
Abstract
Background:
The emergence of insecticide resistance in Aedes vectors mosquitoes poses a real challenge for arboviral-borne disease control. In Côte d'Ivoire, data are available on phenotypic resistance and the presence of kdr mutations in Aedes aegypti. Therefore, information on metabolic resistance in Aedes populations is very scarce. Here, we assessed the insecticide resistance status of Ae. aegypti in periurban and rural areas of Côte d'Ivoire, and we investigated the role of detoxification enzymes as possible resistance mechanisms.
Materials and Methods:
Aedes mosquito eggs were collected between June 2019 to April 2021 in two agricultural sites. Adults of Ae. aegypti were tested using World Health Organization tube assays, with seven insecticides belonging to pyrethroids, organochlorines, carbamates, and organophosphates classes. We determined the knockdown times (KdT50, KdT95) and resistance ratios of pyrethroids in natural populations. The synergist piperonyl butoxide (PBO) was used to investigate the role of enzymes in resistance. Biochemical assays were performed to detect potential increased activities in mixed-function oxidase levels, nonspecific esterases (NSEs), and glutathione S-transferases.
Results:
The results showed that Ae. aegypti populations were resistant to five insecticides with mortality of 46% and 89% for 0.75% permethrin, 68% and 92% for 0.05% deltamethrin, 57% and 89% for lambda-cyhalothrin, 41% and 47% for dichlorodiphenyltrichloroethane (DDT), 82% and 91% for chlorpyrifos-methyl in Songon-Agban and Kaforo, respectively. Susceptibility to carbamates was observed in our study sites. After exposure to PBO, the susceptibility of Ae. aegypti to pyrethroids and DDT was partially restored in Songon-Agban. Whereas in Kaforo, none increase of the mortality rates of Ae. aegypti for these four insecticides was observed after exposure to PBO. Increased activity of NSE (α-esterases) was found in Songon-Agban compared with the reference susceptible strain.
Conclusion:
These findings provide valuable information to support decisions for vector control strategies in Cote d'Ivoire. Also, we highlight the need for the monitoring of insecticide resistance management in Aedes vectors.
Introduction
Dengue, chikungunya, yellow fever, and Zika fever represent four important arboviral diseases worldwide (Weetman et al., 2018). These diseases are real threats to public health in numerous tropical and subtropical countries (Crawford et al., 2017). Among them, dengue is the most widespread and fastest spreading worldwide. People experiencing dengue virus infections are estimated at ∼390 million per year, suggesting 96 million cases of apparent manifestation (Bhatt et al., 2013). The disease is now endemic in >100 countries in the World Health Organization (WHO) regions of Africa, America, Eastern Mediterranean, South east Asia, and Western Pacific (World Health Organization, 2022).
In Côte d'Ivoire, dengue epidemics occurred in several locations, but the city of Abidjan was the most affected over the last two decades (Akoua-koffi et al., 2011; MHPHCI, 2019; World Health Organization, 2017). Aedes aegypti is considered to be the most important Aedes species responsible for arboviruses transmission (World Health Organization, 2010). This species is highly anthropophilic and breeds mostly in artificial containers.
Improper disposal of waste containers provides a perfect breeding environment for Ae. aegypti mosquitoes (Banerjee et al., 2015). In Côte d'Ivoire, yellow fever and dengue viruses are transmitted by Ae. aegypti (Guindo-Coulibaly et al., 2019; Guindo-Coulibaly et al., 2010; Kone et al., 2013; Zahouli et al., 2016). This vector is widely distributed in urban areas, but its ecology and insecticide susceptibility remain poorly understood in periurban and rural areas.
Because of the current lack of treatments or vaccines for most arboviral diseases, the prevention mainly relies on vector controls largely based on insecticide treatments (World Health Organization, 2009). Pyrethroids and organophosphates are the main chemical groups used and approved for public health.
Synthetic pyrethroids: deltamethrin, cypermethrin, lambda-cyhalothrin, and permethrin, are broadly used worldwide because of their effectiveness and rapid activity against insects, their low toxicity for mammals, and their rapid decay in the environment (Amelia-Yap et al., 2018; World Health Organization, 2009). The long-term intensive use of insecticides leads to the emergence of resistance phenomenon in the populations under selection pressure, which is one of the main obstacles to the arthropod pest control (Labbé et al., 2017).
Many control programs are under threat of insecticide resistance in Ae. aegypti reported almost worldwide (Dusfour et al., 2019; Kushwah et al., 2020; Marcombe et al., 2011; Stenhouse et al., 2013). The insect resistance to pyrethroids and dichlorodiphenyltrichloroethane (DDT) can be due to mutations in the sodium voltage-gated channel (VGSC) gene, which are known as “knockdown resistance” (kdr) (Labbé et al., 2017). Mutations in the VGSC genes are reported in several mosquito species, including Ae aegypti. Three kdr mutations (V410L, V1016I, and F1534C) have been identified in Ae. aegypti from West Africa (Badolo et al., 2019; Kawada et al., 2016; Konan et al., 2021).
Resistance may also be associated with the overexpression or qualitative changes in the enzyme involved in xenobiotic detoxification. Previous studies have shown that such mechanisms of metabolic resistance could be also present in African populations of Ae. aegypti (Namountougou et al., 2020; Ngoagouni et al., 2016).
In Côte d'Ivoire, most of the data on insecticide susceptibility concern malaria vectors (Ahoua et al., 2012; Fodjo et al., 2018; Zoh et al., 2018). Aedes species remains poorly documented in this field, especially on the potential implication of detoxification enzymes as resistance mechanisms. The few data available on the resistance of Ae. aegypti populations were reported from the city of Abidjan and Bouaké. These data showed the resistance of Ae. aegypti to chlorpyrifos-methyl as well as pyrethroids and DDT, associated with three kdr mutations V410L, V1016I, and F1534C (Guindo-Coulibaly et al., 2014; Konan et al., 2021; Kouadio et al., 2022).
In periurban and rural areas, the regular intensive use of insecticides in agricultural practices could also affect the susceptibility of vectors to insecticides (Nkya et al., 2014). The implication of insecticide use in agricultural practice in the emergence of resistance of Anopheles gambiae s.l. has been reported in the rice- and vegetable-growing areas (Ahoua et al., 2012; Camara et al., 2018; Zoh et al., 2018) and cotton-growing areas in Côte d'Ivoire (Koffi et al., 2013).
Thus, Ae. Aegypti, which are abundant in domestic and peridomestic environments, could be also exposed to selection pressure induced by agricultural treatments. It is important to deepen knowledge on this vector in different agricultural areas. This study aimed to assess the insecticide resistance status of Ae. aegypti in periurban and rural areas of Côte d'Ivoire, and to investigate the role of detoxification enzymes in the resistance mechanisms.
Materials and Methods
Study areas
The study was carried out in Songon-Agban (periurban site) in the southern and Kaforo (rural area) in northern Côte d'Ivoire (Fig. 1). The GIS software version QGIS 2.14 software was used to generate 20 random points inside each site, devised to 10 points in dwellings and 10 points in plantations. The average distances between ovitraps in the dwelling and the plantations are 100 meters and 500 meters, respectively. A previous study in Côte d'Ivoire showed that Ae. aegypti can fly up to 800 meters from the point of release (Guindo-Coulibaly et al., 2019).
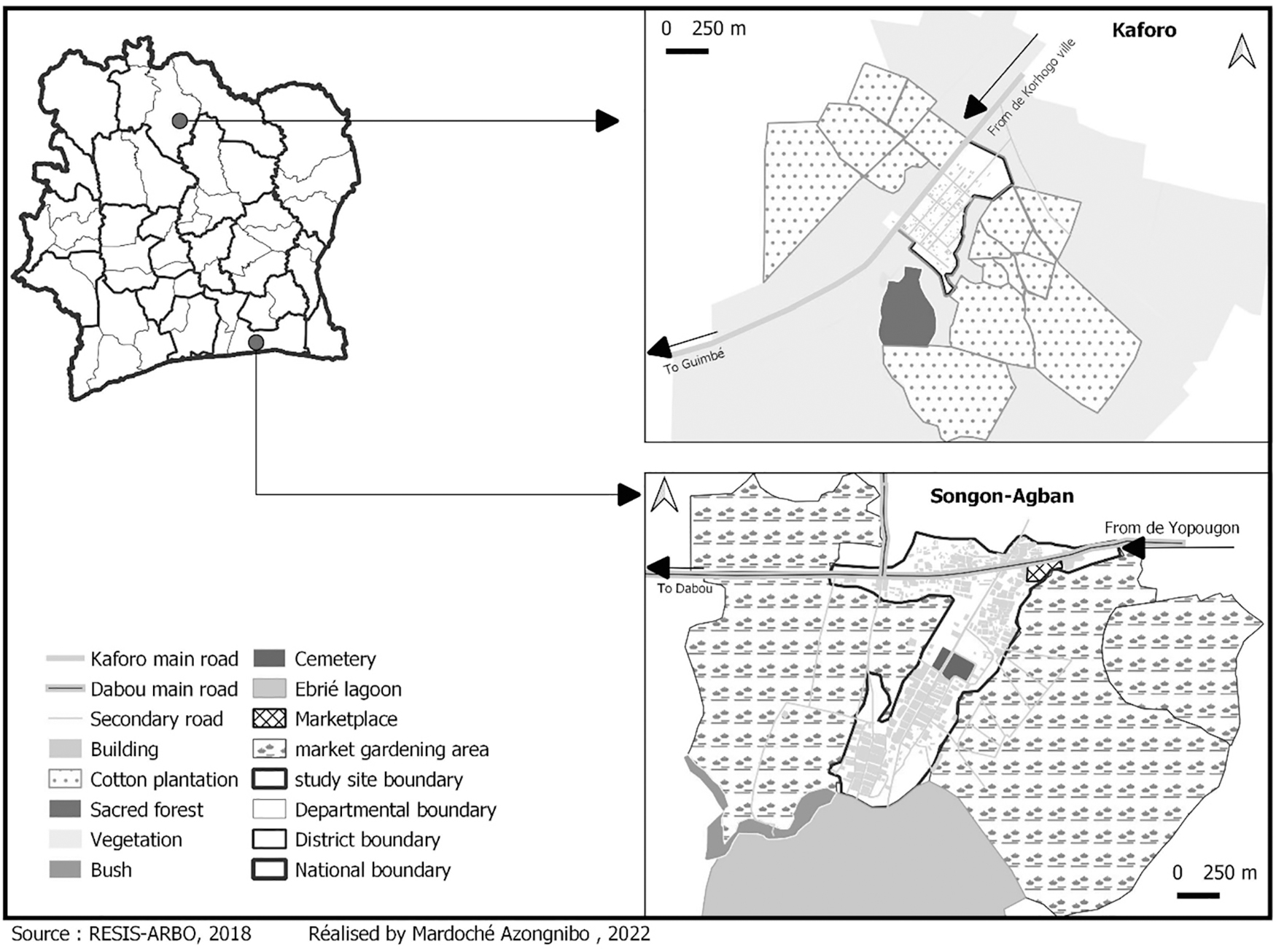
Aedes aegypti collection sites in Songon-Agban (southern) and Kaforo (northern), Côte d'Ivoire.
Songon-Agban is located to the east of the autonomous district of Abidjan. This site is characterized by a subequatorial climate, with four seasons. There are two rainy seasons: a long rainy season (March–July) and a short rainy season (September–November), and two dry seasons: the short dry season (August) and the long dry season (December–February). Songon-Agban is made up of three neighborhoods: Agban-village, Agban station, and the new neighborhood; bordered by the Ebrié lagoon and a large area of shallows where market gardening is cultivated, making Songon-Agban a commercial site.
Kaforo is located in the subprefecture of Tiorognadougou, 15 km from the Department of Korhogo. This village is characterized by a tropical climate, with one long rainy season (March–November) and a short dry season (December–February). The cultivated parcels of cotton surround the village.
Criteria for selection of study sites
Songon-Agban and Kaforo have been chosen because of their historical intensive use of insecticides. Since 1960, when Côte d'Ivoire became independent, agriculture has been widely developed in periurban and rural areas. This activity sometimes requires intensive use of pesticides to control pests and to obtain a better yield. Vegetable and cotton crops are among the agrosystems that require intensive use of pesticides. Songon-Agban and Kaforo sites are well known as the leading areas for vegetable and cotton crops, respectively. Songon-Agban belonged to the “Société de Culture de Banane” company during the 1980s.
Since the 2000s, the lowlands have been developed by local people for market gardening. Kaforo belonged to the cotton area managed by the “Compagnie Ivoirienne de Coton” company since 1980s. We chose these leading sites to have a great probability of observing the highest levels of resistance because of the high insecticide pressure on the insect populations. This pressure could accelerate the development of resistance in mosquitoes, especially Ae. aegypti, and the appearance of several mutations in the genes involved in this résistance. Previous study has reported the emergence of insecticides resistance of An. gambiae s.l. in the vegetable- and rice-growing areas (Ahoua et al., 2012; Camara et al., 2018; Zoh et al., 2018) and cotton-growing areas (Koffi et al., 2013) in Côte d'Ivoire.
Mosquito collection
The sampling was performed in one periurban site (Songon-Agban) and one rural site (Kaforo). Aedes mosquito eggs were collected using the standard WHO ovitraps. Six surveys were carried out at each site from June 2019 to April 2021. The sampling carried out in the dwellings and the plantations for each site. The rearing of mosquitoes was carried out at the insectarium of the Medical and Veterinary Entomological Centre in Abidjan under standard conditions of temperature 25°C (± 2°C) and humidity 75% (± 10%).
After emergence, adult mosquitoes were identified under binocular magnifier using the mosquito morphological identification key of Edward (1941) and the African Aedes species morphological identification key of Huang (2004). The F1 generation (first generation issued from adults obtained with field eggs) of Ae. aegypti adult mosquitoes was used for bioassays.
Insecticides
Seven insecticides were tested: three pyrethroids (0.05% deltamethrin, 0.05% lambda-cyhalothrin, and 0.75% permethrin), two carbamates (0.1% propoxur and 0.1% bendiocarb), one organophosphate (0.4% chlorpyrifos-methyl), and one organochlorine (4% DDT). Insecticide-impregnated papers were prepared by MIVEGEC laboratory according to WHO protocols (World Health Organization, 1998a).
The doses of insecticides used in our study are much higher than those recommended by the WHO for assessing the insecticide sensitivity of Aedes. However, several previous studies carried out in Côte d'Ivoire (Guindo-Coulibaly et al., 2014; Konan et al., 2021; Kouadio et al., 2022) and other African countries (Abdulai et al., 2023; Badolo et al., 2019; Djiappi-Tchamen et al., 2021) have demonstrated the resistance of Ae. aegypti to the same doses of insecticides used in our study. Moreover, in Côte d'Ivoire, vector control tools against Culicidae take into account these high insecticide doses. Therefore, it seemed wiser to consider these doses already used to know the real level of resistance of our Ae. aegypti populations. This could help the authorities to review the choice of insecticide doses used on mosquito populations for vector control in our country.
Bioassays
The bioassays were undertaken with WHO test kits using 2- to 5-day–old nonblood-fed F1 Ae. aegypti females (World Health Organization, 2016). SBE (susceptible reference strain from Benin) was reared in the same conditions that fields strain. Four exposure tubes were used for each insecticide. Twenty to 25 mosquitoes per tube were exposed for 1 h to the insecticide-impregnated papers. For every test, two control tubes of 20–25 females were run in parallel with untreated papers.
Papers impregnated with Risella oil, olive oil, and silicone oil served as controls for DDT, organophosphates/carbamates, and pyrethroids, respectively. The cumulative knockdown rate was recorded for pyrethroid insecticides at 5, 10, 15, 20, 25, 30, 40, 50, and 60 min. After the exposure time, mosquitoes were transferred into holding tubes, provided with 10% sugar solution, and kept at 25°C (± 2°C) and 70% (±10%) relative humidity. Mortality rates were recorded at 24 h postexposure.
Synergist assays with piperonyl butoxide
To assess the involvement of detoxifying enzymes in the resistance of Ae. aegypti to pyrethroids and DDT, females of 2- to 5-day–old nonblood-fed F1 Ae. aegypti were tested using as a synergist. Piperonyl butoxide (PBO)-impregnated papers were prepared by MIVEGEC laboratory according to WHO protocols (World Health Organization, 1998a). Two tubes of 20–25 females were pre-exposed to 4% PBO-impregnated papers for 1 h and subsequently exposed immediately to insecticide-impregnated papers for 1 h. The controls were exposed to PBO for 1 h and subsequently paper without insecticides for 1 h. Mortality rates of females exposed to PBO + insecticides were compared with females exposed to insecticides only.
Biochemical assays
Biochemical tests were carried out by using the WHO procedures (World Health Organization, 1998b). The assays were conducted on 2–4 days old Ae. aegypti adult mosquitoes from field collections (F0) and the reference strain SBE, unexposed to insecticides and stored at −80°C. For each experiment, ∼60 mosquitoes per species were assayed for mixed-function oxidase (MFO), glutathione S-transferases (GST), α- and β-esterases, and total proteins. Adult mosquitoes were individually homogenized in 200 μL of ice-cold distilled water.
The homogenate was centrifuged at 14,000 × rpm for 2 min at 4°C. Two microplate wells were then each filled with 20 μL of the centrifuged homogenate for the oxidases. The other four microplates were filled with 10 μL in two replicates for the dosage of proteins, and the evaluation of GST and esterase activities. The optical densities (ODs) were read using a spectrophotometer (Multiskan GO 1510-04280) Skanlt Software 6.0.1.
Total proteins
Total protein was measured for each mosquito using the method of Bradford (1976). Ten microliters of each homogenate were mixed with 200 μL of a solution of “Coomassie Plus Protein Assay Reagent” half diluted in distilled water (1 volume of protein reagent per 1 volume of water). The plate was held for 5 min at room temperature, and the OD was read at end-point at 590 nm. Standard curve was established with bovine serum albumin.
MFO assay
The assay used to measure MFOs detects increase in the amount of hemes, which are then converted into equivalent units of cytochrome P450. Cytochrome P450 was titrated using the heme peroxidase assay according to Brogdon et al. (1997). In brief, 80 μL of 0.625 M potassium phosphate buffer (pH 7.2) was added to 20 μL of microfuged supernatant and 200 μL of tetramethylbenzidine solution (0.012 g 3,3′,5,5′-tetramethylbenzidine +6 mL methanol +18 mL sodium acetate buffer 0.25 M, pH 5.0). Then, 25 μL of 3% hydrogen peroxide was added to each well, and the plate was incubated for 30 min at room temperature with a cover. The end-point absorbance was read at 630 nm, and values were calculated from a standard curve using cytochrome C.
Glutathione S-transferase assay
GST assay activity was performed according to the WHO guidelines (World Health Organization, 1998b). Ten microliters of each homogenate was mixed with 200 μL of reduced glutathione (GSH)/1-chloro-2,4 dinitrobenzene (CDNB) working solution (0.060 g of reduced GSH in 20 mL of sodium phosphate buffer 0.1 M pH 6.5 + 0.013 g of CDNB diluted in 1 mL of methanol) in a microplate well. The reaction was read at 340 nm immediately as a kinetic assay for 5 min. An extinction coefficient of 5.76 mM−1 (corrected for a path length of 0.6 cm) was used to convert absorbance values to moles of product. GST specific activity was reported as CDNB conjugated μmol product/min/mg protein.
α- and β-esterases
Nonspecific esterase (NSE) activity was assessed with two substrates, α- and β-naphthol acetate using the WHO protocol (World Health Organization, 1998b). For both substrates, 90 μL of 1% saline phosphate buffer (PBS) pH 6.5 was added to each well containing 10 μL of homogenate. After 10 min incubation at room temperature, 100 μL of (α or β-naphthyl acetate solution [600 μL of 0.06 M α-naphthyl acetate +3 mL of 1% PBS buffer (pH 6.5) + 8.4 mL of distilled water]) was added.
After incubation for 30 min at room temperature, 100 μL of solution (0.012 g Fast Garnett Salt +12 mL distilled water) was added to stop the reaction. The reaction mixture was then incubated for 10 min at room temperature. The concentration of the final product was determined at 550 nm, calculated from a standard curve using α- or β-naphthol. The specific esterase activity per individual was reported in μmol product/min/mg protein.
Data analysis
The resistance/susceptibility status of the tested mosquitoes was determined according to WHO criteria (World Health Organization, 2016), with 98–100% mortality at 24 h indicating susceptibility, mortality between 90% and 97%, suggesting possible resistance and <90% mortality confirming resistance. Mortality was corrected using Abbott's formula (Abbott, 1925) when the mortality of the control was between 1% and 5%. The tests were discarded when the control mortality was >5%.
The cumulative knockdown data were subjected to probit analysis (Finney, 1970) to estimate time at which 50% and 95% of the adult population would be killed (KdT50 and KdT95) using the time–effect option. These statistical analyses were performed using PoloPlus 1.0 Software (LeOra Software) (2002–2022).
Resistance ratios (RRs) for knockdown times (KdT50 and KdT95) were calculated by dividing KdT values for the field strains with those of reference strain.
GraphPad Prism version 5.0.1 software was used to analyze the susceptibility and biochemical assay data. Fisher's exact test was used to compare mortality rates before and after pre-exposure to PBO. The Mann–Whitney nonparametric U-test was used for comparison of enzymatic activity levels. The levels of MFO, NSE, and GST of Ae. aegypti from each of two sites were compared with SBE, and between the sites. Statistical significance was assumed at p < 0.05 and confidence intervals were 95%.
Results
The KdT50 and KdT95 of the reference susceptible strain (SBE) were 9 and 23 min for deltamethrin, 9 and 28 min for lambda-cyhalothrin, 10 and 24 min for permethrin, respectively. The laboratory SBE strain of Ae. aegypti confirmed its susceptibility to pyrethroids with 100% mortality. The KdT for Songon-Agban population was all significantly higher than that for SBE with RRs ranging from 1.9 to 9.4 for KdT50 and 8.9 to 311.1 for KdT95. The KdT for Kaforo population was also significantly higher than that for SBE but was lower than that for Songon-Agban ranging between 1.4 and 2 for KdT50 and 1.5 and 2.7 for KdT95 (Table 1).
The 50% Knockdown Time (KdT50), 95% Knockdown Time (KdT95), 95% Confidence Intervals, and Resistance Ratios of Aedes aegypti Field Populations from Songon-Agban and Kaforo Exposed to Pyrethroid Insecticides During a 1-h Exposure
95% CI, 95% confidence interval; KdT50 and KdT95, time at which 50% and 95% of the population are knocked down; RR, resistant ratio between field samples and susceptible laboratory strain; SBE, susceptible reference strain from Benin.
Insecticides susceptibility of Ae. aegypti
The results of WHO tests on Ae. aegypti populations from our study sites are presented in Table 2. For unexposed controls, all mortality rates between 1% and 5% were corrected with Abbott's formula (1925). Among the Ae. aegypti populations from our two study sites, the mortality rates ranged between 41% and 92%. For pyrethroid insecticides, the mortality rates ranged between 46% and 89% for permethrin, 68 and 92% for deltamethrin, and between 57% and 89% for lambda-cyhalothrin.
Mortality Rates After 24 h Holding Period for the Field Populations of Aedes aegypti Using WHO Test Tubes (1 h Exposure)
DDT, dichlorodiphenyltrichloroethane; n, number of tested mosquitoes; % M, mortality rate; St, resistance status; S, susceptible; R, resistant; SR, suspected resistance; p, probability (Fisher's exact test) of comparing the mortality rate between two study sites.
.
The lowest mortality rates were observed from Songon-Agban for all the three insecticides (Fisher's exact test: p ˂ 0.0001). With other insecticides, mortality rates were 41% in Songon-Agban and 47% in Kaforo for DDT, 82% in Songon-Agban and 91% in Kaforo for chlorpyrifos-methyl. However, with the two carbamates (propoxur and bendiocarb) susceptibility of Ae. aegypti was observed at both sites, with a mortality rate of 100% in Songon-Agban and 98% in Kaforo (Table 2).
Effects of PBO synergist on Ae. aegypti
After 24 h of observation, no mortality has been observed in all the control tubes, indicating that the synergist PBO had no toxic effect. Pre-exposure to PBO led to a partial restitution of the susceptibility of Ae. aegypti to pyrethroids (Fig. 2) and DDT (Fig. 3) in Songon-Agban. Mortality rates significantly increased from 46% to 75% for permethrin (Fisher's exact test: p = 0.002; odds ratio [OR] = 0.307; confidence interval [CI] = 0.136–0.696); 68% to 87% for deltamethrin (p = 0.012; OR = 0.303; CI = 0.108–0.847); 57% to 82% for lambda-cyhalothrin (p = 0.001; OR = 0.291; CI = 0.127–0.662); and 41% to 75% for DDT (p ˂ 0.0001; OR = 0.221; CI = 0.101–0.485); whereas in Kaforo, pre-exposure to PBO did not change the mortality rates of Ae. aegypti for these four insecticides (permethrin, deltamethrin, lambda-cyhalothrin, and DDT), p ˃ 0.05.
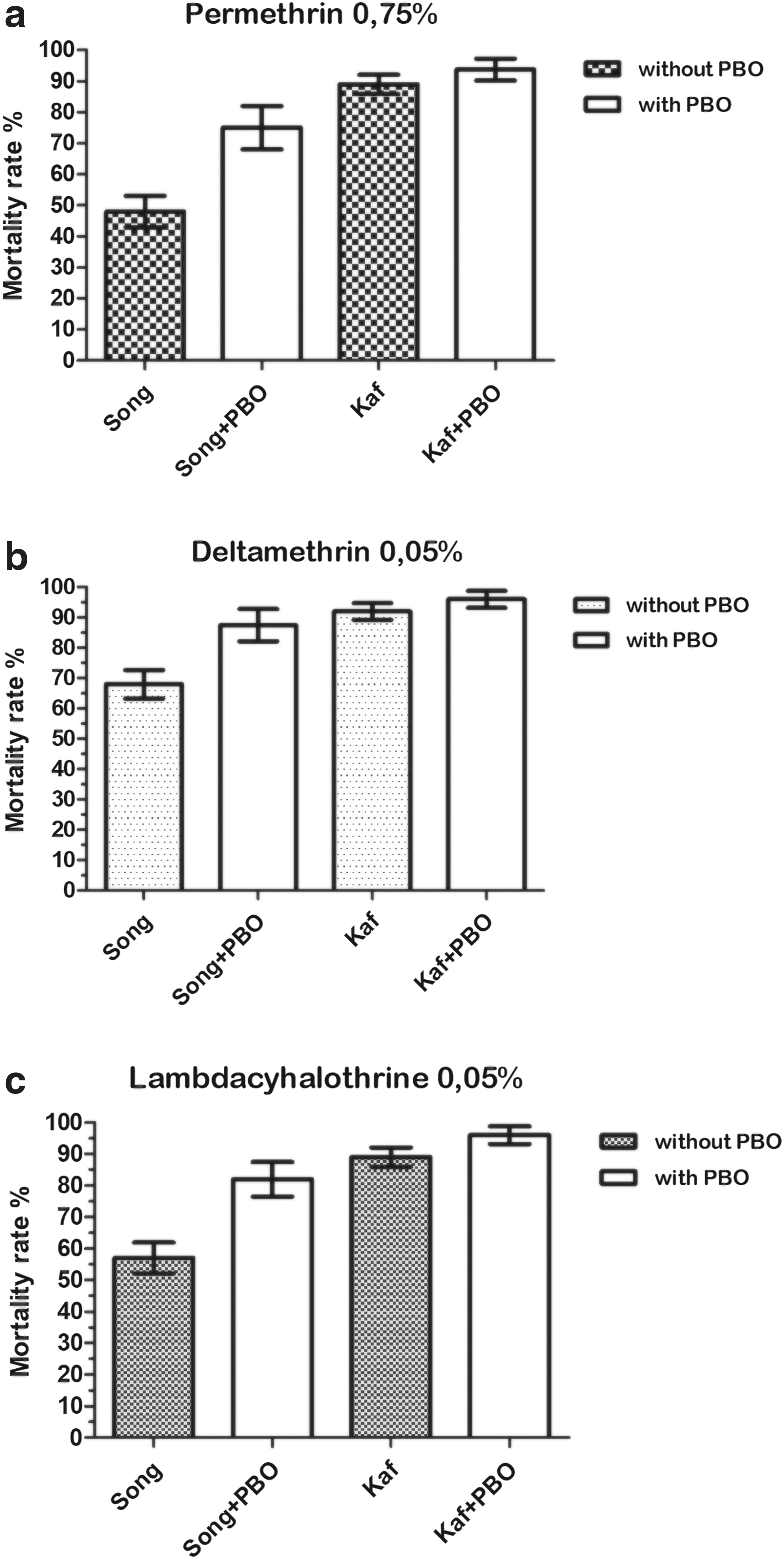
Mortality rate of Aedes aegypti from Songon-Agban and Kaforo to pyrethroids, before and after 1 h of exposure to PBO:
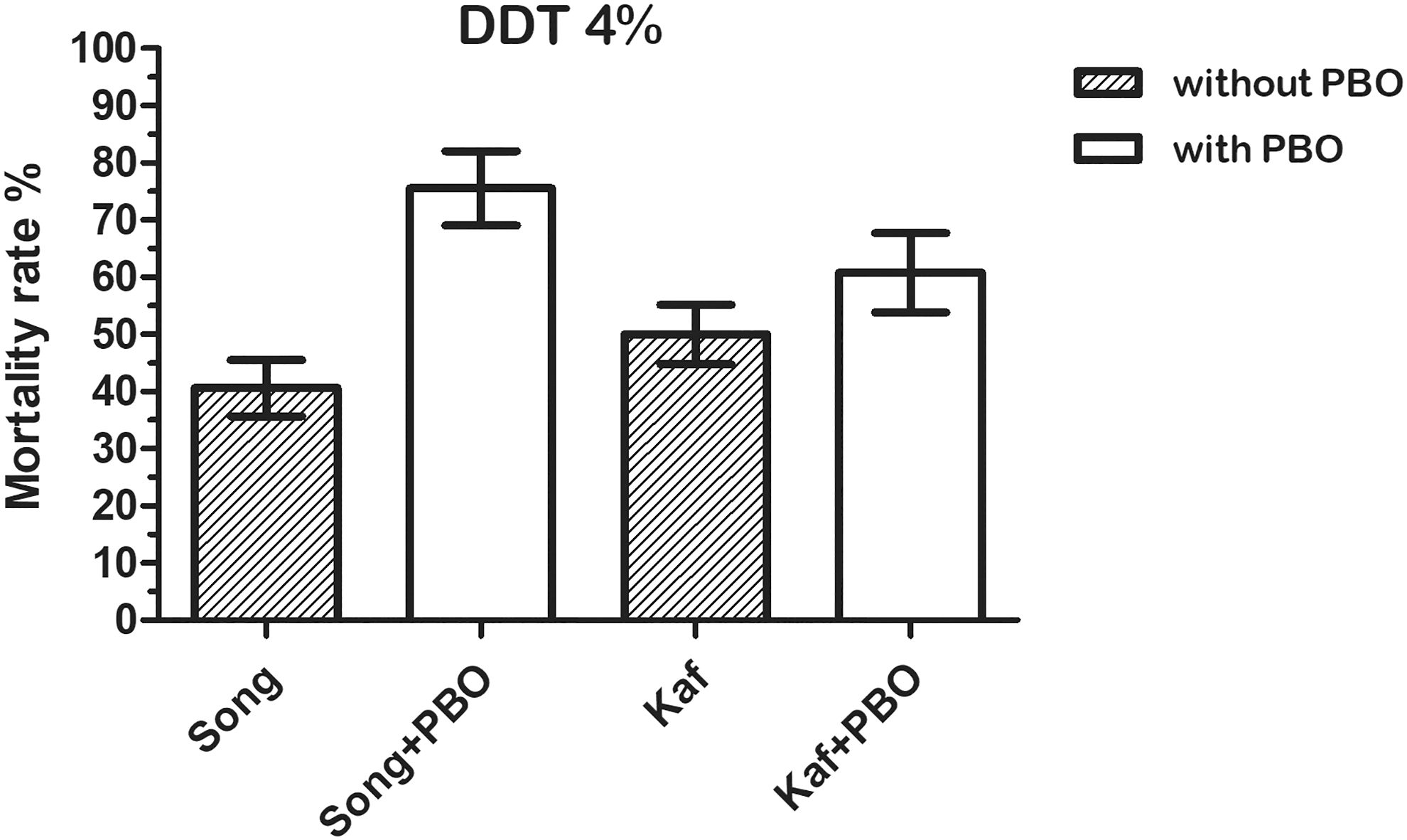
Mortality rate of Aedes aegypti from Songon-Agban and Kaforo to DDT, before and after 1 h of exposure to PBO. Error bars present the 95% confidence interval. DDT, dichlorodiphenyltrichloroethane; Kaf, Kaforo; Song, Songon-Agban.
Involvement of detoxification enzymes in the resistance of Ae. aegypti
We analyzed ∼60 Ae. aegypti females per population to assess their enzymatic activities. Figure 4 shows the distribution of enzymatic activity of NSE (α- or β-naphthyl acetate), GST, and MFO (cytochrome P450) in each Ae. aegypti population compared with the reference susceptible strain SBE.
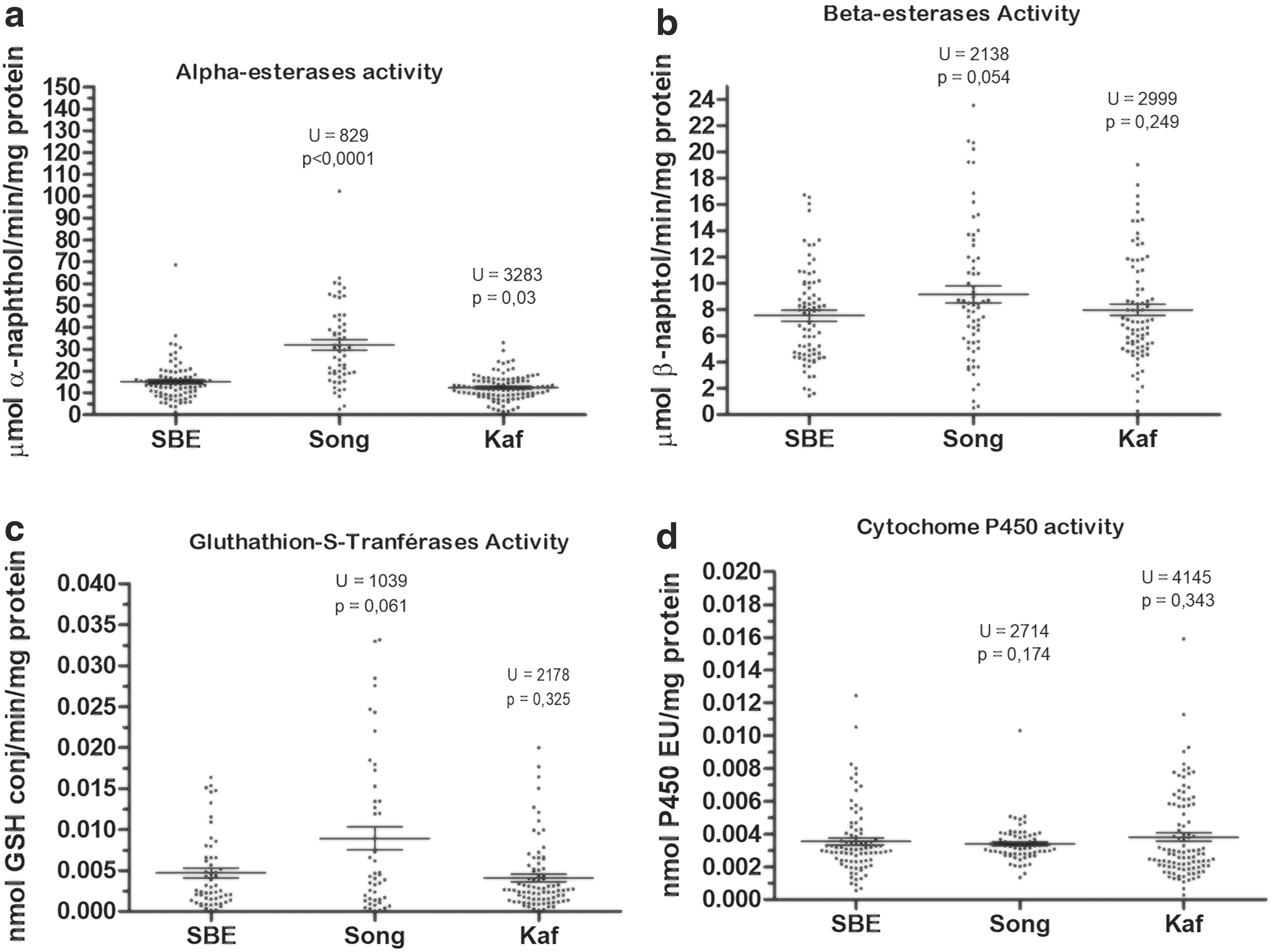
Activity profiles of nonspecific α- and β-esterases
The mean α-naphthyl acetate activities were significantly higher in Ae. aegypti from Songon-Agban (32.08 μmol α-naphthol/min/mg protein) than in the reference SBE strain (15.18 μmol α-naphthol/min/mg protein) (Mann–Whitney test: U = 829; p ˂ 0.0001) (Fig. 4a). Whereas in Kaforo, α-naphthyl acetate activities were significantly lower (12.57 μmol α-naphthol/min/mg protein) than those in the reference SBE strain (U = 3483; p = 0.03).
The other enzymatic activities: β-naphthyl acetate, GST, and cytochrome P450 of the Ae. aegypti populations from our study sites, were significantly similar to the reference SBE strain (p > 0.05).
The mean β-naphthyl acetate activities were 9.16 and 7.99 μmol β-naphthol/min/mg protein in Ae. aegypti samples from Songon-Agban and Kaforo, respectively, and 7.54 μmol β-naphthol/min/mg protein for the reference SBE strain (Fig. 4b).
For GST, the mean was 0.008 and 0.004 nmol GSH conj/min/mg protein in Ae. aegypti samples in Songon-Agban and Kaforo, respectively, and 0.004 nmol GSH conj/min/mg protein for the reference SBE strain (Fig. 4c). However, this activity was significantly higher in Ae. aegypti samples in Songon-Agban than in Kaforo (U = 1479; p = 0.017).
For oxidases, the amount of Cytochrome P450 was equal in the Ae. aegypti populations from both study sites and the reference SBE strain (0.003 nmol P450/mg protein) (Fig. 4d).
Discussion
In Côte d'Ivoire, Ae. aegypti is the main vector of arboviruses, including yellow fever and dengue, and represents a real threat to public health. Especially for the dengue fever, because of the current lack of treatments, the prevention mainly relies on vector control largely based on insecticide treatments during epidemics (MHPHS, 2017; World Health Organization, 2010). Most of the data available on insecticide susceptibility of Ae. aegypti and the control interventions of arbovirus diseases are focused on urban areas, particularly in the Abidjan City.
Unfortunately, the periurban and rural areas of Côte d'Ivoire are underdocumented. The knowledge about insecticide susceptibility of Ae. aegypti populations in these environments is essential to select for the most efficient vector control tools. In this study, we aimed to explore this gap by assessing the susceptibility of Ae. aegypti to different insecticides used in public health and the role of detoxification enzymes as resistance mechanisms. Ae. aegypti was sampled from two agricultural sites in periurban and rural areas.
Bioassays showed resistance of Ae aegypti to pyrethroids and DDT in our two study sites. The resistance status of Ae. aegypti to deltamethrin and DDT was already reported in several neighborhoods of Abidjan city (Guindo-Coulibaly et al., 2014; Konan et al., 2021). The resistance to deltamethrin observed in Ae. aegypti is a main concern since it is usually the first-line insecticide for space sprays during outbreaks (Amelia-Yap et al., 2018; Fernando et al., 2020; Marcombe et al., 2011; World Health Organization, 2009).
Moreover in Côte d'Ivoire, the long-lasting insecticidal nets (LLINs) impregnated with pyrethroids such as deltamethrin are the main tools for the prevention of malaria used by the National Control Program (World Health Organization, 2015). LLINs could also contribute to increased selection pressure on mosquito populations (Meiwald et al., 2022; Sovi et al., 2020).
Insecticide resistance can also result from the use of insecticides against crop pests so we conducted our study in two different agricultural settings. The use of insecticides against insect pests may cause an indirect selection pressure on mosquito species (Cissé et al., 2015; Koffi et al., 2012), but this is also the case of other agricultural pollutants such as herbicides, fertilizers, or fungicides (Nkya et al., 2014). This was observed among Ae. aegypti populations from a vegetable-growing area in Ouagadougou and Bobo-Dioulasso cities in Burkina-Faso (Namountougou et al., 2020).
Previous studies in Côte d'Ivoire showed a strong insecticide resistance of An. gambiae populations in rice- and vegetable-growing areas (Ahoua et al., 2012; Camara et al., 2018; Zoh et al., 2018), and cotton-growing areas (Koffi et al., 2013; Zogo et al., 2019). Ae. aegypti and An. gambiae are not exposed to the same selection pressures at larval stage since they do not share the same breeding sites. However, the presence of pesticide residues on vegetation where adults are resting or in small containers and tires that are present within or close to crop fields may exert a significant selection pressure on Ae. aegypti populations.
In this study, we showed the involvement of metabolic mechanisms in the pyrethroid resistance of Ae. aegypti. Pre-exposure to PBO of Ae. aegypti from Songon-Agban restored a substantial part of susceptibility to the three pyrethroids. As PBO is known to be an oxidase and esterase inhibitor (Khot et al., 1998), the pyrethroid resistance observed in the Ae. aegypti population of Songon-Agban suggests the involvement of these enzymes. These results were confirmed by biochemical assays showing high level of α-esterase activity in Songon-Agban. Some studies across Africa described the involvement of esterases in the resistance of Ae. aegypti to pyrethroids.
The acute activity of detoxifying enzymes, including α and β-esterases, has been reported in deltamethrin-resistant Ae. aegypti in Burkina-Faso (Namountougou et al., 2020). Ngoagouni et al. (2016) showed the involvement of β-esterases in deltamethrin resistance at Ae. aegypti and Aedes albopictus in the Central African Republic. Similar results have also been observed outside Africa, in Asia (Leong et al., 2019), America (Contreras-Perera et al., 2020), and Europe (Seixas et al., 2017). PBO did not fully restore the pyrethroid susceptibility of Ae. aegypti from Songon-Agban, then kdr mutations likely contributed to resistance.
The involvement of kdr resistance was confirmed by the RR. In our two study sites, RR50 and RR95 for KdT, although <3, were significant. This result suggests the presence of kdr mutation(s) at low frequency in two Ae. aegypti populations. Pyrethroids resistance of Ae. aegypti involves the presence of two types of mechanisms in the Songon-Agbna area (metabolic resistance and kdr mutation(s)) and to one in Kaforo (kdr mutation(s)). The different kdr mutation genes involved in pyrethroid resistance if Ae. aegypti should be investigated in further studies. A recent study identified the three kdr mutations V410L, V1016I, and F1534C with a high frequency of the F1534C in near populations of Ae. aegypti from Abidjan (Konan et al., 2021).
The levels of alpha esterase and GST activities were significantly higher in the Ae. aegypti populations of Songon-Agban than in Kaforo. These results show that the insecticide treatments in the vegetable-growing area probably selected more for resistance than in the cotton-growing area. Previous study was conducted on the susceptibility of An. gambiae from different agrosystems in Cote d'Ivoire, and showed that the pesticides were used at repeated and overdosages on rice and vegetable crops compared with other agrosystems such as cotton (Fodjo et al., 2018).
DDT resistance was detected in both populations. It is probably due to mechanisms that confer crossresistance with pyrethroids. DDT resistance of Ae. aegypti is known to be spread worldwide in Americas, South east Asia, and West Africa (Moyes et al., 2017).
The overexpression of α-esterases and the absence of crossresistance with carbamates indicated that the chlorpyrifos-methyl resistance observed in Songon-Agban is mainly due to metabolic mechanisms. Although the effect of PBO was not tested for this insecticide in our study, many studies have demonstrated that NSEs may be involved in phenotypic resistance to organophosphates (Hemingway et al., 2004; Labbé et al., 2017).
In Ae. aegypti, the α-esterase CCEAE3A should be considered as a good candidate for organophosphate resistance since it is widespread, and found to be upregulated and associated with temephos resistance in populations from Asia and America (Moyes et al., 2017). This esterase is also responsible for temephos resistance in Ae. albopictus populations from Europe and North America. Moreover, it was demonstrated that CCEAE3A was able to sequester and to metabolize temephos oxon (Grigoraki et al., 2016).
Biochemical assays showed that there was no overexpression of esterases or other enzymes in Ae. aegypti mosquitoes from Kaforo. Insensitivity of acetylcholinesterase is one of the examples of target site resistance mechanisms, which is the target of organophosphate and carbamate insecticides. However, we cannot involve this mechanism in Kaforo because no carbamate resistance was observed, nor overexpression of esterases and MFO. A previous study showed that organophosphate resistance in Ae. aegypti was closely associated with high esterase, MFO, and ace-1 mutation activity (Muthusamy and Shivakumar, 2015). Resistance to chlorpyrifos-methyl in Kaforo seems to involve other resistance mechanisms that remain to be investigated.
Interestingly, the field populations of Ae. aegypti were susceptible to carbamates (propoxur and bendiocarb) in the two study sites. This result shows that this insecticide class should be a good option for Aedes control in Côte d'Ivoire (Guindo-Coulibaly and Adja, 2014; Konan et al., 2012) contrarily to other African countries such as Burkina Faso (Sombié et al., 2019) and Ghana (Owusu-Asenso et al., 2019) where Ae. aegypti is already resistant to carbamates.
Conclusion
This study revealed that Ae. aegypti populations are resistant to pyrethroids, DDT, and chlorpyrifos-methyl in vegetable- and cotton-growing areas of Côte d'Ivoire. The study reveals the involvement of detoxification enzymes (α-esterases) and likely kdr mutations in insecticide resistance within the most resistant populations of Ae. aegypti. This situation is a real threat for the success of vector control programs, especially in the need for rapid response during emerging dengue epidemics in periurban and rural areas in Côte d'Ivoire. It is therefore necessary to take into account agricultural practices in the management of resistance of dengue vectors.
Knowledge of resistance distribution and underlying mechanisms helps in making rational decisions in selection of appropriate and effective insecticides (Dusfour et al., 2019). In further studies, the different kdr mutations genes involved in the pyrethroid resistance of Ae. aegypti should be investigated in various locations from Côte d'Ivoire.
Footnotes
Acknowledgments
The authors are grateful to the Institut Pierre Richet and Medical and Veterinary Entomologic Centre for Cote d'Ivoire for its technical assistance. They thank the health authorities, the local authorities, and the residents of the study areas and the entomological team who worked on this study.
Authors' Contributions
A.M.A. and. Y.M.-A.E.K. designed conceptualization; Y.M.-A.E.K., D.D.Z., D.F.T., and K.F.A. provided software and data analysis; A.M.A., F.C., and N.G.-C offered validation; Y.M.-A.E.K. and M.D.S.K. conducted investigation; Y.M.-A.E.K. assisted with writing—original draft preparation; A.M.A., F.C., N.G.-C., Y.M.-A.E.K., D.D.Z., M.A.C.S.-K., and D.F.T. supported with writing—review and editing; A.M.A. and A.Y. performed supervision; A.M.A. assisted with project Administration; A.M.A. and F.C. contributed to funding acquisition. All authors have read and agreed to the published version of the article.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This research was integrated to the project RESIS-ARBO. This project was funded by Naval Health Solidarity Fund and Merieux Fund, and Strategic Support Program for scientific Research.