
Abstract
The immune response to Brucella abortus mainly depends on antigen-specific T cell activation, CD4+ and CD8+ T cells, and Brucella-specific humoral response. Protective immune response against Brucella infection has not been performed in the Sprague-Dawley (SD) rat model. We measured bacterial kinetics in addition to in vivo and in vitro interferon gamma (IFN-γ) and interleukin-10 (IL-10) production against crude Brucella protein in the SD rats at different days of postinfection with B. abortus biotype 1 by indirect enzyme-linked immunosorbent assay. Forty SD rats were inoculated intraperitoneally with 0.1 mL sterile injectable pyrogen-free solution containing 1 × 1010 colony-forming units/mL of B. abortus biotype 1 obtained from cattle in Korea. Four rats were used as uninfected control. Serum IFN-γ level at 3 and 7 days postinfection were significantly higher (p > 0.001) compared with the IL-10 level. On the contrary, serum IL-10 levels were observed significantly higher at 21 and 28 days postinfection compared with the serum IFN-γ levels (p < 0.001). The production of IFN-γ by spleen cells was significantly higher at 7 and 14 days postinfection compared with IL-10 (p < 0.001). On the contrary, IL-10 productions were found to be significantly higher at 21, 28, 35, and 42 days postinfection compared with IFN-γ (p < 0.001). The presence of B. abortus in blood was marked till 5 weeks of infection, throughout the experiment in case of spleen, and no bacteria were isolated from the kidney and liver at 6 weeks postinfection. The in vivo and in vitro IFN-γ and IL-10 measurement in our study reported that B. abortus infection in rats primarily educe T helper (Th)1-dominant immune response in acute infection accompanied by Th2-dominant immune response in chronic infection.
Introduction
Brucellosis is a highly contagious bacterial disease caused by the ingestion of unpasteurized milk, dairy products, and undercooked meat from animals infected with Brucella spp. (Lai et al. 2017, Dadar et al. 2019). Humans and animals also infected through aerosol exposure to pathogens (Boschiroli et al. 2001, Pappas et al. 2005). Brucellosis is a widespread zoonosis caused by Brucella species (Grilló et al. 2012). In cattle, brucellosis is mainly caused by Brucella abortus infection (Poester et al. 2013). It has important public health significance because of its zoonotic characteristics (Olsen and Bellaire 2013).
In animals, it causes abortion or low fertility (Carvalho Neta et al. 2010). Brucellosis in humans may include fever, joint pain, fatigue, and headache. Despite that, in severe cases, it causes arthritis, epididymo-orchitis, hepatitis, and endocarditis (Dean et al. 2012, Martirosyan and Gorvel 2013, Young et al. 2014). Based on history, symptoms, and clinical presentation, brucellosis patient can be divided into three groups such as acute (0–2 months), subacute (3–12 months), and chronic (>1 year) (Jimenez de Bagues et al. 2005, Galinska and Zagorski 2013). Despite quick diagnosis and successful treatment, ∼10% of patients progress to chronic disease such as atypical symptoms of brucellosis and relapses (Berbari and Wilson 2001, Irmak et al. 2003).
The immune response to B. abortus mainly depends on antigen-specific T cell activation, CD4+ and CD8+ T cells, and Brucella-specific humoral response. After the invasion of the bacteria through mucosal surface, they use various tricks to circulate the host immune system and insist to survive the host to develop chronic infection (Köhler et al. 2003). Therefore, brucellosis is a serious economic and health problem disease because it is very difficult to diagnose early and prevent (Golding et al. 2001). T helper (Th) lymphocytes are subdivided into mainly Th1 and Th2 subgroups in accordance with the cytokines they secrete (Mosmann and Sad 1996). Th1 responses are mainly responsible for eliminating the Brucella infection (Ko et al. 2002). Interferon gamma (IFN-γ) is one of the main cytokine of Th1 responses that is mostly involved in protection against Brucella spp. (Baldwin and Parent 2002). Interleukin-10 (IL-10), which is secreted by Th2 cells, is mainly involved in the protection of extracellular pathogens through antibody production (Romagnani 1997).
Cytokines have a vital role in the pathogenesis of Brucella and they likely modulate host's sensitivity or resistance to Brucella (Galanakis et al. 2002, Demirdag et al. 2003). The cellular immune response is a critical part of the host defense against Brucella infections (Romagnani 2000). The immune response of Brucella spp. involves the full spectrum of the immune system, from innate to adaptive immunity (Golding et al. 2001). Th1 cells are intermediary of the effector mechanisms necessary for resistance to intracellular pathogens, whereas a Th2 cell response is harmful in contesting this type of infection (Yingst and Hoover 2003).
Sensitivity or resistance to various infectious diseases is connected to the manifestation of particular cytokine profiles. Investigation of cytokine profiles aid to refine functional properties of immune cells, both for research and for clinical diagnosis (Wood and Seow 1996). To know the response of immune system against infectious agent, evaluation of Th1 and Th2 cytokine response is necessary. The fate of a disease depends on the equality of Th1 and Th2 cytokines (Romagnani 1997).
Rats can be infected with Brucella (Islam et al. 2012). Rats harbor places are commonly in houses and cattle farms. Infected rats could excrete Brucella organisms through feces (Corbel 1997) and transmit brucellosis to humans and animals through direct contact. The mouse model is used to study chronic Brucella infection (Silva et al. 2011). Our previous study measured humoral immune responses and distribution of B. abortus Biotype 1 infection in Sprague-Dawley (SD) rats (Khatun et al. 2015).We also measured profiles of IgG and its subclasses (IgG1 and IgG2a) against lipopolysaccharide, whole cell antigen, outer membrane protein, periplasmic protein, cytoplasmic protein, and crude Brucella protein (CBP) of B. abortus biotype 1 by enzyme-linked immunosorbent assay (ELISA) (Khatun et al. 2009, 2020). Most of the experimental Brucella infection to see the immune response is studied in mouse models (Baldwin and Parent 2002). No information is available about IFN-γ and IL-10 measurement related to B. abortus infection in rats. The in vitro and in vivo IFN-γ and IL-10 measurement using CBP during acute and subacute stages of infection with B. abortus biotype 1 were investigated in SD rats in this study.
Materials and Methods
Ethical approval
The protocol of the experiment was accepted by the animal ethical committee of the Chonbuk National University, Jeonju, South Korea.
Experimental animals
Ten- to 12-week-old (n = 44) SD rats weighing 250–300 g were used. The parent stock of rats was purchased from SPF Laboratory Animal Company (Koatech, Pyungtaek City, Korea). The rats were reared in a strict hygienic, climate-controlled environment and provided with sufficient food and water. The rats were sheltered, reared, and fed following the standard humane protocols accepted by the Chonbuk National University, Jeonju, South Korea.
Bacterial strain
Pathogenic B. abortus biotype 1 originating from cattle in Korea was used for experimental infection in SD rats. B. abortus biotype I lyophilized stock culture were picked up from the laboratory stock. Brucella was inoculated into the Brucella agar media (Difco, Kansas City, USA) and incubated at 37°C for 5–7 days under 5% CO2. The grown bacterial culture was collected in normal saline.
Experimental infection
Rats (n = 40) were inoculated intraperitoneally with 0.1 mL saline containing 1 × 1010 colony-forming units (CFU)/mL of B. abortus biotype 1. Four rats were inoculated intraperitoneally with sterile saline as uninfected control. Infected and uninfected control rats were reared separately.
Clinical examinations
Rectal temperature, food and water intake in addition to other abnormal clinical signs such as chills, weakness, and lameness of all infected and control rats were noted daily for 2 weeks.
Collection of specimens
Blood samples were collected by aseptic cardiac puncture from the inoculated rats at 0, 3, 7, 14, 21, 28, 35, 42, 60, 90, and 120 days after infection under general anesthesia induced by intraperitoneal administration of 10 mg/kg of tiletamine and zolazepam (Zoletil 50; Virbac Laboratories, Carros, France). Blood samples were collected in vacuum tubes with or without anticoagulant. All the blood samples were processed within 1 h of being drawn. The rats were then killed and spleen, liver, and kidney were aseptically removed for bacteriological studies. Spleens were weighted by an electronic balance (A & D Company, Tokyo, Japan) and observed for splenomegaly. Blood samples with anticoagulant (ethylenediaminetetraacetic acid, EDTA) were also examined for bacteriological studies. Blood samples without anticoagulant were allowed to clot for 6 h at 25°C before centrifugation. Serum samples were collected and stored at −20°C until tested. All other samples were stored at 4°C until cultured.
Bacteriological analysis
The EDTA-blood samples was exposed to osmotic shock together with distilled water and centrifuged at 3000 rpm for 60 min at room temperature. The following blood pellet was reconstituted in Brucella broth. Then, 50 μL of suspension was immediately inoculated in duplicate onto Brucella agar media and incubated at 37°C for 5–7 days in a 5% CO2 atmosphere. Bacterial growth was counted 15 days after inoculating. Kidney, liver, and spleen samples were mashed individually in a lab blender (Masticator Homogenizator; IUL Instruments, Barcelona, Spain). For the isolation of Brucella, mashed samples were cultured duplicate onto Brucella agar media containing antibiotics (cycloheximide, bacitracin, and polymyxin B) that inhibit the growth of bacteria other than Brucella. Inoculated media were incubated at 37°C for 5–7 days in a humid atmosphere enriched with 5% CO2. The identification of the culture-positive isolates in the specimens was performed by standard methods as narrated previously (Alton et al. 1988).
Determination of cytokine levels
At 0, 3, 7, 14, 21, 28, 35, 42, 60, 90, and 120 days postinfection, rats were killed and spleens were aseptically removed. Approximately one-third of spleen was weighed and homogenized in phosphate-buffered saline and an aliquot of the resulting cell suspension was plated to determine the number of CFU. The remaining two-thirds of the spleens were weighed, minced, and used to prepare spleen cell suspensions. Cytokine expression was evaluated in culture supernatants of splenocytes energized with 2 μg CBP/well. Spleen cells were preserved in duplicate in 24-well round bottom microplates at 37°C in an atmosphere with 5% CO2. Cells were adjusted to 2 × 106 cells/mL in Roswell Park Memorial Institute medium in a final volume of 1 mL. Supernatants were collected on 72 h after antigen stimulation and stored in multiple small aliquots at −80°C until they were needed. The total IFN-γ and IL-10 concentrations in the sera and spleen cells supernatant of B. abortus-infected rats were measured by commercial ELISA kits for IFN-γ (eBioscience, San Diego, CA, USA) and IL-10 (R&D Systems, Inc., Minneapolis, MN, USA). The tests were conducted according to the instructions of the manufacturer.
Statistical analysis
The variances in cytokine production between two groups were analyzed using Student's t-test. A value of p < 0.05 was considered statistically significant.
Results
Clinical observations
All the infected rats showed torpid, anorectic, and febrile illness within 24 h. The uninfected control rats did not exhibit any of the abnormal clinical signs. The highest rectal temperature was 38.30 ± 0.152°C in the infected group and 36.5 ± 0.05°C in the uninfected control group.
Distribution of B. abortus biotype 1 infection
B. abortus biotype 1 was identified from culture of spleen at 7 days postinfection and persisted all over the experiment. B. abortus was also isolated from liver and kidney at 7 days postinfection and detected in those tissues up to 42 days postinfection. Bacteria were isolated from blood at 3 days postinfection and remained in the blood up to 35 days postinfection. At 35 days postinfection, no bacteria were isolated from the blood cultures throughout the experiment. The result of B. abortus biotype 1 recovery from blood, spleen, liver, and kidney at different days postinfection is given in Table 1.
Summary of Brucella abortus Biotype 1 Isolation from Blood, Spleen, Liver, and Kidney of Sprague-Dawley Rats Experimentally Inoculated with Brucella abortus Biotype 1
n, Number of rats from which Brucella abortus was isolated.
Percentage of B. abortus isolated from the tissue in culture-positive animals.
Serum IFN-γ level
The IFN-γ was detected in the sera of B. abortus-infected SD rats at 3 days postinfection (81.75 ± 40.87 pg/mL). The IFN-γ reached the maximal level at 7 days postinfection (127.15 ± 63.57 pg/mL). Then the level of IFN-γ gradually declined to the end of the study. At 60 days after infection, the level of IFN-γ was recorded as 5.64 ± 2.82 pg/mL. During subacute stages of infection, the serum IFN-γ levels were almost the same level compared with the uninfected control rats. The IFN-γ levels in the sera of B. abortus-infected rats are given in Fig. 1.
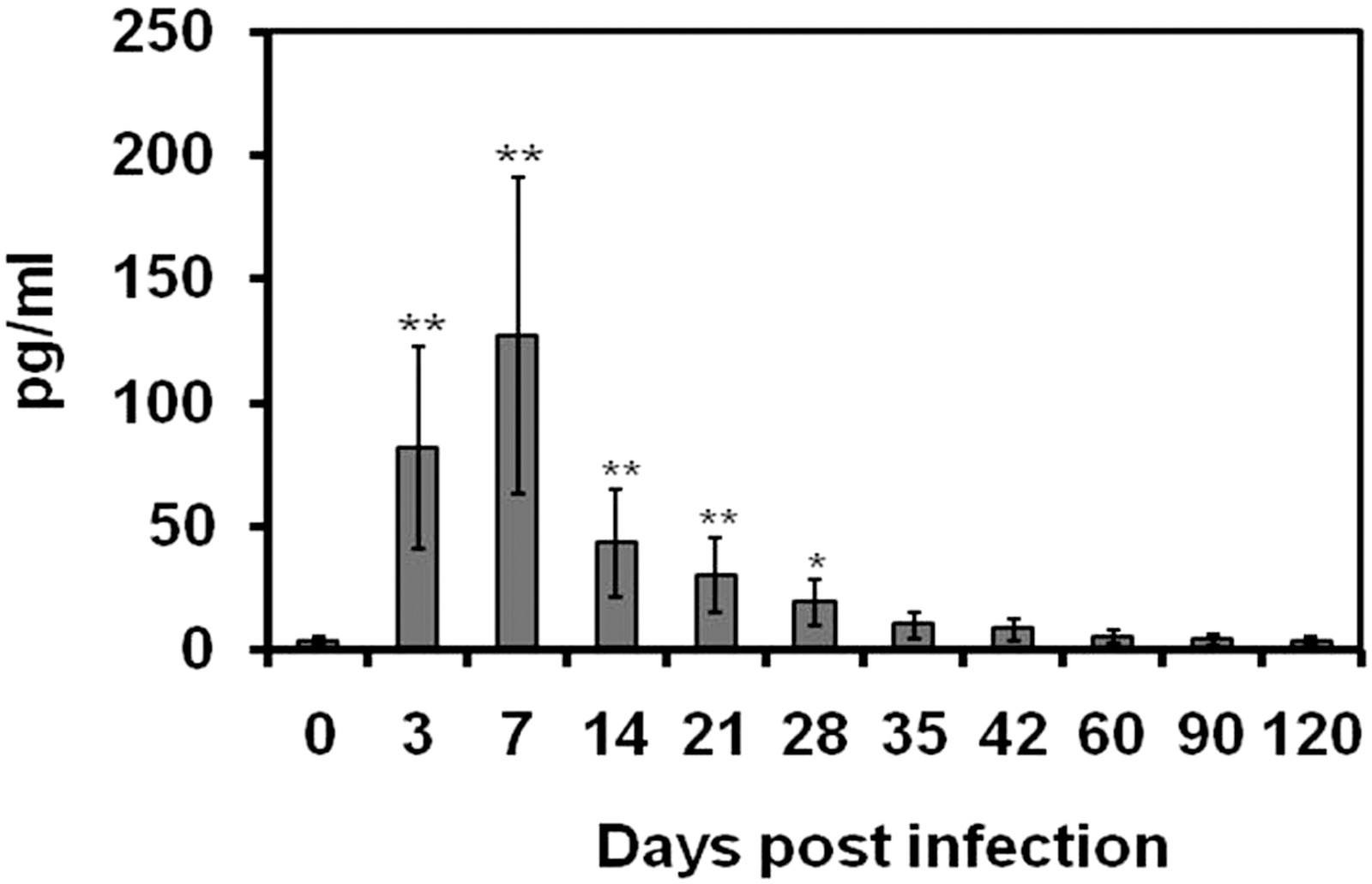
Total serum IFN-γ of rats at 0, 3, 7, 14, 21, 28, 35, 42, 60, 90, and 120 days after infection with Brucella abortus biotype 1. The levels of serum IFN-γ are expressed as mean value of 4 rats ± SD. Statistically significant differences of IFN-γ levels between control and infected rats at different time points of infection are indicated by asterisks (*p < 0.05 and **p < 0.001). IFN-γ, interferon gamma; SD, standard deviation.
Serum IL-10 level
The IL-10 was detected in the sera of B. abortus-infected SD rats as early as 3 days postinfection (11.29 ± 5.64 pg/mL). The IL-10 levels gradually increased from 3 days postinfection and reached the maximal level at 28 days postinfection (79.63 ± 39.81 pg/mL). Then the level of IL-10 gradually declined until the end of the study. At 60 days postinfection, the level of IL-10 was recorded as 10.23 ± 5.11 pg/mL. During subacute stages of infection, the serum IL-10 levels were almost the same level compared with the uninfected control rats. The IL-10 levels in the sera of B. abortus-infected rats are given in Fig. 2.
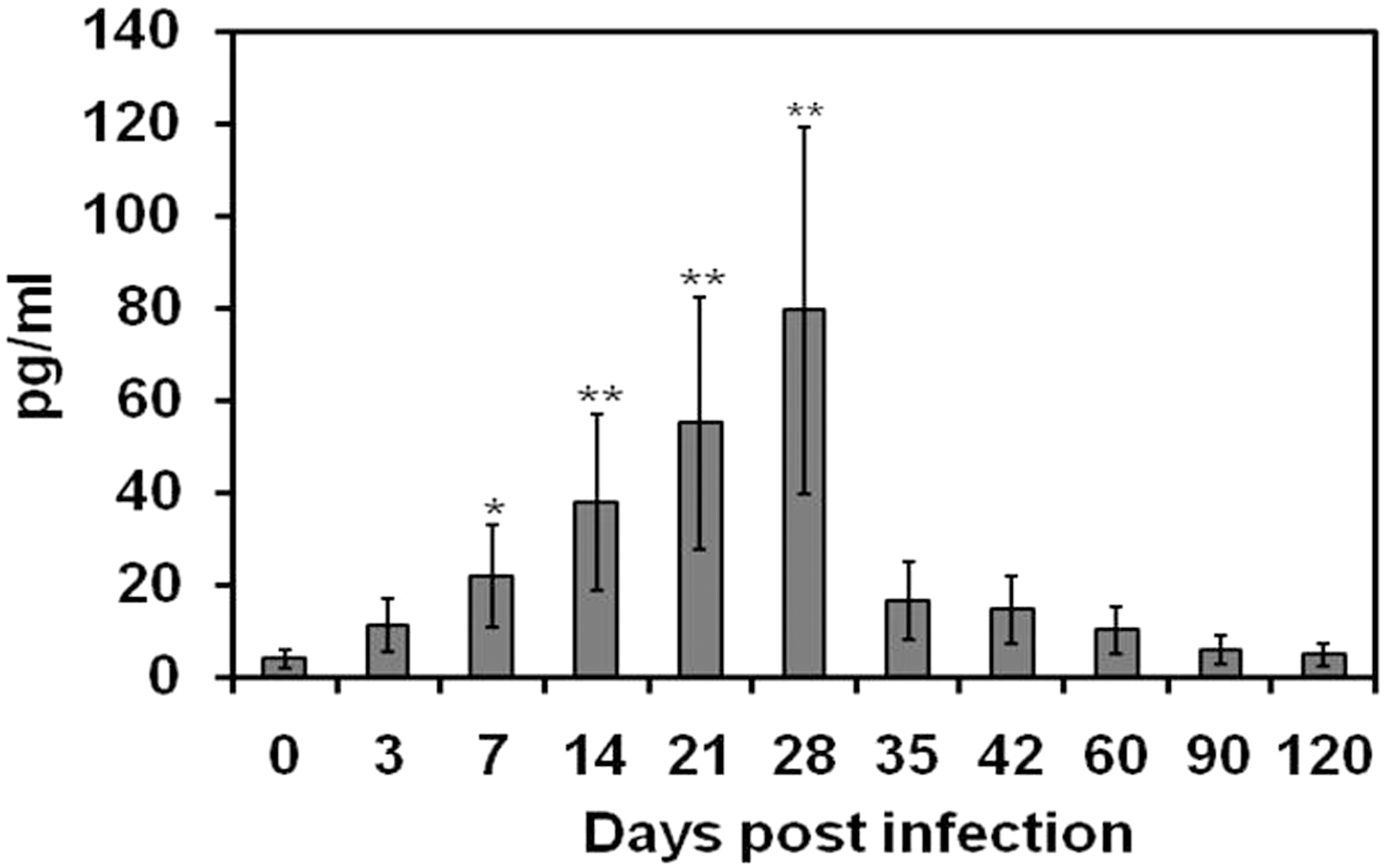
Total serum IL-10 of rats at 0, 3, 7, 14, 21, 28, 35, 42, 60, 90, and 120 days after infection with Brucella abortus biotype 1. The levels of serum IL-10 are expressed as mean value of 4 rats ± SD. Statistically significant differences of IL-10 levels between control and infected rats at different time points of infection are indicated by asterisks (*p < 0.05 and **p < 0.001). IL-10, interleukin-10.
In vitro IFN-γ production by spleen cells
The IFN-γ was detected in the spleen supernatant of rats at 3 days postinfection (164.2 ± 82.1 pg/mL). The highest level of IFN-γ in the spleen supernatant was recorded at 7 days postinfection (483.31 ± 221.63 pg/mL). Then the IFN-γ level gradually declined until the end of the experiment. Very low level of IFN-γ was marked at 90 days postinfection (8.65 ± 4.32 pg/mL) and IFN-γ level reached almost base level at 120 days postinfection (5.64 ± 2.82 pg/mL) compared with the uninfected control rats (4.98 ± 2.49 pg/mL). The results of IFN-γ level in the spleen supernatant of rats at different time points of infection are given in Fig. 3.

In vitro IFN-γ of rats at 0, 3, 7, 14, 21, 28, 35, 42, 60, 90, and 120 days after infection with Brucella abortus biotype 1. Results are expressed as mean value of 4 rats ± SD. Statistically significant differences of IFN-γ levels between control and infected rats at different time points of infection are indicated by asterisks (*p < 0.05 and **p < 0.001).
In vitro IL-10 production by spleen cells
At 3 days postinfection, IL-10 level was detected in the spleen supernatant (191.3 ± 95.65 pg/mL). The IL-10 level gradually increased from 3 days postinfection and reached the maximal at 14 days postinfection (443.01 ± 221.5 pg/mL). Then IL-10 level gradually declined until the end of the study. The IL-10 level in the spleen supernatant at 90 and 120 days postinfection was recorded as 12.27 ± 6.13 and 6.95 ± 3.47 pg/mL, respectively. The results of IL-10 level in the spleen supernatant at different time points of infection are given in Fig. 4.

In vitro IL-10 of rats at 0, 3, 7, 14, 21, 28, 35, 42, 60, 90, and 120 days after infection with Brucella abortus biotype 1. Results are expressed as mean value of 4 rats ± SD. Statistically significant differences of IL-10 levels between control and infected rats at different time points of infection are indicated by asterisks (*p < 0.05 and **p < 0.001).
Discussion
It is familiar that lymphoid tissue of the susceptible host is the predilection site for B. abortus (MacMillan et al. 1982) that localizes at first in the regional lymph node and then disperses hematogenously to the organ of the reticuloendothelial system (Memish et al. 2000). The data of bacteriological studies suggest that after experimental infection, B. abortus disseminated to blood and various lymphatic tissues. Variation of persistence of B. abortus was seen in blood and among lymphoid organs at different times of infection. The existence of B. abortus was seen in blood, kidney, and liver up to 35, 42, and 42 days postinfection, respectively. Young et al. (1979) isolated great numbers of living bacteria from the tissues of B. abortus-infected mice 30 days postinfection. In this study, existence of B. abortus was seen in the spleen over the 16-week study period. These results are in agreement with High et al. (2007) who isolated B. abortus 2308 bacteria from spleen in mice at 5 and 8 weeks postinfection. Damir et al. (1989) also isolated B. abortus from the spleen but they did not find B. abortus from the kidney in female camel after experimental infection.
Presence of B. abortus was noted in spleen until 20 weeks in mice and 6 months in wolves (Stevens et al. 1994, Tessaro and Forbes 2004). It has been reported that Brucella organisms may still be found from the lymph node and spleen of mice after 6 months of infection (Bosseray et al. 1982, Sangari et al. 1988), suggesting that virulent Brucella might persist in mice for life. In case of blood, presence of B. abortus was noted until 5 weeks of infection in this experiment. Garcia-Carrillo (1990) observed an impermanent bacteremia in the laboratory-infected mice and the bacteria were gradually removed few weeks after the beginning of the infection. The continuous presence of B. abortus in the spleen in this study showed that it is the most preferable colonization site in rats compared with blood, liver, and kidney.
Cytokines seem to have a key role in the pathogenesis of brucellosis and the Th1/Th2 balance may be engaged in the sensitivity or resistance to the disease (Pasquali et al. 2001, Galanakis et al. 2002). Some intracellular bacteria, including Brucella spp. and Mycobacterium tuberculosis, display Th1-type responses during cell proliferation and cytokine production (Cowie et al. 2014). A Th1 response is necessary for accomplishment of the elementary infection caused by Brucella and the essential aspect of this response appears to be IFN-γ production (Mielke et al. 1998). IFN-γ is necessary for the removal of Brucella (Zhan and Cheers 1993). On the contrary, the Th2 cytokine, IL-10, can inhibit macrophage function and increase susceptibility to infection (Fernandes and Baldwin 1995). IL-10 is essential for regulation of the immune responses (Cyktor and Turner 2011). Moore et al. (2001) stated that IL-10 is a powerful anti-inflammatory cytokine associated with macrophages that plays a central role in limiting host immune response to pathogens.
A statistically significant higher serum level of IFN-γ was recorded at 3 and 7 days postinfection (p < 0.001). The mean serum levels of IFN-γ and TNF-α and the frequency of Th1 cells were significantly higher in the brucellosis patients in comparison with healthy subjects (p < 0.05) (Xu et al. 2019). On the contrary, statistically significant higher serum IL-10 levels were recorded at 21 and 28 days postinfection compared with the serum IFN-γ levels (p < 0.001). Akbulut et al. (2007) recorded statistically significant IFN-γ level in the sera of humans during acute brucellosis. Paul et al. (2018) found that patients with brucellosis have significantly higher levels of circulating Th1 cells and cytokines. Higher serum IFN-γ level was also recorded by Demirdag et al. (2003). It was reported that CD4+ IFN-γ+ lymphocytes play a key role in containing Brucella infection in cattle (Skendros and Boura 2013). In this study, the IFN-γ levels in the spleen supernatant were significantly increased at 7 and 14 days postinfection compared with IL-10. Shim et al. (2020) found significantly higher IFN-γ and IL-4 in the spleen of BALB/c mice immunized with B. abortus proteins. Increased levels of IFN-γ and TNF-α have been detected in the spleens and serums of mice 7 days after being infected with the Brucella bacteria (Corsetti et al. 2013, Reyes et al. 2016).
IFN-γ may be one of the central proinflammatory cytokines that promote resistance to Brucella infection. On the contrary, IL-10 responses were found to be significantly increased at 21, 28, 35, and 42 days postinfection compared with IFN-γ. These findings are in agreement with Fernandez-Lago et al. (1996) who recorded simultaneously significant production of IFN-γ and IL-10 in the spleens of mice between the second and eighth days after infection with B. abortus 2308 strain. The in vivo and in vitro serum cytokine profiles in this study showed that B. abortus infection in rats primarily educe Th1-dominant immune response accompanied by Th2-dominant immune response. Th1 cytokines (IL-2 and IFN-γ) and Th2 cytokines (IL-4 and IL-10) are mostly studied in the B. abortus infection. Although we have not screened all Th1 and Th2 cytokines profiles in this study, most of the investigators pointed out that IFN-γ is mainly responsible for Th1-dominated immune response in B. abortus infection.
The IL-2 cytokine stimulates T lymphocytes to produce IFN-γ (Kasahara et al. 1983). Therefore, we have screened only IFN-γ for measuring Th1-dominated immunity. On the contrary, IL-4 helps B cell to enhance the production of IL-10 and skew the immune response toward Th2 cells (Yao et al. 2005). Therefore this study only screened IL-10 to understand Th2-dominant immunity in experimental B. abortus infection in rats. However, the identification of T cell activation maker needs further study to better understand how B. abortus infection impacts on T cell differentiation. In our previous studies, we found significantly dominant IgG2a response (indicative of a Th1 response) over IgG1 response (indicative of Th2 response) (p < 0.001) (Khatun et al. 2009).
Intracellular pathogens cause chronic infection by downregulating Th1 cytokines (IFN-γ or IL-2) and upregulating Th2 cytokine production (IL-4 or IL-10) (Fernandes et al. 1995), which ultimately results in IgG1 antibody production (Finkelman et al. 1990). In mouse, IFN-γ helps transforming immunoglobulin to IgG2a (Golding et al. 2001). A switching of IgG1 and IgG2a immunoglobulin isotypes with the production of IFN-γ and IL-10 cytokines was noticed in this experiment. Data in this study indicated that SD rats infected with B. abortus biotype 1 elicit Th1-dominant immune response in acute infection and Th2-dominant immune response in chronic infection.
Footnotes
Authors' Contributions
M.M.K. and M.A.I. prepared the article; M.M.K. and M.A.I. conducted an animal experiment, collected and processed the samples and conducted diagnostic tests; B.K.B. reviewed the article. The article has been read and approved by the all authors.
Acknowledgments
This study was a part of the PhD research work at the Chonbuk National University, Jeonju, South Korea. The authors thank the Islamic Development Bank (IDB), Saudi Arabia for funding the research. The authors thank and acknowledge Dr. Stephen M. Boyle from Virginia Tech, USA for editorial comments.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This work was supported by the Islamic Development Bank, Saudi Arabia (Grant no. 4/BD/P27).