
Abstract
Background:
Emergence of mosquito-borne arboviruses has caused significant public health burden. The life cycle of arboviruses comprises sylvatic and urban cycles, including a wildlife reservoir, a human host, and an arthropod vector. However, many questions remain on the sylvatic cycles of arboviruses. In this study, we investigate the prevalence of IgG antibodies to arboviruses of public health importance in African bats.
Material and Methods:
We collected dried blood spots from bats in Cameroon, Guinea, and the Democratic Republic of the Congo (DRC). To detect IgG antibodies to 10 antigens of 6 arboviruses (Dengue, Zika, West Nile, Usutu, Chikungunya, and O'nyong nyong viruses), we adapted a previously validated multiplex detection assay based on the Luminex technology.
Results:
We tested samples from 2579 bats, representing 1917 frugivorous and 641 insectivorous bats distributed in 7 families and 21 species. Overall, 218/2579 (8.45%) bat samples reacted with at least 1 of the 10 antigens tested. The highest prevalence was observed against Usutu virus with 2.3% (59/2579), followed by 1.9% (49/2579) and 1.35% (35/2579) for the Dengue virus serotypes 4 and 3, respectively. The global seroprevalence varied by country and collection site: 11% (151/1376) in Cameroon, 3.5% (20/565) in DRC, and 7.3% (47/638) in Guinea. The highest rates were observed in Hypsignathus monstrosus (17.9%), Rousettus aegyptiacus (16.4%), and Eidolon helvum (10.7%), and in species from the insectivorous Molossidae family (7.8–8.9%). Finally, we observed changes in seroprevalence over the year in E. helvum and H. monstrosus colonies, which could be related to population structure.
Conclusion:
On more than 2500 bat samples tested, we showed variable IgG seroprevalences against multiple arboviruses. Overall, the prevalence of IgG antibodies of 8.45% against arboviruses found in bats suggest that they could play a role in arboviruses cycles in the wild, in addition to other animal species.
Introduction
The number and impact of emerging infectious diseases (EIDs) has significantly increased over time. The large majority of EIDs have a zoonotic origin, and >70% originate from wildlife (Jones et al. 2008). Among them, mosquito-borne arboviruses constitute a significant threat to public health in worldwide. Arboviruses are predominantly RNA viruses and approximately one third have pathogenic potential for humans, such as Dengue, Zika, West Nile, Usutu, Chikungunya, O'nyong-nyong, and many other viruses (Guzmán et al. 2020). The life cycle of arboviruses comprises in general a sylvatic or enzootic cycle and an urban cycle that include a wildlife reservoir, a human host, and an arthropod vector in a complex interplay.
Many arboviruses causing epidemics today have their origin in Africa or were first identified in Africa; that is, Yellow Fever, West Nile, Zika, Chikungunya, and Usutu viruses (Dick 1953, Mason and Haddow 1957, Shope 1994). For many of them, such as Zika, Dengue, or Chikungunya, nonhuman primates (NHPs) were considered to play a major role in sylvatic cycles, but this was based on limited data. We recently showed that the observed seroprevalence for arboviruses on a large number of NHPs in Africa was not sufficient for them to be considered as the main reservoir species. Thus, other species most likely also play a role to maintain these viruses in the sylvatic cycles in Africa (Valentine et al. 2019, Raulino et al. 2021).
Bats are known to harbor many viruses and arboviruses have also been detected in bats (Luis et al. 2013, Melaun et al. 2014, Calderon et al. 2016, Fagre and Kading 2019, Ayhan et al. 2021). Bats represent 20% of all mammalian species, with >1400 species worldwide, have high population densities and roosting behavior that favor the likelihood of transmission of pathogens (Omatsu et al. 2007). The majority of studies reporting on arboviruses in bats are from southern America (Ubico and McLean 1995, Thompson et al. 2015, Hernández-Aguilar et al. 2021). Very few studies have been conducted in Africa, the majority were done before 1970 and reported antibodies to Zika virus or Yellow Fever in several bat species in East Africa (Williams et al. 1964, Simpson et al. 1968).
The limited data available on this important question are probably due to lack of assays for bat's immune typing, the difficulty of arbovirus serology because of antigenic proximity among different arboviral lineages, and limited sample volumes that can be drawn from bats. Recently, our group has developed a high-throughput serological tool to detect IgG antibodies to different arboviruses of global interest that circumvent some of the aforementioned difficulties (Raulino et al. 2021). In the present report, we adapted this serological tool for the detection of IgG antibodies to six arboviruses (Chikungunya, O'nyong nyong, Dengue, Zika, West Nile, and Usutu viruses) and tested bat samples collected in Cameroon, Guinea, and Democratic Republic of the Congo (DRC).
Our data showed that seroprevalence varied by country, by collection site, by bat families and species, and ranged from 1% to 20%. These findings suggest that bats could play a role in sylvatic life cycles of some arboviruses.
Materials and Methods
Bat sampling and sites
Samples from free-ranging frugivorous and insectivorous bats were collected between 2016 and 2019 from in Cameroon, DRC, and Guinea in the framework of a study on Ebola and other viruses in bats. Bats were captured using mist nets or harp traps in roosting and foraging sites and immediately released after sampling as previously described (De Nys et al. 2018). Dried blood spots (DBS) were prepared on Whatman 903 filter paper (GE Healthcare, Feasterville-Trevose, PA) from whole blood samples collected by venipuncture of the propatagial or brachial vein. The study received approvals of adminsitrative authorities from the three countries where it was conducted.
DBS samples were preserved as described in De Nys et al. (2018), stored in the field at ambient temperature and then at −20°C. For each sample, information on capture site (GPS coordinates [Supplementary Table S5], ecological environment), capture method, morphology (body measurements, weight, and color), sex, age class (adult and juvenile), reproduction cycle, and visual species identification were recorded in the field. Permissions to perform this research were obtained from administrative authorities of Cameroon, Guinea, and the DRC.
Detection of antibodies to arboviruses
Samples were tested with our previously developed assay based on the Luminex technology with sensitivity and specificity and accuracy >95% on a reference panel of well-documented human samples (Raulino et al. 2021). We used 10 commercially available recombinant proteins from 6 arboviruses: CHIKV_E2 (Chikungunya virus Envelope protein 2), ONNV_E2 (O'nyong nyong virus envelope protein 2), DENV_NS1 (Dengue virus-nonstructural protein 1; 1–4 serotypes), USUV_NS1 (Usutu virus-nonstructural protein 1), WNV_NS1 (West Nile virus nonstructural protein 1), WNV_DIII (West Nile virus domain-3 protein), and ZIKV_NS1 (Zika virus nonstructural protein 1) (Raulino et al. 2021).
The assay was performed as previously described and optimized for bats using goat anti-bat biotin–labeled IgG (Euromedex) and Streptavidin-R-phycoerythrin (Fisher Scientific/Life Technologies, Illkirch, France) (Raulino et al. 2021). Whole blood was reconstituted from DBS as previously described and a final plasma dilution 1/200 was used (Ayouba et al. 2019). The results were expressed as median fluorescence intensity (MFI) per 50 beads. Each plate included a blank well, positive samples for Chikungunya, Dengue, Zika, and West Nile viruses, and also two negative samples to validate interassay repeatability.
Calculation of cutoff values for the different antigens
In the absence of positive and negative control samples from bats, we analyzed our data with different statistical methods to determine MFI cutoff values for each antigen as previously described (Gilbert et al. 2013, Peel et al. 2013; De Nys et al. 2018, Ayouba et al. 2019; Raulino et al. 2021). We used a change-point analysis with the R package “changepoint” and calculated one single shift in the arithmetic mean with the AMOC (at most one change) method (Killick and Eckley 2014, Lardeux et al. 2016). We also used fitted univariate distributions (binomial and negative exponential distributions) to our data and defined the cutoff based on a 0.001 risk of error. Data were bootstrapped 10,000 times and averaged for each antigen. Analyses were done with R software version 3.3.6. We then compared the cutoff values identified by the three different methods and calculated their mean as a consensus cutoff that we used in this study (Supplementary Table S1). A sample was considered reactive if MFI was above the cutoff value.
Molecular confirmation of bat species
Species identification recorded in the field was molecularly confirmed on a subset of samples by sequence analysis of an 800 bp fragment of the mitochondrial cytochrome b (CytB) like in our previous studies (De Nys et al. 2018, Lacroix et al. 2020). Sequences were submitted to NCBI for BLAST analysis to identify the most similar bat species. For samples with none or low similarity (<97%) hits with species in GenBank, phylogenetic tree analysis was done using maximum likelihood methods implemented in PhyML (Guindon and Gascuel 2003) with reference sequences to approach genus identification.
To discriminate between Epomophorus gambiensis and Micropteropus pusillus, we used also morphological details on forearm and weight measurements because species identification was not possible based on CytB sequences (Nesi et al. 2011). Species identification was extrapolated by per capture method, GPS coordinate, and morphological descriptions for the other samples.
Results
Distribution and diversity of bat species
We tested a total of 2579 bats captured in 21 different sites in Guinea (n = 8), Cameroon (n = 4), and DRC (n = 9) (Fig. 1). The samples were from bats living in urban areas (52%), villages (26%), forest (10.5%), and caves (10.9%). For 2313 (89.6%) samples, the species identification in the field was confirmed by sequence analysis; however, it was not possible to clearly discriminate between certain Chaerophon and Mops species from the Molossidae family based on CytB sequences and morphological data. We tested at least 12 different species of frugivorous bats, and 6 different families of insectivorous bats. Details on the different bat species that were collected in each country are shown in Table 1.
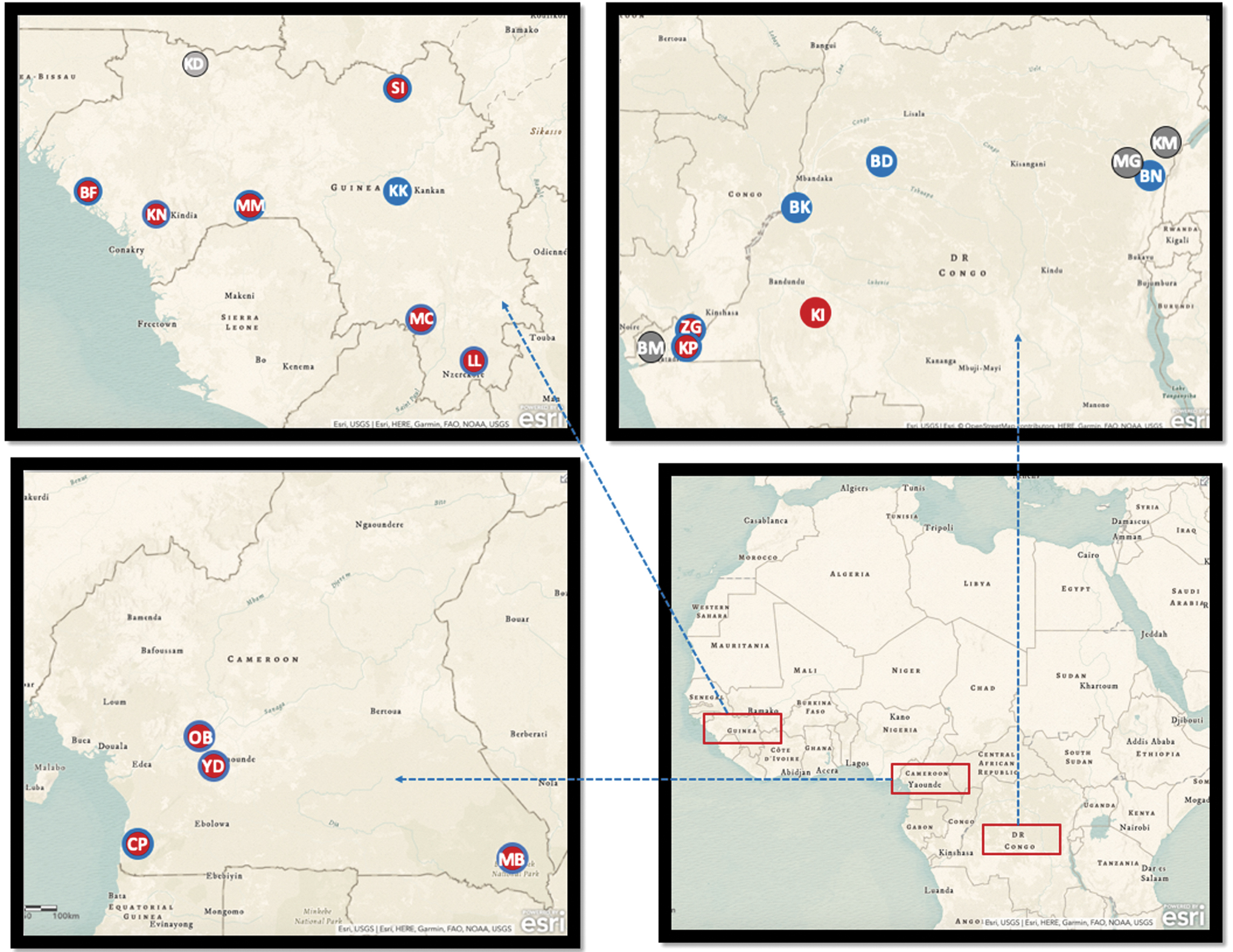
Bat collection sites and presence of arboviruses in Cameroon, Guinea, and DRC. The maps represent Africa and the three countries (Guinea, top; Cameroon, center; and the DRC, bottom) where studied samples have been collected. Gray dots indicate sampling sites where no antibodies to arboviruses were detected; red dots, sites where antibodies to alphaviruses were detected; blue dots, sites where antibodies to flaviviruses were detected; red dots with blue circles, sites were antibodies to flavi- and alpha viruses were detected. Site abbreviations: Cameroon—Village: CP/Campo; MB/Mambele, Urban area: OB/Obala; YD/Yaoundé. DRC—Caves: BM/Boma; KP/Kimpse; ZG/Zongo, Forest: ZG/Zongo; BD/Boende, Urban area: BN/Beni; BK/Bikoro; BM/Boma; KP/Kimpse; KM/Komanda; MG/Mangina, Village: BD/Boende; KI/Kikiwit. Guinea—Caves: KK/Kankan; KN/Kindia; MC/Macenta; MM/Mamou; SI/Siguiri, Forest: MC/Macenta, Urban area: KN/Kindia; MM/Mamou; Villages: BF/Boffa; KK/Kankan; KN/Kindia; KD/Koundara; LL/Lola; MC/Macenta; MM/Mamou; SI/Siguiri. DRC, Democratic Republic of the Congo.
Number of Different Bat Families and Species Collected in Each Country
In Cameroon and DRC, Epomops franqueti, in Guinea Epomops buetikoferi.
DRC, Democratic Republic of the Congo.
Overall, 1193 bats were male, 1381 female, and for 5 data were not available. Among females, 77% were adults, 8.0% were reproductively immature, and 14% were juveniles. For males, 70%, 12%, and 17% were adults, reproductively immature, and juveniles, respectively. For the remaining samples information on age and sex were not available.
Evidence of antibodies to Alphaviruses and Flaviviruses in bats
Antibodies to alphavirus (Chikungunya and O'nyong-nyong) and flaviviruses (Dengue 1–4, Usutu, West Nile and Zika) tested in this study were observed in Eidolon helvum, Epomophorus gambianus, Epomops sp., Hypsignathus monstrosus, Lissonycteris angolensis, M. pusillus, Myonycteris torquarta, Rousettus aegyptiacus, Hipposideros sp., Miniopterus sp., Chaerephon sp., Mops sp., and Rhinolophus sp. bats from the three countries (Fig. 1). Overall, 8.45% (218/2579) of the samples reacted with at least one arbovirus antigen (Table 2). Globally, 2.3% of the bats had antibodies to Usutu virus, 0.85–1.35% to serotypes 1–4 of Dengue virus and the lowest proportions were observed for Zika virus (0.54%).
Prevalence of Antibodies to Alphaviruses (Chikungunya Virus and O'Nyong Nyong Virus) and Flaviviruses (Zika Virus, Dengue Virus 1–4, Usutu Virus, and West Nile Virus) in Bats in the Different Collection Sites Shown in Fig. 1
Total means number of samples reactive with at least one antigen.
CHIKV_E2, Chikungunya virus Envelope protein 2; CI, confidence interval; DENV_NS1, Dengue virus nonstructural protein 1; ONNV_E2, O'nyong nyong virus envelope protein 2; USUV_NS1, Usutu virus-nonstructural protein 1; WNV_DIII, West Nile virus domain-3 protein; WNV_NS1, West Nile virus nonstructural protein 1; ZIKV_NS1, Zika virus nonstructural protein 1.
Global rates and stratified per virus varied among countries and within countries. In Cameroon, 11% (151/1376) of bat samples had antibodies to arboviruses, ranging between 0.58% for Zika virus to 4.0% for Usutu virus. In Guinea, we also observed antibodies to all viruses with a total seroprevalence of 7.3% (47/638) and a predominance to O'nyong-nyong (2.35%). In DRC, 3.5% (20/565) of the bat samples tested had antibodies with a predominance to antigen NS1 of West Nile virus, and in contrast to the two other countries, no antibodies to Chikungunya virus were observed.
Antibodies to at least one arbovirus were observed in 14 (70%) of the 21 species tested, that is, in all species with sufficient samples (>20) (Table 3). The highest rates were observed among the following frugivorous bat species, Hypsignathus monstrosus (17.9%), R. aegyptiacus (16.4%), and E. helvum (10.7%) and in species from the insectivorous Molossidae family, Mops sp. (8.9%), and Mops/Chaerophon sp. (7.8%). E. helvum is a common species, which showed IgG antibodies against all viruses tested, but with different positivity rates for each virus in different sites tested, which is probably related to differences in the numbers of samples tested.
Seroprevalence of IgG Antibodies to Arboviruses in Bat Species
Number positive with at least one antigen.
In Cameroon and DRC, Epomops franqueti, in Guinea Epomops buetikoferi.
CI was not calculated.
p = 0.02825.
H. monstrosus is also a common species, but large numbers were only tested in Cameroon, and antibodies were observed against all viruses tested with 10% of samples reactive to Usutu virus. In R. aegyptiacus, which were highly represented in the sample set from Guinea, antibodies against almost all viruses were detected reaching 8.9% for O'nyong-nyong virus. Also, in bats from the Molossidae family, antibodies against almost all alphaviruses and flaviviruses were observed with 7.4% of samples reactive against to serotype 4 of Dengue in Cameroon.
In other species, for example M. pusillus, IgG antibodies were only observed against flavivirus, and in Myonycteris torquata only IgG antibodies to alphavirus were detected (Table 3). Overall, antibodies were observed in 10% (192/1917) of frugivorous bats versus 3.9% (25/641) in insectivorous bats. Frugivorous bats had antibodies to all viruses tested, and in insectivorous bats no antibodies were observed against the Usutu and to antigen NS1 of West Nile virus (Table 3). Only for the serotype 2 of Dengue virus, the difference between the positivity in frugivorous and insectivorous was statistically significant (p = 0.028; odds ratio: 4.5626; 95% confidence interval: 1.41–39.7).
Ten percent of frugivorous and insectivorous bats captured in urban areas had antibodies to arboviruses versus 6.6% in villages. E. helvum bats were captured in cities, villages, and forests with the following positivity rates, 10.7%, 18%, and 9.8%, respectively. Similar rates were observed for H. monstrosus captured in urban areas (18.5%) and villages (16.6%). R. aegyptiacus bats were also positive in all settings with rates between 11.5% and 30%. Most insectivorous bats were captured in more remote areas and showed lower rates, except for species of the Molossidae family, with 7.7% of Chaerephon/Mops, which were captured in villages having antibodies to arboviruses (Table 4).
Proportion of Samples with Antibodies to Arbovirus for Each Species According Ecological Site Where They Were Captured
In Cameroon and DRC, Epomops franqueti, in Guinea Epomops buetikoferi.
The difference in positivity rates of antibodies to arboviruses between frugivorous and insectivorous might reflect the different biotopes. Supplementary Table S2 shows in detail the reactivity with the different viruses according to the ecological settings independently from the species. Overall, antibodies to almost all viruses studied are observed in all settings, although at different rates with a trend of lower rates in caves as compared with the three other settings.
Antibodies in E. helvum and H. monstrosus according to seasons, age, and reproductive cycle
The 815 E. helvum and 227 of the 249 H. monstrosus samples in Cameroon were part of colonies roosting in Yaoundé and foraging in neighboring areas. Samples were collected on a monthly basis between December 2018 and November 2019. For E. helvum, we tested 221 juveniles, 218 reproductively immature and 378 adult bats, and comparable seroprevalence rates were observed in all age categories ranging from 10.1% to 11.1%. For H. monstrosus, differences according to age category were observed, with highest proportion of positives among adults (31/132, 23.5%), 10/72 (13.9%) of juveniles, and lowest proportions in reproductively immature (1/22; 4.6%) (Fig. 2).

Percentage of positive samples in colonies of bats, according to age and reproductive cycle. The figure represents the proportions of positive sample in two species (Hypsignathus monstrosus and Eidolon helvum) of fruit bats followed up over a year in Yaoundé, Cameroon.
Seropositivity rates were also significantly higher in adult H. monstrosus as compared with adult E. helvum bats, 23.5% versus 11.1%, respectively (p < 0.0001) (Supplementary Table S3). We also examined whether antibody status can vary in females during gestation and lactation. Among the female H. monstrosus bats, 2/11 (18.1%) of the gestating bats were positive versus 28.5% (18/63) of the other adult female bats, and for E. helvum the opposite trend was observed with 18.1% (2/11) gestating bats positive versus 9.3% (15/160) for the other adult female bats.
The differences observed were not significant statistically. Similarly, there were also differences between female adults during lactation period, as shown in Fig. 2, but they were also not statistically significant. Taken together, the data from the E. helvum and H. monstrosus bat colonies showed fluctuations over time with a minimum rate in June (4.26%) and maximum in February (34.78%) (Supplementary Table S4); however, the number of samples are low for certain months and the population structure at a given period could influence positivity rates as shown earlier.
Simultaneous reaction to multiple alphavirus and flavivirus in bats
We observed simultaneous reactivity to antigenically related viruses, but also to genetically diverse and less antigenically related viruses (Table 5). The highest simultaneous reactivity was observed for NS1 antigens of the four Dengue virus serotypes, reaching 68% between the serotypes 1 and 4 of Dengue virus. The results showed also that 30% of the E2 of Chikungunya virus positive samples reacted with E2 of O'nyong-nyong virus, and 5% with NS1 of Usutu virus and to the serotype 3 of Dengue virus. Among the 31 positive samples of O'nyong-nyong virus, 6 (19%) had simultaneous reactions with the E2 of Chikungunya virus. For the other antigens, simultaneous reactions with other antigens were in general <15%.
Proportion of Simultaneous Reactivity to Antigenically Related Virus and Less Antigenically Related Ones
Discussion
The objective of this study was to estimate the prevalence of antibodies to arboviruses of public health importance in a large number of bats in Central and West Africa using the same serological assay. We tested 2579 samples from at least 21 bat species, representing to our knowledge one of the most comprehensive report on arboviruses in bats in Africa. Our data showed spatial and temporal variation of IgG antibodies to diverse arboviruses. We observed an overall seroprevalence to arboviruses of 8.45%. This is lower than what was reported from (the few) previous seroprevalence studies in African bats, but different serological methods were used (i.e., agglutination vs. Luminex enzyme immuno assay [EIA]), the geographical location and the study period can also have an impact (Shepherd and Williams 1964).
Bats are flying mammals that nightly fly up to 20 km for foraging and up to hundreds or thousands of kilometers during seasonal migrations for some species (Roberts et al. 2012, Schloesing et al. 2020). Comparing prevalence in different collection sites is thus only meaningful if they are sufficiently distant. In this study, almost all our study sites are 20–300 km apart (Fig. 1). IgG antibodies to at least 1 arbovirus antigen were detected in 17 of the 21 sites investigated and, in the 4 sites with no positive samples, the sample size was low (i.e., <50). Furthermore, the positivity rate profile for antibodies to arboviruses was statistically different according to the ecological settings (Supplementary Table S2).
These differences could be related to bat species, environment, human factors, or a combination of those. For example, in Yaoundé and Obala where most bat samples studied were collected are urbanized areas, which could favor a vector population to interact between bats and humans in the same environment, thus contributing to sustain the transmission cycle of arboviruses.
Similar to other studies, we found variable prevalence by bat species, most likely corresponding to different ecology and exposure to arthropod vectors (Shepherd and Williams 1964, Kading et al. 2018, Hernández-Aguilar et al. 2021). We observed the highest prevalences in three frugivorous bat species; E. helvum, H. monstrosus, and R. aegyptiacus. E. helvum is widespread in equatorial Africa (Ruiz 2002), and is roosting in both urban and rural areas representing thus a good intermediate between sylvatic and urban cycles of arboviruses. Combined entomological and bat ecology studies are, therefore, urgently needed for a better understanding of the role of Chiroptera in the lifecycle of arboviruses in Africa.
Contrasting with the recurrent outbreaks of Chikungunya virus in DRC (Selhorst et al. 2020), we did not observe IgG antibodies to Chikungunya virus in bat samples from this country, which might be due to the overall limited number of bat samples tested in this large country. Nevertheless, antibodies to Chikungunya virus have been readily detected in free-ranging NHPs in DRC, but in different areas as where we tested bats in this study (Raulino et al. 2021). It is also possible that bats are not equally exposed to the same array of mosquito vectors than NHPs that are involved in the sylvatic cycles of arboviruses as suggested by others (Pastorino et al. 2004, Simo et al. 2019, Guzmán et al. 2020, Selhorst et al. 2020).
Usutu virus has been isolated from mosquitoes, birds, and some mammals in Europe and in some countries in Africa such as Senegal, Central African Republic, South Africa, and Uganda but data on Usutu virus in Africa are still very limited (Nikolay et al. 2011, Clé et al. 2019, Diagne et al. 2019). In this study, we observed antibodies against to Usutu virus, indicative of the circulation of this virus in at least three African fruit bat species (Table 3). To the best of our knowledge, this is the first evidence of Usutu virus circulation in bats from Africa, suggesting that they might contribute to the enzootic cycle of Usutu virus in Africa, but additional investigations to clarify the roles of bats in the viral cycle in Africa is needed.
The 12-month longitudinal follow-up of bat colonies in the same geographical location allowed to study impact of age, reproduction cycle, and eventual seasonality. We observed higher prevalence in adults than in immature or juvenile adults for H. monstrosus species, but not for E. helvum, suggesting a possible difference in behavior and ecology and subsequent exposure to vectors. During the female reproductive cycles, that is, gestation or lactation, no statistically significant difference was observed for both species.
Nevertheless, our study has some limitations, first of all the lack of confirmation of some of the seroreactivity by affinity-based virus neutralization assayer affinity measurement assays, which was hampered by the limited volumes of blood samples, that is, only a few drops of blood on filter paper. Another limitation is the absence of sequence confirmation of circulation of arboviruses that induced IgG antibodies detected during this study. Arbovirus RNA can only be detected during a limited period during acute infection; therefore, prospective studies are needed that should include blood samples in adequate storage conditions for molecular analysis.
Conclusion
Overall, the presence of antibodies suggests that bats could play a role in the sylvatic cycle of arboviruses in addition to NHPs and other mammals or animal species. Population size and the number of susceptible NHP at any time are most likely not high enough to maintain sylvatic arbovirus cycles and, therefore, other species such as bats with large numbers of offspring, that is, naive animals at any time, could thus also play also a role in sylvatic cycles. Moreover, several bat species live close to human settlements and create thus opportunities for contacts through an arthropod vector. More studies are still needed to document diversity of animal species susceptible to arboviruses and their role in sylvatic cycles and emergence of new viruses in humans.
Footnotes
Acknowledgments
We thank the staff and SIV team from Projet PRESICA (Innocent Ndong Bass, Aimé Mebenga, Joseph Moudindo, Thomas Atemkem and Flaubert Mba Djondzo) for the collection of samples and logistical support in Cameroon. In addition, we thank the field staff from DRC (Mubonga Mukulumanya, Lunguya-Metila Octavie, Guy Midingi and Mbenzo-Abokome Valentin).
Authors' Contributions
All authors participated in the study and approved the final version of the article. A.A. and M.P. conceived the study. S.-P.N.-K., I.N.B., R.D., S.G., and A.K.K. collected the samples. R.R., G.T., and A.E. performed the experiments. R.R., G.T., A.A., and M.P. analyzed the data. R.R., A.A., and M.P. drafted the article. A.A. and M.P. received funding. All the authors reviewed preliminary and final versions of the article.
Author Disclosure Statement
All the authors declared no conflicting financial interests.
Funding Information
R.R. received the PhD grant from INSERM and Montpellier University of Excellence (MUSE). A.K.K. was supported by fellowships from IRD and the University of Montpellier (MUSE, ANR-16-IDEX-0006). This study was supported by EBO-SURSY project funded by the European Union (FOOD/2016/379-660), by Research Fund Flanders (FWO, grant No. G054820N), and by a grant from MUSE (AbroSud project). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the article.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.