
Abstract
Background:
Wuxiang virus was isolated from sandfly specimens collected in Wuxiang County, Shanxi Province, China in 2018, representing the first reported isolation of sandfly-borne virus from sandflies collected in a natural environment in China. The local sandfly density, seasonal fluctuations, and temporal and spatial distributions of the virus in Wuxiang County remain unclear.
Materials and Methods:
Four fixed sandfly collection sites were set up in Wuxiang County and sandfly specimens were collected continuously from June to August 2019. All sandfly specimens were subjected to viral isolation and molecular biological analysis.
Results:
The data on sandfly specimens collected in Wuxiang County from June to August 2019 showed a significant difference in the density of sandflies between June 26 and August 16 (p < 0.05). No statistical difference was found in sandfly density among collection sites (p > 0.05). A total of 33 virus isolates causing cytopathic effects in mammalian (BHK-21) cells were obtained from 7466 sandflies (91 pools) collected from June to August 2019. The results of molecular genetic evolution analysis of the nucleotide sequence of these isolates showed that the L and S genes (encoding NS and N proteins) of the 33 viruses isolated in 2019 are in the same evolutionary branch as the previously isolated Wuxiang virus. No significant difference was found in the virus isolation rate (the pool isolation rate of virus) among sandflies collected at different times from June to August (p > 0.05). The virus isolation rate of sandflies collected at different collection sites showed a statistically significant difference (p < 0.05).
Conclusions:
The results of this study suggest that the Wuxiang virus is a stable viral population in local sandflies. Strengthened research into Wuxiang virus infection of humans and animals and clarification of the public health hazards posed by Wuxiang virus to both humans and animals are urgently needed.
Introduction
The genus Phlebovirus belongs to order Bunyavirales and family Phenuiviridae. The International Committee for the Taxonomy of Viruses (ICTV) currently recognizes 10 viral species within the genus Phlebovirus, including Rift Valley fever phlebovirus, sandfly fever Naples virus (SFNV), and Punta Toro phlebovirus (Abudurexiti et al. 2019).
The transmission vectors of Phlebovirus include sandflies, mosquitoes, and ticks (Alkan et al. 2013). Among them, sandfly-borne phleboviruses, such as sandfly fever Sicilian virus (SFSV) and SFNV, can cause febrile diseases after infection, referred to as “three-day fever” (Ayhan et al. 2017). The Toscana virus transmitted by sandflies can cause acute meningitis (Depaquit et al. 2010). Thus, sandfly-borne viruses are closely associated with human public health risks. Sandfly-borne viruses and related diseases occur mainly in Mediterranean countries, such as Italy (Vocale et al. 2012), Turkey (Ergunay et al. 2011), and Cyprus (Papa et al. 2006), but have not been reported in Mainland China or other parts of eastern Asia (Depaquit et al. 2010, Liang et al. 2018).
We reported the isolation of a new type of Phlebovirus, Wuxiang virus, from sandfly specimens collected in Wuxiang County, Shanxi Province, Central China. This is the first report of a Phlebovirus that was isolated from sandfly specimens collected from the natural environment in China (Wang et al. 2020), suggesting that viruses transmitted by sandflies occur not only in the Mediterranean Basin, but also in Mainland China. Although phleboviruses have been isolated from sandfly specimens collected in China, the seasonal fluctuations of local sandfly populations, the density of sandflies in various habitats, and the temporal and spatial distributions of viruses carried by local sandflies remain unclear.
In this study, we collected sandfly specimens and conducted virus isolation and identification in Wuxiang County between June and August 2019 to clarify the temporal and spatial distribution characteristics of local sandflies and viruses and to provide basic data for control of the public health hazards associated with sandflies and the viruses they carry. The results are reported below.
Materials and Methods
Specimen collection
Four fixed sandfly specimen collection sites were set up in two villages in Wuxiang County, Shanxi Province, from June to August 2019. Two collection sites (collection site 1: 1 mule, collection site 2: 1 mule, and 10 chickens) were located in Moyu Village; and 2 collection sites (collection site 3: 1 mule and 1 dog, collection site 4: 10 chickens) were located in Qianshenlianggou Village (see Fig. 1 for sandfly collection sites). At each collection site, four sandfly collection campaigns were conducted between June and August. Each collection site was equipped with a sandfly collection tool (Wen Taitai MM200; manufacturer: Guangzhou Changsheng Chemical Technology Service Co., Ltd.). The specimen collection time was 18:00 PM to 6:00 AM on the following day. All collected sandfly specimens were placed in a low-temperature refrigerator for 30 min and then removed for processing under ice bath conditions. All specimens were classified according to morphology, collection time, and collection site. Specimens were stored in liquid nitrogen until laboratory testing (Huemer et al. 2017, Song et al. 2017, Wang et al. 2020).

Collection sites of sandfly specimens in Wuxiang County, Shanxi Province in 2019. Note: MY and QSLG are located ∼5 km apart, and four collection sites were set up in the two villages (1, 2, 3, and 4 in red in the figure). The two collection sites in Moyu Village are referred to as 1 and 2, and are marked in red in the figure. The two collection sites in Qianshenlianggou Village are referred to as 3 and 4, as shown in red in the figure. Collection site 1 is located at the edge of Moyu Village, near an area of natural environment, whereas collection sites 2, 3, and 4 are all located in internal areas of the villages. The distances from 1 to 2, 3, and 4 are 0.3 km, 4.5 km, and 4.8 km, respectively. MY, Moyu Village; QSLG, Qianshenlianggou Village. Color images are available online.
The density of sandflies was calculated according to the following formula: sandfly density = number of sandflies collected/(number of lights × number of nights of sampling) (Banjara et al. 2019).
Cells
BHK-21 cells (golden hamster kidney cells) and C6/36 cells (Aedes albopictus egg cells) were maintained in the laboratory. The BHK-21 cell culture medium included 90% Eagle's (laboratory preparation), 7% fetal bovine serum (FBS; Invitrogen), 1% penicillin and streptomycin (100 U/mL), 1% glutamine (30 g/L), and 1% NaHCO3; C6/36 cell culture medium contained 89% RPMI 1640 (Invitrogen), 10% FBS (Invitrogen), and 1% penicillin and streptomycin (100 U/mL). BHK-21 and C6/36 cells were cultured in incubators at 37°C with 5% CO2 and 28°C, respectively (Fu et al. 2017, Song et al. 2017).
Virus isolation
Pools of 50–100 sandflies or 50 mosquitoes were ground using a glass grinder, added to grinding fluid (93% Eagle's, 5% penicillin and streptomycin [100 U/mL], 1% glutamine [30 g/L], and 1% NaHCO3), washed twice, and then added to 1.5 mL fresh grinding fluid and ground under ice bath conditions until the specimen was completely homogenized. The samples were centrifuged (4°C, 12,000 rpm, 30 min) after grinding, and 100 μL of the ground supernatant was inoculated on an 80% confluent monolayer of BHK-21 or C6/36 cells on culture plates (24-well plate; Corning Incorporated). BHK-21 and C6/36 cells were continuously cultured in an incubator at 37°C with 5% CO2, and at 28°C, respectively. Cells were observed for signs of cytopathic effects (CPEs) under a microscope every 12 h. When the cells showed CPEs, the virus solution was collected and stored in a low temperature refrigerator at −80°C for further identification. The first-generation cell culture supernatants without CPEs were blindly passed through the above two cell types for three generations, and those without CPEs were discarded (Fu et al. 2017, Song et al. 2017, Wang et al. 2020).
Species identification of sandfly specimens
According to the instructions of the DNA Extraction Kit (QIAamp; Qiagen, Valencia, CA), DNA was extracted from sandfly specimens from which viruses were isolated. PCR was used to amplify the cytochrome c oxidase I (COI) gene using primer (LCO1490: GGTCAACAAATCATAAAGATATTGG and HCO2198: TAAACTTCAGGGTGACCAAAAAATCA) (Folmer et al. 1994). The positive PCR products were sequenced using four-color fluorescently labeled dideoxy termination method in Tianyi Huiyuan Biotech Co., Ltd. (Beijing, China). The sequences were Blast aligned in GenBank on the NCBI website (
Viral RNA extraction and cDNA library preparation
According to the instructions of the Viral RNA Mini Kit (QIAamp; Qiagen), total RNA was extracted from samples. The extracted total RNA was incubated in a 65°C water bath for 10 min, and in an ice bath for 2 min immediately, then 32 μL RNA was added to the first-strand reaction tube of the Ready-To-Go Kit (GE Healthcare, Little Chalfont, Buckinghamshire, United Kingdom), which was maintained at room temperature for 1 min before the addition of 1 μL of random primers (TaKaRa, Shiga, Japan). This mixture was centrifuged immediately and incubated in a water bath at 37°C for 1 h. The total volume of the cDNA library was 33 μL, and cDNA was used immediately or stored at −40°C for future use (Feng et al. 2017, Ren et al. 2017).
Viral gene amplification and nucleotide sequencing
The viral gene amplification (PCR) system was 25 μL and contained cDNA template, GoTaq® Green Master Mix 2 × (Promega, Madison, WI), and 10 μmol/L upstream and downstream primers. Wuxiang virus gene-specific primers were used for PCR amplification of samples (Wang et al. 2020). After the PCR reaction, 5 μL of the gene amplification product was subjected to 1% agarose gel electrophoresis. Products showing successful gene amplification were subjected to nucleotide sequencing (Feng et al. 2017, Ren et al. 2017).
Nucleotide sequence analysis
The nucleotide sequences were aligned using BLAST (NCBI). Seqman software (DNAStar, Madison, WI) was used for nucleotide sequence splicing and quality analysis; BioEdit (Version 7.0; Thomas) software was used for nucleotide multiple sequence alignment; and MEGA6.0 software was used to conduct phylogenetic analysis based on the neighbor-joining method, with a bootstrap value of 1000. Information about the strains used in viral molecular genetic evolution analysis is provided in Supplementary Table S1 (Feng et al. 2017, 2019, Song et al. 2017).
Statistical methods
The data were statistically processed using SPSS version 25 software, and the density of sandflies at different times and different collection sites were compared using the nonparametric Kruskal–Wallis test. The temporal and spatial distributions of sandflies carrying viruses were tested using the χ2 test and Fisher's exact test, with p < 0.05 considered statistically significant.
Results
Collection of sandfly specimens
This study was conducted in Moyu Village (113°5′ east longitude, 36°47′ north latitude, elevation 966 meters above sea level) and Qianshenlianggou Village (113°2′ east longitude, 36°46′ north latitude, elevation 910 meters) in Wuxiang County, Shanxi Province, on June 14, June 26, July 19, and August 16, 2019. A total of four fixed collection sites were placed at livestock houses and sampled four times for sandfly specimen collection (collection sites are shown in Fig. 1). A total of 7466 sandflies and 749 mosquitoes were collected in the 2 villages, including 340 Anopheles sinensis, 327 Armigeres subalbatus, 37 A. albopictus, and 45 Culex pipiens pallens.
Spatial and temporal distributions of local sandflies
Seasonal changes in sandfly abundance
The results in Table 1 show that the density of sandflies collected in this study was 466.63 per light per night, but the densities of sandflies collected at the four time points from June to August differed. The density of sandflies was 458.75 per light per night on June 14, 2019, 1116.25 per light per night on June 26, 284 per light per night on July 19, and 7.5 per light per night on August 16. Thus, the density of sandflies was highest on June 26, and very few sandflies were collected on August 16. According to statistical analysis, the density of sandflies differed among the four collection times (χ2 = 10.339, p < 0.05). Based on pairwise comparison, the densities of sandflies on June 26 and August 16 were significantly different (p < 0.05).
Sandfly Collection in Wuxiang County, Shanxi Province, in 2019
Sandfly density is the number of sandflies captured per sandfly collection tool per night.
Distribution of sandflies in various habitats
In this study, four livestock houses were selected as sandfly collection sites. Table 1 shows that sandflies were captured in four different livestock houses at densities of 847.75, 411.5, 525.75, and 81.5 per light per night, respectively. Among collection sites, collection site 1 (1 mule) had the highest sandfly density, while collection site 4 (10 chickens) had the lowest density. However, no statistically significant difference was found in sandfly density among the four different livestock houses (χ2 = 3.54, p > 0.05). In this study, collection sites 1 and 2 were located in Moyu Village, and 3 and 4 were located in Qianshenlianggou Village. A total of 5037 sandflies were collected in Moyu Village and 2429 sandflies were collected in Qianshenlianggou Village. The density of sandflies collected in the two villages was 629.63 per light per night and 303.63 per light per night, respectively. Statistical analysis showed no significant difference in the density of sandflies between the two villages (p > 0.05).
Virus isolation and identification
The blood-sucking insect samples were pooled, and the supernatants from 106 ground sample pools (91 sandfly pools and 15 mosquito pools) were used to inoculate BHK-21 and C6/36 cells, respectively. In total, 33 strains of virus isolates capable of infecting BHK-21 cells were obtained by culturing BHK-21 cells with the ground supernatants of sandfly samples (Table 2). Figure 2 shows CPEs on the third day after inoculation of BHK-21 cells with virus isolate SXWX1924-1, exhibiting cell shrinkage and shedding. Gene amplification and nucleotide sequencing were performed on the 33 pools of cytopathic BHK-21 cell culture supernatants using primers specific to the L gene and S gene (NS and N proteins) of the Wuxiang virus, and all 33 virus isolates showed positive amplification of the L and S genes of Wuxiang virus.
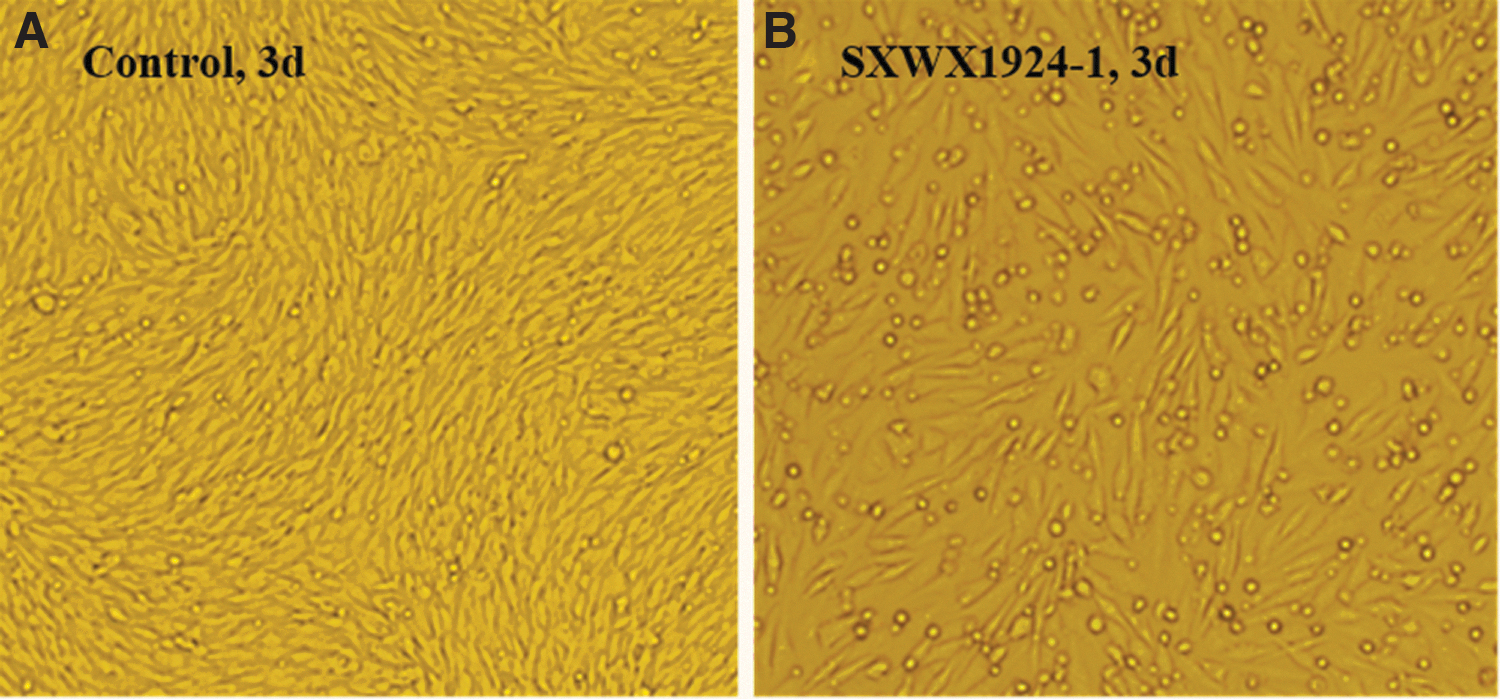
CPEs of virus isolate SXWX1924-1 in BHK-21 cells.
Wuxiang Virus Isolates in 2019
+ Indicates CPEs; — indicates no CPEs.
CPE, cytopathic effect.
No CPEs were observed in C6/36 cells inoculated with 91 pools ground supernatants of sandfly samples for 3 consecutive generations, and no positive gene amplification was detected in the C6/36 cell supernatant using primers specific to the L and S genes of Wuxiang virus. A total of 15 pools of mosquito specimens were cultured continuously in BHK-21 and C6/36 cells, and no CPEs or positive gene amplification were observed. These results indicate that Wuxiang virus does not cause CPEs or gene amplification in C6/36 cells and that Wuxiang virus was not present in local mosquito specimens.
Species identification of sandfly specimens
To identify the sandfly species, PCR was used to amplify the COI gene, and the results showed that 33 pools of sandfly samples, from which viruses were isolated, are identified as Phlebotomus chinensis. The nucleotide sequence of sandfly COI gene was BLAST aligned on the NCBI website, and the results showed that the COI gene sequence of 33 Pools of sandfly samples had more than 97% homology with GenBank no. MF966721 (P. chinensis). Species identification sequences of 33 pools of P. chinensis were added into Supplementary Sequence File.
Molecular genetic evolution of virus isolates collected in Wuxiang in 2019
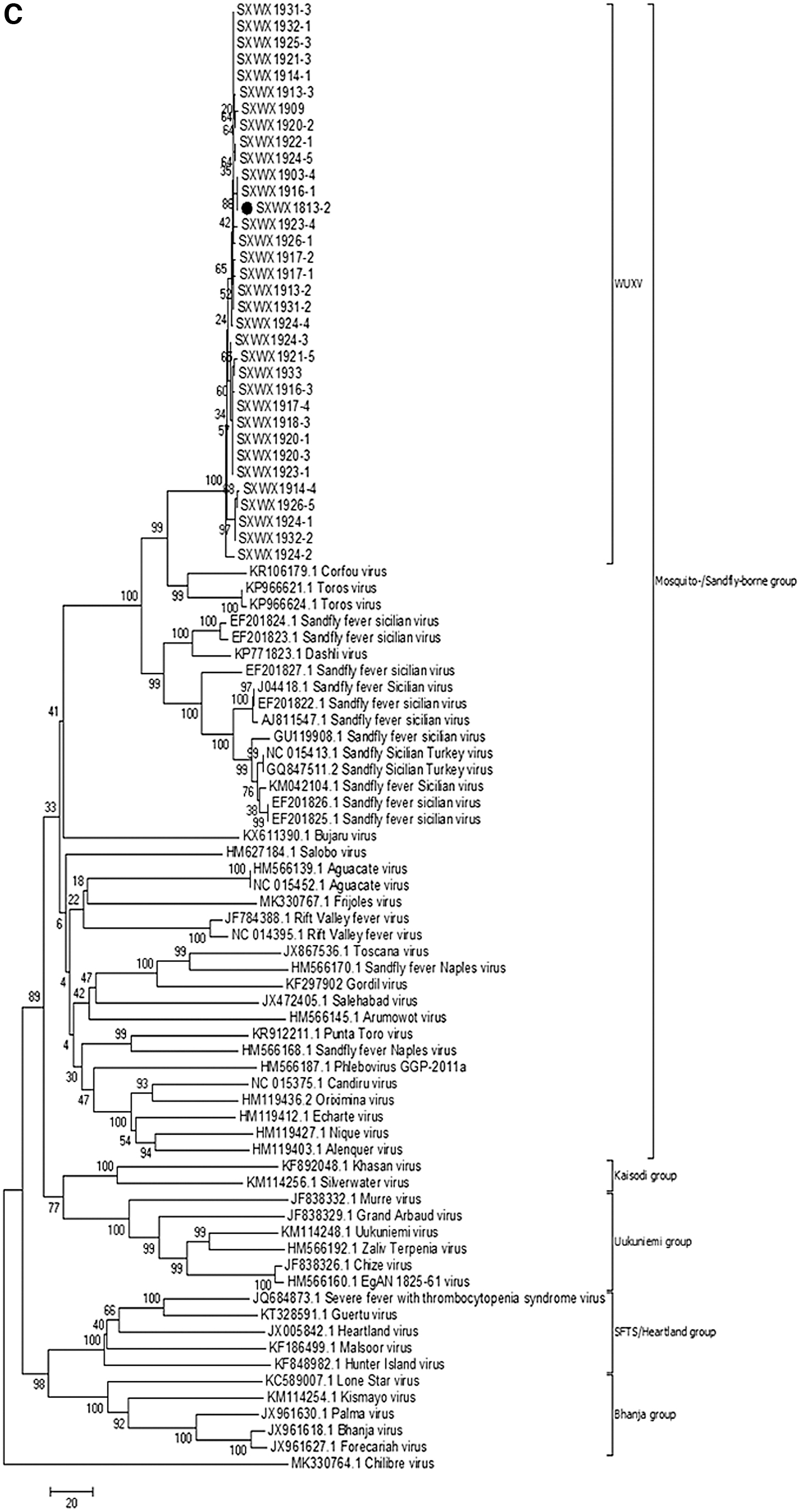
The S gene sequence (NS and N proteins) and L gene sequence of 33 virus isolates and the corresponding nucleotide sequences of Phlebovirus were analyzed to explore viral molecular genetic evolution. The results showed that all 33 virus isolates, based on both the L and S genes (NS and N proteins) of the virus, are members of the mosquito-borne or sandfly-borne virus group within the genus Phlebovirus. Further analysis showed that the 33 virus isolates isolated from sandfly specimens in Wuxiang County in 2019 are on the same evolutionary branch as the representative strain of Wuxiang virus, SXWX1813-2 (Fig. 3).

Molecular genetic evolution of virus isolates collected in Wuxiang in 2019.

Spatial and temporal distributions of Wuxiang virus carried by sandflies
Seasonal changes in the isolation rate of Wuxiang virus
A total of 7466 sandflies were collected in Wuxiang County, Shanxi Province, from June to August 2019 (Table 1). A total of 33 Wuxiang virus isolate strains were obtained from inoculating cells with 91 pools of ground sandfly specimens. Therefore, the virus isolation rate of Wuxiang virus in the local sandfly population is 36.26% (33/91). The virus isolation rates of sandflies collected at the four time points from June 14 to August 16 were 24% (6/25), 39.22% (20/51), 50% (7/14), and 0% (0/1), as shown in Table 3. Although the isolation rates of viruses collected at the four time points differed markedly, only the isolation rate of sandflies collected on July 19 was found to be significantly higher than those of other times. Statistical analysis showed no significant difference in the virus isolation rate among the four collection times (p > 0.05).
Isolation Rates of Sandflies Carrying Viruses from June to August 2019
Isolation rates of Wuxiang virus in various habitats
In this study, the virus isolation rates at the four sandfly collection sites were as follows: 22.50% (9/40) at collection site 1, 40% (8/20) at collection site 2, 57.69% (15/26) at collection site 3, and 20% (1/5) at collection site 4. Collection site 3 (1 mule and 1 dog) had the highest virus isolation rate, whereas collection site 4 (10 chickens) had the lowest virus isolation rate. Differences were observed in the isolation rates of Wuxiang virus from sandfly specimens collected in four different livestock houses (χ2 = 8.865, p < 0.05). Through pairwise comparison, a statistically significant difference was found in the isolation rate of the virus (p < 0.01) between collection site 1 (one mule) and collection site 3 (one mule and one dog). Comparing the two villages in which the four collection sites were located, the virus isolation rate in Moyu Village was 28.33% (17/60), whereas that in Qianshenlianggou Village was 51.61% (16/31). Analysis showed that the virus isolation rates in the two villages differed significantly (χ2 = 4.793, p < 0.05), as shown in Table 3.
Discussion
In previous arbovirus surveys, sandfly-borne viruses in China have been investigated. However, for various reasons (mainly the limited number of sandflies collected in each survey), no viruses had yet been isolated from sandfly specimens collected in natural environments in China (Liang et al. 2018). Our team collected sandfly specimens in Wuxiang County, Shanxi Province, Central China, in 2018 and isolated the Wuxiang virus from those specimens. This report describes the first isolation of a virus from sandfly specimens collected from the environment in Mainland China (Wang et al. 2020). The research team conducted several surveys in Wuxiang County in 2019, with the goal of analyzing the ecological habits of local sandflies and their status as carriers of Wuxiang virus.
Ecological habits of sandflies in Wuxiang County
In this study, we conducted sandfly density surveys in Wuxiang County from June to August 2019. Among sampling times, the density of sandflies collected was greatest on June 26 at 1116.25 per light per night, whereas that in August was 7.5 per light per night (Table 1). Wuxiang County is located in the Taihang Mountains in central China. The nighttime temperature in April and October is ∼0°C, which is not conducive to the growth of sandflies. In this study, nine and three sandflies were collected in Wuxiang County in May and September, respectively, but these specimens are not included in Table 1 due to their small numbers. Therefore, the annual emergence of sandflies may occur in May in Wuxiang County. The number of sandflies is greatest from June to July, and they disappear in September. These results suggest that the sandflies in Wuxiang County have a single-generation growth characteristic. Studies have shown that P. chinensis is the dominant species of sandfly in northern China (Guan et al. 2016, Chen et al. 2020). In the latest study (Chen et al. 2020), sandflies in Wuxiang County were identified as P. chinensis, and our research of sandflies in Wuxiang County was also identified as P. chinensis, the two results were consistent and mutually supportive. Guan et al. (1980) found that P. chinensis appears in May every year, with a peak period from June to July, and disappears in mid-to-late August, with only one generation reproducing per year in northern China, it is consistent with the results obtained in our study. Some studies have shown that certain climatic factors, such as temperature and relative humidity, are related to sandfly density (Pareyn et al. 2019). A temperature rise to 16–33.3°C coupled with a relative humidity of 60% may increase the density of P. chinensis in northwestern China (Guan et al. 1980, Xiong et al. 1984). This finding indicates that the Wuxiang area provides optimal temperature and relative humidity conditions for the survival of sandflies from June to July. Previous investigations on the ecological habits of P. chinensis in the natural environment of the Loess Plateau in China showed that P. chinensis breeds in the wild, and lives in nearby villages. This species sucks blood from cows, donkeys, mules, chickens, and domestic dogs (Xiong 1992). In this study, the results from four collection campaigns conducted in four livestock houses in two villages in Wuxiang County showed that no significant difference in the number of sandflies among the four livestock houses was used as collection sites (χ2 = 3.54, p > 0.05).
Wuxiang virus is a stable viral population in local sandflies
The results of molecular genetic evolution analysis show that the virus isolated in this study, regardless of whether the L gene or S gene (NS or N) was analyzed, falls on the same evolutionary branch as the Wuxiang virus isolated from sandfly specimens in Wuxiang County in 2018 (Fig. 3). Although the virus isolation rate from sandfly specimens in this study differed from June to August 2019, for example, with rates of 24% on June 14, 39.22% on June 26, 50% on July 19, and 0% on August 16, no statistical difference was found among collection times (p > 0.05). From January 1969 to March 1971, a survey on viruses carried by sandflies was conducted in Panama. The results showed that the isolation rate of sandfly viruses (such as Chagres virus) was generally constant throughout the study period (Tesh et al. 1974). The results of the present study suggest that Wuxiang virus is not an occasional virus in sandflies in Wuxiang County, but rather that a stable viral population is present in sandflies in this area.
The sandflies transmit sandfly virus to vertebrates mainly by feeding their blood. After vertebrates are infected with the virus, viremia is produced and maintained for a certain period of time before the virus can be transmitted to arthropods, in which it reproduces. Vertebrates are an essential host for virus transmission (Karabatsos 1978, Hall et al. 2016). Studies have shown that SFSV shows a high positive rate of neutralizing antibodies in dogs. For example, the level of antibodies is 50.8% in dogs in Portugal (Alwassouf et al. 2016b), 38.1–59.2% in Tunisia (Sakhria et al. 2014), 71.9% in Greece, and 60.2% in Cyprus (Alwassouf et al. 2016a). Sandflies in different regions are known to favor different blood supply hosts. Studies conducted in Sichuan Province, China, showed that pigs are the preferred blood source for local P. chinensis (Chen et al. 2016), whereas Phlebotomus perniciosus in western Spain fed on a wide range of blood supply hosts, including pigs, humans, sheep, chickens, horses, and donkeys (Bravo-Barriga et al. 2016). An Ethiopian study in Northwest Africa showed that the most common blood source for local sandflies was donkeys (33.9%), followed by cattle (24.2%), humans (17.6%), dogs (11.8%), and goats or sheep (8.6%) (Yared et al. 2019). According to the latest study in 2017 (Chen et al. 2020), 64.8% (70/108) of 108 sandflies collected in Wuxiang County where the virus was isolated in this article, were detected as human blood, wereas the positive rates of dog blood, chicken blood, and goat blood were 0.9% (1/108), 21.3% (23/108), and 34.3% (37/108), respectively, suggesting that the bloodsucking rate of local sandflies to humans was higher compared with local animals such as dogs, chickens and goats. Wuxiang virus is a stable viral population that is present in local sandflies, and whether it can infect people and animals, we will next collect the sera of local people and animals to carry out the serological epidemiological study of sandfly virus infection.
Conclusions
In this study, we investigated the basic ecological habits of sandflies at virus isolation sites in Wuxiang County and their relationships with the virus. After several consecutive sandfly collection events and virus isolation from the collected specimens, we found that June to July is the peak period for sandflies, and Wuxiang virus is a stable viral population in local sandflies. These results provide important basic data for elucidating the relationship between sandflies and viruses in Wuxiang County.
However, some questions require further research, including the reproductive cycle of Wuxiang virus in local animals, and identification of its animal reservoir. In addition, sandfly-transmitted viruses are prevalent mainly in Mediterranean countries, such as Italy and Cyprus, but Wuxiang County is tens of thousands of kilometers from the Mediterranean. Thus, the relationship between Wuxiang virus and sandfly-borne viruses that occur in the Mediterranean region requires further investigation to explore possible similarities between the origins of the viruses in these two areas. In particular, we only collected sandfly specimens from livestock houses in Wuxiang County, but did not set up any collection sites around human habitations. Therefore, this study cannot provide information about the status of local sandflies in human residences in Wuxiang County. In addition, information on local human and animal infections with Wuxiang virus remains lacking. This shortcoming is the subject of upcoming work by our research group. This research is of great significance, as it clarifies the public health hazards faced by local humans and livestock due to sandfly virus infection.
Footnotes
Author Contributions
X.X. and G.L. were involved in drafting the article. X.X. and G.L. were involved in revising the article. X.X., J.C., S.F., Q.W., J.W., X.L., X.T., J.C., S.N., Y.H., F.L., and S.X. participated in collection of sandfly specimens and acquisition of data. X.X., J.C., S.F., and G.L. participated in the analysis and interpretation of data. J.C., S.F., H.W., B.W., and G.L. supported funding for the study. H.W., B.W., and G.L. contributed to conception and design of the article.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This work was supported by the National Science and Technology Major Project of China (2018ZX10711001, 2018ZX10734404-003, 2018ZX10102001-001); the National Key Research and Development Program (no. 2018YFA0900800); Key Research and Development (R&D) Projects of Shanxi Province, China (201803D31205); and the Development Grant of State Key Laboratory of Infectious Disease Prevention and Control (2015SKLID505, 2014SKLID103).
Supplementary Material
Supplementary Sequence File
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.