
Abstract
The objective of this study was to analyze the effect of the rural and urban microclimate on the presence of mosquitoes. Temperature (T) and relative humidity (RH; indoors and outdoors), as well as mosquito richness and abundance were measured in two sites (urban and rural) of the Yucatan State, Mexico. Species richness was higher in the urban site, whereas mosquito abundance was higher in the rural site. The microclimates of urban and rural housing differently affect mosquito richness and abundance. Mosquito richness and abundance were higher outdoors than indoors in the urban site, but they were higher indoors than outdoors in the rural site. For the urban site, analysis of the relation of T and RH with the registered parameters revealed that species richness increased with increasing indoor RH, and that mosquito abundance increased with increasing indoor T and RH. In the rural site, species richness was not affected, but abundance increased with increasing T and RH (indoors as well as outdoors). Results are discussed in the context of the management of mosquito transmitted diseases. No IRB approval was necessary since no ethical implications were identified to be reviewed by the ethical committee for the research of the ECOSUR Institution.
Introduction
The distribution and abundance of mosquitoes is influenced by a range of physical, chemical, and biological factors, including the type of landscape, degree of anthropization, and breeding site type and availability (Liu et al. 2012, Ferraguti et al. 2016, Chen et al. 2019). A key role is played by environmental conditions (Githeko et al. 2000, Delatte et al. 2009), particularly by factors such as temperature and relative humidity (RH) (López-Latorre and Neira 2016). Even subtle variations in these factors can modify the life cycle of mosquitoes, affecting their metabolism (Murdock et al. 2017), larval survival rate, and emergence time (Delatte et al. 2009, Couret and Benedict 2014, López-Latorre and Neira 2016). Each mosquito species is characterized by an optimal temperature and RH range that regulate its abundance and promote or limit its dispersal (Thomson 1938, Platt et al. 1957).
The ongoing process of urbanization has changed housing conditions, vegetation cover, and population density (Figuerola and Mazzeo 1998, Stone 2007). Urban sites with sparse vegetation cover can reach temperatures that are 5–11°C higher than the surrounding countryside (Aniello et al. 1995, Oke 2002, Chen et al. 2006, Mohajerani et al. 2017). Vegetation cover has an important cooling effect on the urban environment (Shiflett et al. 2017).
This heterogenicity shapes the local microclimate (Romero et al. 2010, Syed Othman Thani et al. 2017) and can affect mosquito communities. The type of construction, surface, and used building materials can influence the microclimate in and outside of buildings (Jatta et al. 2018) and hence determine the presence or absence of mosquitoes (Honório et al. 2009, Rodrigues et al. 2015). Although it is known that variations in temperature and RH affect mosquito abundance (Dow and Gerrish 1970, Githeko et al. 2000, Delatte et al. 2009, Costa et al. 2010, Yamana and Eltahir 2013), the impact of the rural and urban microclimate on mosquito populations has not been fully studied.
This study was conducted in two tropical localities with different levels of urbanization in Yucatan state, Mexico; the objective was to examine the effects of temperature and RH of the microclimate on the richness and abundance of mosquitoes indoors and outdoors.
Materials and Methods
Study area
The study was conducted in two locations in the northwest of Yucatan state, Mexico: the city of Mérida (MER; 20°58′03″ N, 89°37′27.81″ W, 10 meters above sea level [asl]), an urban area with 921,771 inhabitants, and the town of San Antonio Tedzidz (SAT; 20°50′01.77″ N, 89°58′33.83″ W, 8 meters [asl]), which is a rural locality with 1287 inhabitants (INEGI 2020). The two localities are 40.1 km apart. The climate of the study area can be described as tropical wet-and-dry, with an average annual temperature of 26°C and an annual precipitation of 1100 mm.
Vegetation and built (paved and construction areas) cover were estimated for both localities by using a geographic information system (GIS; ArcMap 10.1). An unsupervised classification procedure was applied to Landsat 8 satellite images (2019 U.S. Geological Survey, 30 meters/pixel resolution). The perimeter of the study area was determined from its radius (from the center to most distant point of the locality). In MER (166.7 km2), vegetation and built cover were 34.8% and 65.2%, respectively, compared with 51.5% and 48.5%, respectively, in SAT (0.94 km2).
In each locality, 21 residences were selected by snowball sampling (Vogt 1999, Atkinson and Flint 2001, Miller and Brewer 2003) in as random a fashion as possible. Each residence was georeferenced and defined as sampling unit (SU) in Fig. 1.
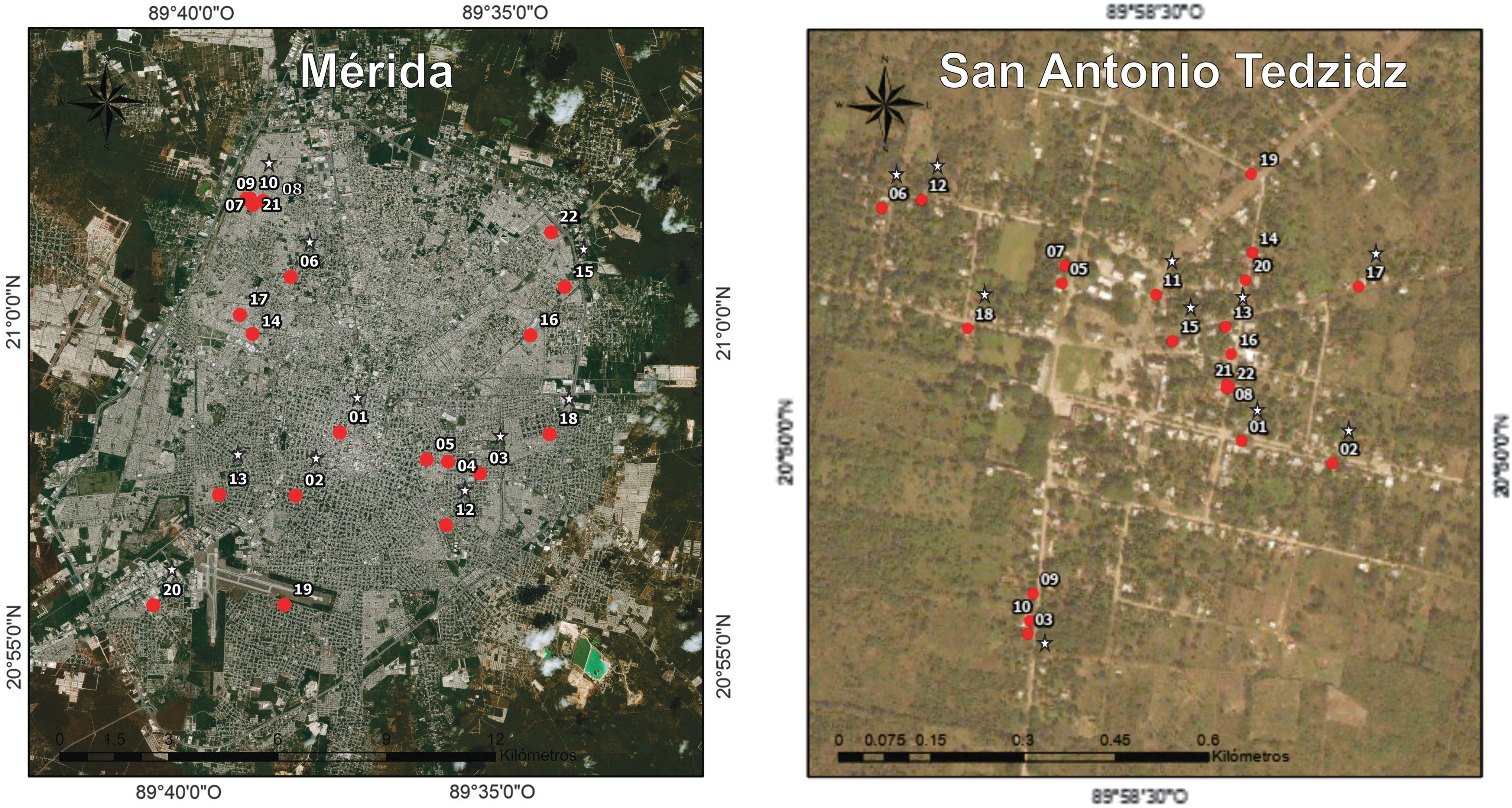
Study area, Mérida as the urban site with 21 SU, and San Antonio Tedzidz as the rural site with 21 SU. ✩: Subgroup of 10 SU per site for indoors and outdoor temperature and relative humidity record. SU, sampling unit.
Temperature and RH
A subgroup of 10 SU was selected in each locality. Their distribution in the locality was considered homogenous and included residences with different characteristics (e.g., construction material, with or without courtyard, courtyard with or without plants, presence or absence of swimming pool, water well, etc.). Thermohygrometers were installed in each SU (one indoors and one outdoors; datalogger STH-1; TOHTICS) and were programmed to register the temperature (T, as°C) and the RH (as %) every hour from September 2019 until March 2020. Thermohygrometers installed indoors were set up in the living or dining room, whereas those installed outdoors were placed in the shadow at 2 meters from the building. This setup allowed to obtain minimum, maximum, and average daily values (indoors as well as outdoors) of the temperature and RH for both localities.
Mosquito collection
Adult mosquitoes were collected from all SU by utilizing an aspirator (Improved Prokopack Aspirator, John W. Hock, model 1419) from 7:00 am to 1:00 pm (Silver 2007, Vazquez-Prokopec et al. 2009, Young et al. 2021). In each SU, mosquito collection lasted <10 min indoors (dark sites) and <10 min outdoors (vegetated sites and accumulated containers). Collections in each SU were performed once monthly from September 2019 to March 2020 (there was no mosquito collection in December) to represent the different climatic seasons of the region.
The collected mosquitoes were stored and transported under cold chain conditions (4°C) to the insectarium of the Laboratorio de Arbovirología of the Centro de Investigación Regional Dr. Hideyo Noguchi (Universidad Autónoma de Yucatán) in Mérida. Collected specimens were identified by using the taxonomic keys of Carpenter and LaCasse (1955).
Data analysis
Daily average indoor and outdoor T and RH were compared between sites by using mixed linear models (MLM). The models showed a normal distribution of residuals (Shapiro-test >0.05).
A generalized linear model (GLM; gamma-distributed with log link) was applied to correlate the richness and abundance of indoor-collected with outdoor-collected mosquitoes. The gamma distribution handles over-dispersed data that do not follow a normal distribution (McCullagh and Nelder 1989, Crawley 2012), which is the case for the variation in mosquito abundance between SU. Mosquito richness and abundance were considered as response variables, whereas the collection site (indoors or outdoors) was entered into the model as an independent variable.
The effect of the average temperature and RH on mosquito richness and abundance (in and outdoors) was analyzed independently for each locality and collection site by using GLM (Poisson distribution with log link, recommended for count data). The possible influence of the absence of collected mosquitoes in SU was calculated with zero-inflated GLM. This model integrates a traditional Poisson GLM (used for count data) with a binomial model and considers a possible excess of zero counts (Crawley 2012). These analyses allow to reliably assess the effect of positive (+) or negative (−) of environmental variables on mosquito richness and abundance and indicate whether the richness and abundance will increase or decrease if the variables increase.
All statistical analyses were performed with the R project software package (R Development Core TEAM 2012). The pscl package (developed at Stanford University) (Jackman et al. 2015) was used for zero-inflated GLM.
Results
Temperature and RH
Due to the malfunctioning of several sensors, it was not possible to obtain all of the planed data. In total, 43,459 T and RH data points were recorded in SAT (23,062 indoors and 20,397 outdoors) and 38,015 data points were recorded in MER (19,367 indoors and 18,648 outdoors).
Temperature values registered in MER showed less variation than those recorded in SAT. In both localities, minimum as well as maximum T were registered outdoors. Indoor and outdoor average T were similar in MER (χ2 = 1.57, p > 0.05) but significantly different in SAT (χ2 = 16.693, p < 0.0001). In SAT, the average indoor T was 2.3°C higher than the average outdoor T (Table 1).
Minimum, Maximum, and Average Temperature, and Relative Humidity Registered During the Mosquito Sampling Period in Two Localities with Different Urbanization
RH, relative humidity; T, temperature.
The RH data generally followed the same tendency as T and showed less variation in MER than in SAT. In both localities, the largest variation between minimum and maximum RH was registered outdoors. Also, average RH was significantly higher outdoors than indoors in both sites (Table 1; MER: χ2 = 4.38, p < 0.05; SAT: χ2 = 5.50, p < 0.05).
Mosquito collection
In total, 2665 mosquitoes distributed in two genera and six species were collected: Aedes aegypti (Linnaeus), Aedes taeniorhynchus (Wiedemann), Culex quinquefasciatus Say, Culex tarsalis Coquillett, Culex nigripalpus Theobald and Culex coronator Dyar y Knab. Twenty-one-point-six percent of the mosquitoes was collected in MER (226 indoors and 352 outdoors) and 78.4% in SAT (1543 indoors and 544 outdoors). Ae. aegypti and Cx. quinquefasciatus were the most abundant species; although Ae. aegypti was the more abundant species in MER (indoors as well as outdoors), Cx. quinquefasciatus was the more abundant species in SAT (indoors as well as outdoors). Cx. tarsalis and Cx. coronator—were only encountered in peripheral SU in MER (Cx. tarsalis: SU 3 [2 ind.] and SU 15 [1 ind.]; Cx. coronator: SU 8 [1 ind.] and SU 22 [1 ind.]) (Fig. 1 and Table 2).
Abundance of Mosquito Species (Indoors and Outdoors, n = 21) in Two Localities with Different Urbanization
The GLM analysis revealed that mosquito abundance and species richness were significantly higher outdoors in MER. In SAT, mosquito abundance and species richness were significantly higher indoors (Fig. 2).
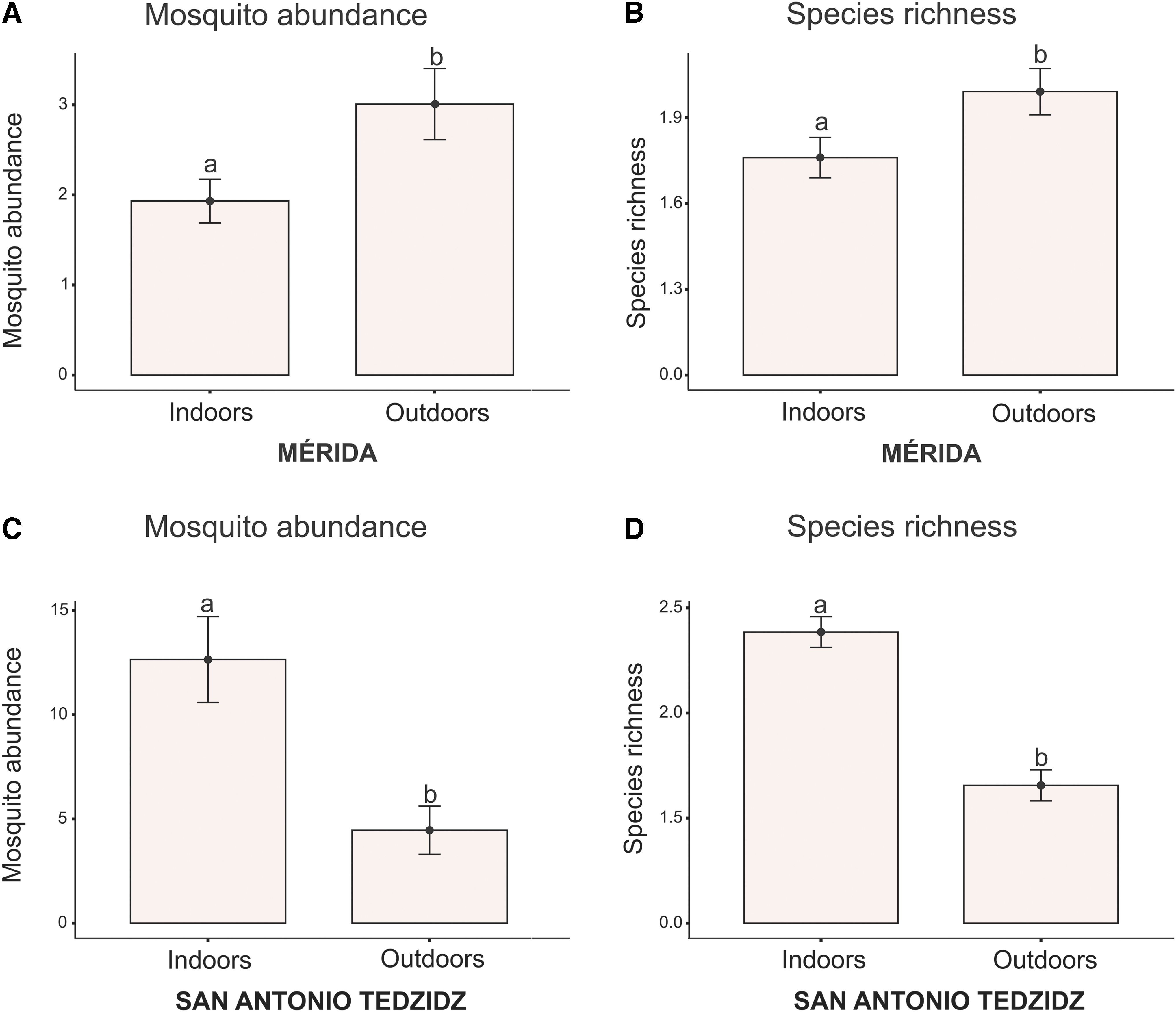
Generalized linear model (gamma-distributed with log link) of species richness and mosquito abundance in both localities: Mosquito abundance
Effect of T and RH on mosquito abundance and richness
In the case of mosquito abundance, the T and RH had an impact indoors as well as outdoors in SAT, whereas in MER the impact could be only observed indoors (Table 3). In the case of mosquito richness, only the indoor RH had an impact in MER, whereas no impact was observed in SAT (Table 4).
Analysis of Indoor and Outdoor Mosquito Abundance in Mérida and San Antonio Tedzidz Compared with the Average Temperatures and Relative Humidity (Generalized Linear Model)
Bold values of p represent statistically significant effect of the parameter.
A negative (−) value for the estimate ± SE indicates a negative effect of the variable on mosquito abundance.
SE, standard error.
Analysis of Indoor and Outdoor Species Richness in Mérida and San Antonio Tedzidz Compared with the Average Temperatures and Relative Humidity (Generalized Linear Model)
Bold values of p represent statistically significant effect of the parameter.
A negative (−) value for the estimate ± SE indicates a negative effect of the variable on mosquito species richness.
Discussion
The present study demonstrates that microclimatic conditions evaluated (indoors or outdoors, rural or urban) influence the richness and abundance of mosquitoes. Mosquito abundance was lower in MER than in SAT. Urban zones can be warmer and less humid than rural areas, which means that mosquito larval survival, body size, and growth rate can be reduced in urban areas (Murdock et al. 2017). In MER, 60% of the mosquitoes were collected outdoors; on the other hand, in SAT, 73% of the individuals were collected indoors.
Moreover, it was in these conditions where similar values for T (14–15°C) and RH (24%) between both sites were found, that there seem to be optimal microclimatic values for the survival of adults mosquitoes (Thomson 1938, Platt et al. 1957, Delatte et al. 2009). The abundance of Ae. aegypti females was found to be associated with the minimum temperature in São Paulo, Brazil (Rodrigues et al. 2015) (between 12°C and 24°C following Fig. 2C); however, in Rio de Janeiro, Brazil, the infestation with Ae. aegypti and transmission of dengue were related to an increase in minimum temperatures (Câmara et al. 2009).
Species richness was higher in MER than in SAT. Landscape anthropization has been shown to reshape the local habitat and create new opportunities for the invasion of anthropophilic mosquitoes (Ribeiro et al. 2012, Mayi et al. 2020). The urban landscape accommodates a diverse mosaic of land-use types. In this urban patchwork, vegetation can influence temperature variations (Misslin et al. 2016) and promote the establishment of mosquito species with different ecological requirements.
Ae. aegypti and Cx. quinquefasciatus, the most abundant mosquito species in both localities, are well-known transmission vectors for clinically important arboviruses. Both species are widely distributed in the study area (Baak-Baak et al. 2017). The highly anthropophilic Ae. aegypti, which is known to predominate in urban environments (Nelson 1986, Wilke et al. 2019), was the more abundant species in MER. This species, however, was more than twice as abundant in rural SAT. Nevertheless, Cx. quinquefasciatus was the most abundant species in SAT and was 6.6 times more abundant than in MER. Cx. quinquefasciatus oviposits in many different water sources and can attain a high larval density in water with high organic content. The species is adapted to the muddy waters of sewage treatment plants, drains, and pit latrines (Calhoun et al. 2007).
In addition, Ae. taeniorhynchus and Cx. nigripalpus were encountered in both localities. Ae. taeniorhynchus is a vector of the Venezuelan equine encephalitis virus. Larvae grow in coastal marshes, freshwater pools, brackish water swamps, and intermittent water bodies (Nielsen and Nielsen 1953, Carpenter and LaCasse 1955). Cx. nigripalpus, on the other hand, can transmit the Saint Louis encephalitis and the West Nile virus (Dow and Gerrish 1970, Godsey et al. 2013). This mosquito breeds in perennial and intermittent clear water bodies, such as marshes, canals, and ponds.
Cx. tarsalis and Cx. coronator were only encountered in MER and have previously been reported there by Baak-Baak et al. (2017). In Cameroon, mosquitoes of the genus Culex have been documented to prefer habitats with less vegetation (Mayi et al. 2020). Cx. tarsalis transmits the West Nile (Turell et al. 2002) and Cache Valley virus (Ayers et al. 2018), whereas Cx. coronator vectors the West Nile (Alto et al. 2014) and the Saint Louis encephalitis virus (Turell et al. 2005). These species can be encountered in ditches, temporary ponds, springs, and natural gas easement (Goddard et al. 2006).
The present report describes the influence of T and RH on mosquito populations in different environments and opens the door to other questions: What is the effect of minimum T on mosquito abundance? How do certain aspects—social factors, demographics (Rodrigues et al. 2015), domestic water storage, connection to a central water supply system, waste collection (Causa et al. 2020), and house design (Jatta et al. 2018)—shape the dynamics of mosquito populations in areas with different levels of urbanization? Since many mosquitoes can transmit diseases, an integrated understanding of these aspects will allow the development of improved mosquito control and prevention strategies.
The results from the present investigation agree with other studies (Oke 2002, Shiflett et al. 2017) that show how vegetation cover can substantially reduce the temperature and increase the environmental humidity. Our results indicate that the average T was higher and the RH lower indoors than outdoors. The indoor average and maximum T were higher in SAT than in MER. This can perhaps be explained by the heat capacity differences of surface (Ando et al. 2009) and building materials used for the construction of the SU.
These heat capacity differences could affect the indoor microclimate of the residence (EPA 2008, Mohajerani et al. 2017, Jatta et al. 2018). Several publications recognize that the temperatures in urban zones are higher than in rural areas because of the presence of urban elements. However, temperatures recorded in MER (with 65.2% urban elements) were lower than in SAT. Perhaps this observation can be explained by local variations in urban vegetation, which is dispersed and heterogeneous. The resulting mosaic of spaces can become a patchwork of warm and cold clusters that cause local temperature variations (Ando et al. 2009, Misslin et al. 2016).
Conclusion
Our results present useful information for mosquito management. Implementation of this information in the field could be used to improve microclimatic conditions indoors and outdoors to mitigate mosquito-related issues.
Footnotes
Acknowledgments
The authors would like to thank Christopher Albor Pinto for his assistance with statistical analyses, and César Tzuc Dzul for his support with the taxonomical identification of mosquitoes. They are grateful to Alma Rosa Dzul Rosado and Mildred Patricia López Uribe for their invaluable help during the field work, and to the participating families of Mérida and San Antonio Tedzidz for allowing entrance to their properties. They also wish to thank Dr. Carlos Machain Williams for providing access to the insectarium of the Laboratorio de Arbovirología (Universidad Autónoma de Yucatán) and storage of mosquitoes. They are thankful to Dieter Waumans for the article translation. S.S.F.R. would like to acknowledge support from a master's scholarship (952430) awarded by Consejo Nacional de Ciencia y Tecnología (CONACyT).
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by a student fellowship awarded by CONACYT (No. 952430).