108) collected from a migratory bird captured in 2008–2009 at Hong-do (Kang et al. 2013). However, Bartonella spp. was not detected from the migratory bird ticks during 2010, 2011, and 2016 in this study.
Genetic analysis of A. phagocytophilum detected in ticks collected from migratory birds in two regions (Jomfruland and Lista) of Norway were similarly detected in Common Blackbird (Turdus merula), Song Thrush (T. philomelos), and Common Starling (Sturnus vulgaris) (Paulauskas et al. 2009). In Europe, T. merula was the predominant migratory species, in which A. phagocytophilum was detected. Borrelia spp. and A. phagocytophilum in migratory bird ticks may be transmitted to small rodents at migratory bird stopovers and as they transit land masses/islands on their way to feeding and breeding grounds, resulting in the potential for establishing exotic zoonotic infectious diseases. Continuous monitoring of migratory bird ticks and their associated pathogens is required for the early detection of imported nonendemic tick-borne pathogens.
The sequence of A. phagocytophilum in the migratory bird ticks (HS78) is in complete agreement with that of A. phagocytophilum derived from small mammals (KY458570) and humans (KP306520), so migratory birds likely play a role in A. phagocytophilum transmission (Fig. 2). These results are consistent with previous surveys (Vorou et al. 2007, Kang et al. 2013). The sequence of A. phagocytophilum detected in I. nipponensis collected from migratory birds was confirmed to be a genetic lineage similar to A. phagocytophilum detected in other ticks, voles, raccoon dogs, water deer, horses, and humans (Fig. 2). This suggests the potential transmission of A. phagocytophilum by migratory bird ticks to zoonotic hosts and incidentally to domestic animals, birds, and humans.
Borrelia spp. (HS136) was detected for the first time from H. flava collected from migratory birds in the ROK in this study. However, there were many genetic differences in phylogenetic analysis of the 16S rRNA and groEL genes, compared with Borrelia spp. from I. turdus (Fig. 3). This implies that these new genetic variants most likely originated from other countries and may become established in the ROK. Haemaphysalis flava (HS136) positive for Borrelia spp. was collected in November and the observed genetic differences are likely due to the host's northernmost breeding ground.
These results provide an insight into the potential role of migratory birds in the introduction and dispersal of ticks and associated tick-borne pathogens throughout their range in Asia. Furthermore, long-term tick-borne disease surveillance is necessary to better understand the potential dynamics of tick-borne disease transmission in the ROK.
Footnotes
Acknowledgments
We thank the many employees at the Migratory Birds Research Center, National Park Research Institute, Korea National Park Service, on Heuksan Island, Jeollanam Province, ROK, for collecting nest soil and litter during seabird conservation and breeding surveys and birds captured in mist nets during the migratory banding survey. We also thank Jiyeon Lim at Animal and Plant Quarantine Agency (APQA) for genetic analysis of tick pathogens. Funding for the “2016 Bird Survey and Research” was provided by the Korea National Park Research Institute and funding for molecular detection of the pathogens by a R&D Grant for Quarantine Technology of Agriculture, Rural Affairs, and Livestock from APQA in the Republic of Korea. Partial funding also was provided by the Armed Forces Health Surveillance Branch-Global Emerging Infections Surveillance and Response System (AFHSB-GEIS), Silver Spring, MD, and the 65th Medical Brigade, Seoul, Korea (ProMIS ID #P0025-2016-2018-ME).
Disclaimer
The opinions expressed herein are those of the authors and are not to be construed as official or reflecting the views of the United States Departments of the Army or Defense.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This work was supported by a grant from Animal and Plant Quarantine Agency in the Republic of Korea (Project No. B-1543081-2020-22-03), and by the Armed Forces Health Surveillance Branch-Global Emerging Infections Surveillance and Response System, Silver Spring, MD, USA and the 65th Medical Brigade, Seoul, Korea (ProMIS ID #P0025-2016-2018-ME).