
Abstract
Background:
Severe fever with thrombocytopenia syndrome (SFTS) is an emerging tick-borne zoonosis in China, the Republic of Korea (ROK), and Japan. The presence of the SFTS virus (SFTSV) in companion, livestock, and wild animals has been reported. Recently, human SFTS-like clinical symptoms in cats and cheetahs have been reported in Japan. Therefore, the prevalence of the SFTSV gene or antibody in cats is important for public health as well as veterinary medicine.
Materials and Methods:
Sera were collected from 201 feral and house cats in the ROK in 2017. Samples were analyzed for the presence of the SFTSV gene after RT-nested PCR amplification and for anti-SFTSV antibodies after enzyme linked immunosorbent assay.
Results:
Eight (4.0%) and nine (4.5%) of 201 cat sera were found to be positive for the SFTSV gene and anti-SFTSV nucleocapsid protein antibodies, respectively. Specifically, 5.9% feral and 2.0% house cats were positive for the SFTSV gene, and 6.9% feral and 2.0% house cats were positive for anti-SFTSV antibodies. All sequences of the SFTSV S segment obtained were included in Japanese/Korean SFTSV clades, as opposed to the Chinese clade.
Conclusions:
This study constitutes the first serological study of SFTSV in house and feral cats in the ROK. Evidence of SFTSV in companion animals indicates that SFTSV can circulate in homes and that more intensive precautions and education measures are needed for companion animal guardians and veterinarians.
Introduction
Severe fever with thrombocytopenia syndrome (SFTS) is an emerging zoonotic disease caused by Huaiyangshan banyangvirus (formerly SFTS virus, SFTSV) belonging to the Phenuiviridae family of Bunyavirales. This virus is mainly found in China, Japan, and the Republic of Korea (ROK) (Yu et al. 2011, Kim et al. 2013, Takahashi et al. 2014, ICTV 2018). Haemaphysalis longicornis is the main vector of SFTSV (Zhuang et al. 2018). However, the definitive host of SFTSV has not been confirmed. Many studies have been conducted to assess the presence of SFTSV in various companion, livestock, and wild animals and have isolated SFTSV from dogs, sheep, cattle, rodents, and cheetahs (Niu et al. 2013, Ni et al. 2015, Matsuno et al. 2018, Kang et al. 2019). The common clinical signs of SFTS are characterized by high fever, myalgia, anorexia, lethargy, thrombocytopenia, and leukopenia (Yu et al. 2011). In animals, only two cases of SFTS-like clinical symptoms have been reported in cats and cheetahs in Japan (Matsuno et al. 2018, Matsuu et al. 2019).
Although the main mode of SFTSV transmission is through tick bites, some studies have reported that human-to-human transmission or animal-to-human transmission could occur in certain environments, such as during close contact (Jung et al. 2019, Kida et al. 2019). These findings indicate that some dogs and cats, which inhabit the same space and are in close contact with humans, could transmit SFTSV to humans or other animals. Several studies have reported SFTSV infections in cats in the ROK and Japan (Hwang et al. 2017, Lee et al. 2017, Matsuu et al. 2019). However, the seroprevalence of SFTSV in cats has not previously been reported. In this study, we investigated the prevalence of SFTSV and anti-SFTSV antibodies in sera from house and feral cats.
Materials and Methods
Sample collection and ethics statement
All samples used in this study were obtained from two local animal hospitals in Seoul between May and October 2017. A total of 201 sera were collected from 101 feral cats (50 males and 51 females; average age = 2.31 ± 0.89 years) and 100 house cats (62 males, 38 females; average age = 3.48 ± 3.40 years) (Table 1). Sera were separated from whole blood through centrifugation and stored at −80°C until use. RNA was extracted from sera using a Viral Gene-spin™ Viral DNA/RNA Extraction Kit (iNtRON Biotechnology, Gyeonggi, ROK) according to the manufacturer's instructions. This study was approved by the Institutional Biosafety Committee of Seoul National University (SNUIBC-R190826-1).
Analysis of the Prevalence of Severe Fever with Thrombocytopenia Syndrome Virus Using RT-PCR and Enzyme Linked Immunosorbent Assay of Cat Sera From the Republic of Korea, 2017
ELISA, enzyme linked immunosorbent assay.
Reverse transcriptase-nested polymerase chain reaction
One-step reverse transcriptase (RT)-nested PCR was conducted to amplify the S segment of the SFTSV RNA gene using SFTSV genome-specific primer sets and a one-step RT-PCR premix kit (SolGent, Daejeon, ROK), as previously described (Hwang et al. 2017). The first set of PCR primers (expected amplicon size: 461 bp) were NP-2F (5′-CATCATTGTCTTTGCCCTGA-3′) and NP-2R (5′-AGAAGACAGAGTTCACAGCA-3′), and the nested PCR primers (expected amplicon size: 346 bp) were N2F (5′-AAYAAGATCGTCAAGGCATCA-3′) and N2R (5′-TAGTCTTGGTGAAGGCATCTT-3′). An SFTSV human isolate, strain KH1, was used as a positive control (Lee et al. 2019). RT reactions were performed at 50°C for 30 min followed by 94°C for 5 min. The PCR was performed according to the following conditions: 20 s at 94°C, 40 s at 52°C, and 30 s at 72°C for 40 cycles (first PCR) and 25 cycles (nested PCR), followed by a final extension step at 72°C for 5 min. The amplified PCR products were purified using QIAquick Gel Extraction Kits (Qiagen, Hilden, Germany).
Nucleotide sequencing and phylogenetic analysis
All amplicons were directly sequenced with an automatic sequencer (3730xl capillary DNA Analyzer; Applied Biosystems, Foster City, CA). The obtained sequences were analyzed using Chromas software (Ver 2.66). Sequences were aligned using Clustal X (Ver 2.1) and analyzed with MEGA 7 to compare sequences from this study with those previously deposited in the GenBank database. Phylogenetic trees were constructed using a maximum likelihood method based on the Kimura 2-parameter model, and the data set was resampled 1000 times to generate bootstrap values.
Indirect enzyme linked immunosorbent assay
Nucleocapsid protein (NP) recombinant protein was purified using HisTrap HP histidine-tagged protein columns (GE Healthcare, Chicago, IL) according to the manufacturer's instruction and as described previously (Kang et al. 2020). To evaluate the antibody titers of specific NP in cat sera, 96-well immunoassay plates (Nalge Nunc, Rochester, NY) were coated with 100 ng/well of purified NP and incubated at 4°C overnight. After antigen coating, the plates were incubated for 2 h at room temperature in a blocking solution (PBS containing 0.05% Tween 20 and 5% skim milk). Each serum sample (1 μL) was incubated for 90 min at room temperature and detected using horseradish peroxidase (HRP)-conjugated goat anticat IgG antibodies (Invitrogen, Frederick, MD). TMB (3,3′,5,5′-tetramethylbenzidine) substrate solution (KPL, Gaithersburg, MD) was then added to develop color for 7 min, and the reaction was stopped by the addition of 1 M H3PO4 solution. Optical density was measured at 450 nm using a microplate reader (Tecan, Mannedorf, Switzerland).
Results
Eight out of 201 (4.0%) cat sera were positive for SFTSV as assessed by RT-PCR. Of the 8 positive samples, 6 out of 101 (5.9%) feral and 2 out of 100 (2.0%) house cats were positive for the SFTSV gene. This indicated a slightly higher prevalence of SFTSV in feral than in house cats (p = 0.2791). The infection rate of SFTSV was higher in females than in males, with 3 out of 112 (2.7%) males and 5 out of 89 females (5.6%) testing positive for the SFTSV gene (p = 0.4702). The SFTSV S segment sequences (346 bp) obtained from cat sera showed 97.1–100.0% similarity and were deposited in GenBank under the accession numbers MN580502-KP580509. The phylogenetic analysis revealed that isolates from this study were clustered in the B-2 genotype (Fig. 1) (Yun et al. 2020). Moreover, the H89 sequence (MN580505) was identical to the dog-derived Dog22 strain from the ROK, and the H51 sequence (MN580502) was very similar to the cat-derived Cat/Miyazaki/2018 (99.4%) strain from Japan (Fig. 1).
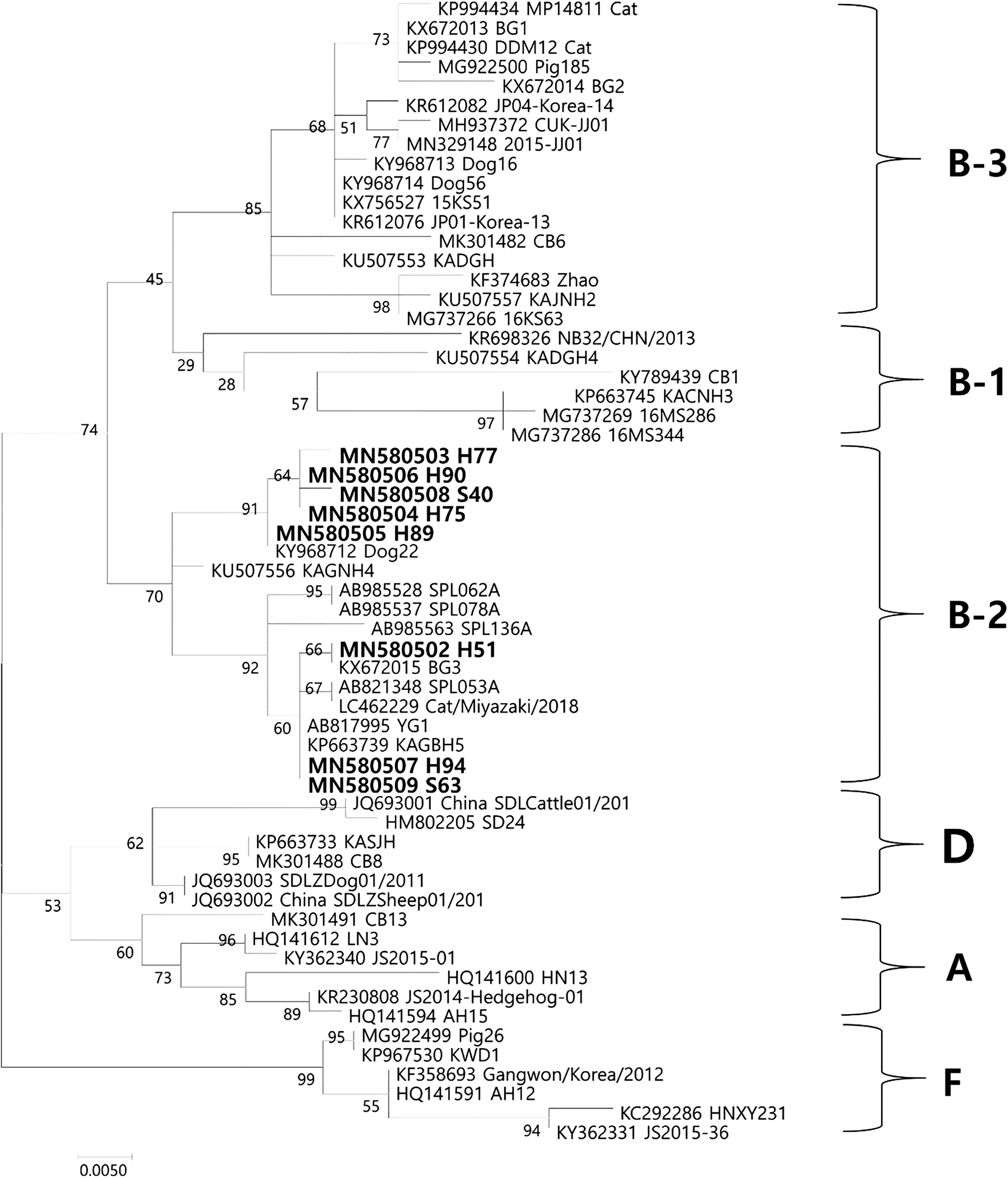
Phylogenetic analysis of SFTSV based on the partial S segment (346 bp). The partial S segment was amplified through RT-nested PCR, and the resulting products were sequenced and compared with previously deposited sequences. Phylogenetic analysis was performed, and the evolutionary history was inferred by using the maximum likelihood method based on the Kimura 2-parameter model (1000 bootstrap replicates). The percentage of trees in which associated taxa clustered together is shown next to the branches. The sequences identified in this study are indicated in bold letters. The scale bar indicates the number of nucleotide substitutions per position. SFTSV, severe fever with thrombocytopenia syndrome virus.
Nine out of 201 sera (4.5%) were positive for anti-SFTSV NP antibodies as assessed by enzyme linked immunosorbent assay (ELISA) (Table 1). The prevalence of anti-SFTSV antibodies was higher in feral cats, with 7 out of 101 (6.9%) feral and 2 out of 100 (2.0%) house cats testing positive. Unlike the levels of SFTSV genes, the levels of antibodies against SFTSV were slightly higher in males than in females, with 6 out of 112 (5.4%) males and 3 out of 89 females (3.4%) testing positive for anti-SFTSV antibodies. However, differences in the levels of anti-SFTSV antibodies were not significant between feral and house (p = 0.1005) or male and female (p = 0.4978) cats. No cats were positive for both the SFTSV gene and anti-SFTSV antibodies.
Discussion
The epidemiology of SFTSV has been surveyed in domesticated, wild, and companion animals, where different prevalence rates were reported depending on the animal species (Niu et al. 2013, Ni et al. 2015, Hwang et al. 2017, Lee et al. 2017, Matsuno et al. 2018, Kang et al. 2019, Matsuu et al. 2019). In these surveys, the overall levels of anti-SFTSV antibodies were higher than those of SFTSV genes. This is similar to our findings, where a slightly higher number of cats were positive for anti-SFTSV antibodies than the SFTSV gene. Furthermore, previous studies reported a high prevalence of anti-SFTSV antibodies in cattle (100%), goats (82%), and sheep (70%) (Chen et al. 2019, Huang et al. 2019). However, our study detected anti-SFTSV antibodies in only 4.5% of cat sera, suggesting species- and/or geographic-specific influences on prevalence. Previous SFTSV studies in the ROK have investigated several animals, including shelter dogs and cats, feral cats, goats, domesticated pigs, and wild animals (Oh et al. 2016, Hwang et al. 2017, Lee et al. 2017, Kang et al. 2018a, 2018b). In contrast to reports from China, the prevalence of the SFTSV gene or antibody in the ROK was <20%. These discrepancies could be explained by differences in sampling sites, antibody and gene detection protocols, and animal raising methods.
Two previous studies revealed a significant difference in the prevalence of SFTSV between feral (17.46%) and shelter cats (0.47%) (Hwang et al. 2017, Lee et al. 2017). In comparison, we found lower SFTSV infection rates of feral cats (5.9%), but a higher rate in house cats (2.0%) than reported for shelter cats. In general, it is unlikely that cats in the ROK will be exposed to the outside environment, as house cats cannot go outdoors in this country. These differences may also be associated with variations in sample sizes, collection sites, and sampling periods.
Apart from a case report, no serological survey of cats has been reported to date. However, this study shows for the first time a larger scale serological survey of SFTSV in cats. When compared with other animal species, the antibody-positive rate in cats (4.5%) is very low, further suggesting that cats in a metropolitan city are less likely to be exposed to SFTSV. However, due to differences in methodologies, it is difficult to confidently compare our antibody-positive rate to other studies. Further surveys with higher sample sizes, a range of species, and consistent methodologies are needed to fully understand the prevalence of SFTSV in cats.
Recent studies have demonstrated that SFTSVs could be classified into several clades by phylogenetic analysis (Yoshikawa et al. 2015, Yun et al. 2020). SFTSVs could be divided into two major clades at an early evolutionary point. One clade consists mainly in Chinese SFTSV strains and the other clade is composed of Korean and Japanese SFTSV strains (Fig. 1). Although the partial sequences of the S segment were used in this study, the results of the phylogenetic analysis were very similar to full sequences of the S segment. All sequences obtained belonged to the B-2 genotype (Fig. 1; Yun et al. 2020) and the J1 clade (data not shown, Yoshikawa et al. 2015), which is mainly composed of Korean and Japanese strains. The results of this study were consistent with previous reports, suggesting that Korean SFTSVs are more closely related to Japanese SFTSVs than to Chinese groups and that various genotypes of SFTSV exists in the ROK.
Although SFTSV transmission without tick biting has rarely been reported (Jung et al. 2019, Kida et al. 2019), cats could be a source of direct transmission to humans or other animals. Cats are highly susceptible to SFTSV, and high levels of SFTSV RNA have been detected in the serum, eyes, and saliva in an experimental environment (Park et al. 2019). Although direct SFTSV transmission has not been confirmed, a clinical case of cat-to-human transmission of SFTSV was reported in Japan (Kida et al. 2019). Cats usually lick themselves for self-grooming, thereby exposing their hair or skin to potentially infected saliva. Furthermore, cats sometimes scratch other animals or humans, providing an opportunity for SFTSV transmission if high viral loads are present. In contrast, it is very difficult to explain about how house cats were exposed to the virus. So, further studies are required to elucidate the potential transmission cycle of SFTSV and the first infection route of SFTSV in companion animals.
Conclusions
SFTS is an emerging zoonotic disease that is an important public health consideration, with human infections becoming endemic in Asia (Yu et al. 2011, Kim et al. 2013, Takahashi et al. 2014, ICTV 2018). Although the animal host of this vector-borne disease is currently unconfirmed, this study detected viral genes and antibodies in house and feral cats. Humans live in close contact with domesticated animals, and SFTSV-infected felines or humans could transmit SFTSV. This is the first study to demonstrate the presence of anti-SFTSV antibodies in the sera of house and feral cats in the ROK. Although direct transmission has not been reported, results from this study suggest that veterinarians and companion animal guardians should be cautious when handling an animal suspected of being infected. Because the ROK is an endemic area for SFTSV, precautions and continuing education are needed for guardians and veterinarians.
Footnotes
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This research received funding from the Basic Science Research Program through the National Research Foundation of Korea funded by the Ministry of Education (NRF-2018R1D1A1B07049140) and the Government-wide R&D Fund for Infectious Diseases Research (HG18C0021).