
Abstract
Australian bat lyssavirus (ABLV) is a negative-sense, single-stranded RNA rhabdovirus capable of causing fatal acute encephalitis in humans with similar pathogenesis to its closest serologic relative, rabies virus (RABV). In this review, we describe emergence and classification of ABLV, its known virology, reservoirs, and hosts, as well as both the pathogenesis and treatment approaches currently employed for presumed infections. ABLV was first identified in New South Wales, Australia in 1996 and emerged in humans months later in Queensland, Australia. Only five known bat reservoirs, all of which fall within the Pteropus and Saccolaimus genera, have been identified to date. Although ABLV antigens have been identified in bats located outside of Australia, the three known human ABLV infections to date have occurred within Australia. As such, there remains a potential for ABLV to expand its presence within and beyond Australia. ABLV infections are currently treated as if they were RABV infections by administering neutralizing antibodies against RABV at the site of the wound and employing the rabies vaccine upon possible exposures. Due to its recent emergence, there is still much left unknown about ABLV, posing concerns with how to safely and effectively address current and future ABLV infections.
Initial Discovery of Australian Bat Lyssavirus
Australian bat lyssavirus (ABLV) was inadvertently first identified in June of 1996 by researchers searching for the natural reservoir of Hendra virus (HeV) (Fraser et al., 1996; Hanna et al., 2000; Weir et al., 2014a). While testing for serological evidence of HeV in two female black flying foxes (Pteropus alecto), unexpected characteristics were observed within the bats' isolated brain tissues that were not indicative of an HeV infection. One of these female bats had exhibited abnormal, heightened aggression and contained both cytoplasmic inclusion bodies and remnants of lyssavirus nucleoprotein antigens within her brain tissue. The brain tissue isolated from the other female bat, which was euthanized in 1996 for its inability to fly, displayed signs of nonsuppurative encephalitis and the presence of lyssavirus nucleoprotein antigens. Both cytoplasmic inclusion bodies and nonsuppurative encephalitis are characteristic of infections caused by lyssaviruses (Weir et al., 2014a).
To identify this unknown lyssavirus, isolated viral nucleoproteins (N) from the two bats were both sequenced and evaluated for recognition by various monoclonal antibodies known to neutralize different lyssavirus nucleoproteins (Fraser et al., 1996; Weir et al., 2014a). The antibody testing results indicated that the nucleoproteins were serologically distinct from rabies virus (RABV) and other known lyssaviruses. Sequence analysis determined that the proteins shared 92% sequence similarity with RABV, supporting the discovery of a closely related but distinct new lyssavirus: ABLV.
Identification and Known Cases of ABLV in Humans
As of November 2022, only three humans have had confirmed ABLV infections, and all three cases were ultimately fatal (Table 1).
Summary of Confirmed Cases of Human Australian Bat Lyssavirus Infections
The information pertaining to each confirmed case of ABLV disease is described. The ABLV variants responsible for infection in each human case were either determined through PCR analysis or deduced by which bats were most likely in direct contact with each infected human. The confirmation tests listed represent which tests confirmed an ABLV infection by taking samples from each infected human in the hospital.
ABLV, Australian bat lyssavirus.
October 1996
The first confirmed case of ABLV in a human occurred in 1996 in Rockhampton, Queensland, where a 39-year-old animal handler suffered scratches over several weeks from handling bats, including contact with a yellow-bellied sheath-tailed bat (Saccolaimus flaviventris) in her care (Allworth et al., 1996; Samaratunga et al., 1998; Weir et al., 2014a). She was admitted to the hospital 3 days after experiencing headaches, vomiting, fever, and lightheadedness. Within days of admission, her condition dramatically worsened to complications with swallowing, uncontrollable drooling, and double vision. Testing of cerebrospinal fluid and serum from the patient revealed both the presence of anti-lyssavirus antibodies and PCR confirmation of ABLV N gene (Allworth et al., 1996; Weir et al., 2014a).
Within 11 days of admission, she was dependent upon a ventilator for respiration, unresponsive to external stimuli, and exhibited hyperthermia. Twenty days after onset of symptoms, she passed away from severe cerebral damage, complete paralysis, and diffuse encephalitis (Allworth et al., 1996; Weir et al., 2014a).
November 1998
The second confirmed case of ABLV in humans occurred in a 37-year-old woman who had been bitten by a flying fox (Hanna et al., 2000). She was admitted to the hospital, ∼27 months later, with a fever, difficulty swallowing, sore throat, and vomiting, which had been sustained over the previous 5 days. Upon admission, her symptoms rapidly deteriorated into severe muscle spasms, slurred speech and mental decline, unstable blood pressure and body temperature, and ultimately paralysis. Although samples of cerebrospinal fluid and serum were collected for confirmation of an ABLV infection via enzyme immunoassay tests, it was her sample of saliva that revealed an amplified ABLV-specific PCR product (Hanna et al., 2000; Weir et al., 2014a). Shortly after confirming an infection with ABLV, this patient died from acute encephalitis within 19 days of symptoms onset.
February 2013
The last fatal human ABLV infection occurred in an 8-year-old boy admitted to the hospital 2 months after sustaining a scratch on his left-forearm from an unknown species of a flying fox (Francis et al., 2014b; Weir et al., 2014a). Like the prior two cases, he was suffering from a fever, as well as convulsions and abdominal pain. Within 28 days of symptom onset, he died from severe brain damage and acute encephalitis associated with ABLV infections.
Classification and Virology of ABLV
ABLV expresses an 11.8 kb negative-sense RNA (-ssRNA) genome, which is packaged in enveloped, bullet-shaped virions consistent with the known structures of other lyssaviruses (Fig. 1) (Calisher and Ellison, 2012; Gould et al., 2002; Weir et al., 2021; Weir et al., 2014a). Genetic regions which closely align with the five main viral proteins found within lyssaviruses (matrix protein [M], nucleoprotein [N], phosphoprotein [P], RNA-dependent RNA polymerase protein [L], and glycoprotein [G]) have been identified in available ABLV genomes (Gould et al., 1998; Rupprecht et al., 2017; Weir et al., 2021).
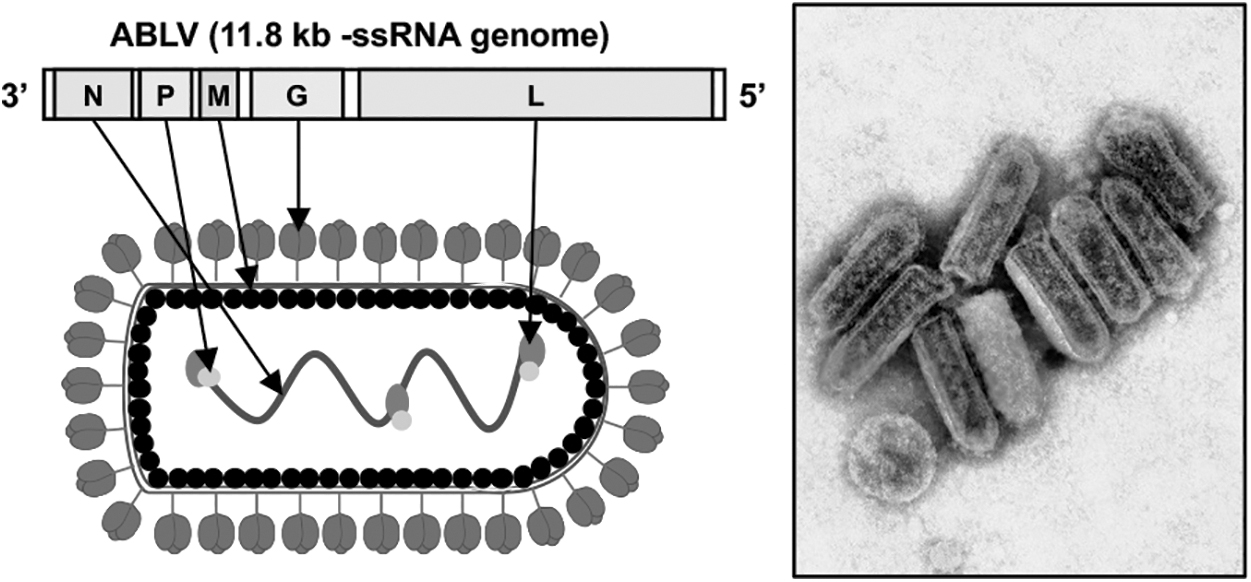
ABLV genome and virion structure. The 11.8 kb negative-sense RNA (-ssRNA) genome of ABLV and a schematic depicting the virion structure with labeled viral gene products indicated by arrows are shown (left). A transmission electron micrograph of ABLV virions is shown at right (adapted from the Commonwealth Scientific and Industrial Research Organization, CSIRO). ABLV, Australian bat lyssavirus.
ABLV is classified in the order Mononegavirales, family Rhabdoviridae, and genus Lyssavirus (Field, 2018; Gould et al., 1998; Guyatt et al., 2003; Warrilow et al., 2002). Within the Lyssavirus genus, there are 17 known viral species separated into 2 phylogroups, several of which are currently ungrouped, based on several distinctions: (1) serologic and genetic components of their glycoproteins, (2) pathogenicity, (3) potential for cross-neutralization by antibodies, and (4) genotype (Badrane et al., 2001; Warrilow et al., 2002). The viral species found within Phylogroup I include Rabies lyssavirus (RABV), ABLV, Aravan lyssavirus (ARAV), Duvenhage lyssavirus (DUVV), Gannoruwa bat lyssavirus (GBLV), Bokeloh bat lyssavirus (BBLV), Irkut lyssavirus (IRKV), Khujand lyssavirus (KHUV), European bat 1 lyssavirus (EBLV-1), European bat 2 lyssavirus (EBLV-2), Kotalahti bat virus (KBLV), and Taiwan bat lyssavirus (TWBLV) (Weir et al., 2021). Phylogroup II contains Lagos bat lyssavirus (LBV), Shimoni bat lyssavirus (SHIB), and Mokola lyssavirus (MOKV).
The lyssaviruses that currently remain uncategorized are Ikoma lyssavirus (IKOV), West Caucasian bat lyssavirus (WCBV), and Lleida bat lyssavirus (LLEBV). All known infections caused by lyssaviruses have the capacity to progress into disease symptoms representative of those manifested in clinical rabies such as fatal acute encephalitis and severe neurological damage (Weir et al., 2021; Weir et al., 2014a). Phylogenetic analyses illustrate that the closest genetic relatives of ABLV are RABV and GBLV, a lyssavirus first identified in bats in Sri Lanka in 2016 (Fig. 2) (Gunawardena et al., 2016).
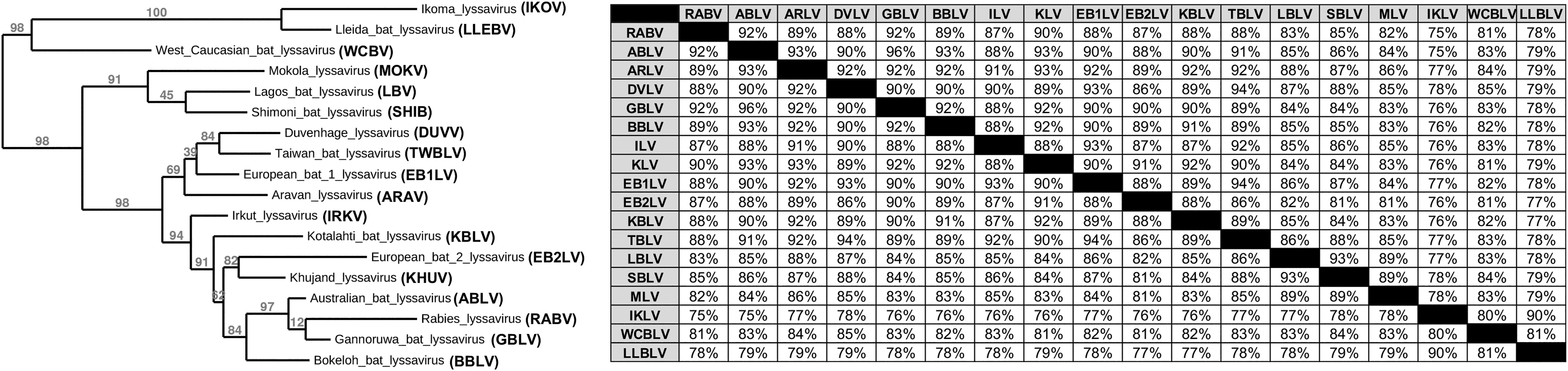
Phylogeny and sequence identity of lyssaviruses based on nucleoprotein (N) sequence. A Bayesian phylogenetic tree generated using nucleoprotein (N) protein sequences. The numbers on the branches represent the estimates for probabilities from the Bayesian inference. The percent identity between lyssaviruses is shown at right.
Very little remains known about ABLV entry and complete pathogenesis. However, due to its high relatedness to RABV, it is believed that ABLV shares many of the same features of replication. Despite great understanding of RABV replication and pathogenesis, there is still much to be desired when it comes to confirming current scientific propositions about ABLV biology. ABLV is believed to attach to host target cells using its G glycoprotein. Despite the genetic and serologic similarities between ABLV and RABV, preliminary research suggests that the proposed RABV host cell receptors (neuronal cell adhesion molecules, nicotinic acetylcholine receptor, or p75 neurotrophin receptor) do not support ABLV entry and replication (Weir et al., 2014a; Weir et al., 2013).
However, current research suggests that ABLV uses distinct receptor(s) or coreceptor(s) abundant in lipid rafts and conserved among mammalian species, but these the specific molecules remain unknown (Fraser et al., 1996; Gould et al., 1998; Warrilow et al., 2002; Weir et al., 2014a; Weir et al., 2013). Testing in cell lines of ABLV variants isolated from the two main genera of bat hosts, Saccolaimus (ABLVs) and Pteropus (ABLVp), suggests that ABLV variant differences in either infectivity or entry likely exist (Weir et al., 2013). ABLV is believed, like most lyssaviruses, to be internalized through clathrin- and dynamin-dependent receptor-mediated endocytic pathway (Weir et al., 2014b).
Upon entry, acidification in the early endosome triggers G-mediated fusion and escape from the endocytic compartment, where the virus utilizes its pre-packaged RNA-dependent RNA polymerase (L) and phosphoproteins (P) to promote and regulate replication and transcription of the viral genome. Recently, analysis of the phosphoproteins (P) in both ABLV variant lineages appears to show clear differences in their roles in immune evasion and nuclear trafficking (Deffrasnes et al., 2021). This might suggest that other differences in ABLV replication events may exist based on variant and host cell type.
Reservoirs and Hosts of ABLV
To date, only five Australian bat species within the genus of Pteropus (suborder Megachoptera) and Saccolaimus (suborder Microchoptera) have been identified as natural reservoirs of ABLV: black flying foxes (P. alecto), little red flying foxes (Pteropus scapulatus), gray-headed flying foxes (Pteropus poliocephalus), spectacled flying foxes (Pteropus conspicullatus), and insectivorous yellow-bellied sheath-tailed bats (S. flaviventris) (Field, 2018; Warrilow et al., 2002). Although only 5 natural reservoirs have been identified, it is believed that all 70+ Australian bat species have the potential of maintaining and propagating ABLV (Merritt et al., 2018; Tidemann et al., 1997; Weir et al., 2014a). There are two genetically distinct lineages of ABLV maintained within the Pteropus and Saccolaimus Australian bat reservoirs: ABLVp and ABLVs, respectively (Guyatt et al., 2003). Nucleotide sequencing suggests that these two variants differ in less than a fifth of their genome.
Yellow-bellied sheath-tailed bats have an extensive range across mainland Australia; whereas the combined geographic range of the four frugivorous Australian flying foxes expands across five of the six Australian states: southern Victoria extending into New South Wales, Queensland, Northern Territory, and Western Australia (Fig. 3) (Guyatt et al., 2003; Weir et al., 2014a). A recent study evaluating samples submitted for testing of ABLV from bats in New South Wales between 2008 and 2021 showed that 4.9% of all submissions tested positive for the virus (O'Connor et al., 2022). While the prevalence of ABLV in this selected population is likely greater than in the general population, this study highlights the continued concern for exposures in humans.

Geographic range of ABLV reservoirs. The five known bat reservoirs of ABLV are located across the southern hemisphere, specifically observed within Indonesia, Papua New Guinea, and Australia. The geographic range for each bat species is indicated by the shading as defined at bottom right.
Although ABLV has only been identified in Australia, little red flying foxes, black flying foxes, spectacled flying foxes, and yellow-bellied sheath-tailed bats are also known to inhabit Papua New Guinea; spectacled flying foxes and black flying foxes have been located within Indonesia as well. The geographic range of gray-headed flying foxes is limited to the south-eastern coasts of Australia. Although the known incidence of ABLV is restricted to Australia, surveillance efforts revealed the presence of neutralizing antibodies in various bat species inhabiting the Philippines and Cambodia (Guyatt et al., 2003; Weir et al., 2014a). Due to the extensive range of these ABLV reservoirs and the discovery of neutralizing antibodies against ABLV in neighboring countries, there is obvious potential for ABLV to expand its presence beyond Australia.
Due to the vast tropism of RABV and the high serologic similarity between RABV and ABLV, there remains much concern about the potential of ABLV to expand its tropism in the future, comparable to that of its serologic relative. Although ABLV infections have not been observed in any other mammal species to date besides Australian bats, three cases in humans, and two cases in horses, one scientific study suggests that domestic cats and dogs have the potential to become infected with ABLVp and will likely recover instead of succumbing to the disease (Annand and Reid, 2014; McColl et al., 2007). Unfortunately, this study could not address the potential of domestic animals spreading ABLVp to humans or explain why domestic animals were less likely to succumb to the disease.
Both lineages of ABLV have also been demonstrated to replicate in various mammalian cell lines (including cell lines from monkey, small rodent, human, horse, and rabbit species) (Weir et al., 2014a). These studies suggest that ABLV has the potential to expand its tropism across more warm-blooded mammalian species in the future with the right mutations (Weir et al., 2014a). The only recorded spillover event in horses also suggests the potential for ABLV to infect other terrestrial mammals in its future, but researchers have not alluded to which terrestrial animal is most likely to harbor the next spillover event (Annand and Reid, 2014). Understanding the extent of potential ABLV hosts is an area of research that deserves the attention as this knowledge would enhance current and future protocols on how to appropriately and effectively minimize the spread, evolution, and transmission of ABLV.
Disease, Pathogenesis, and Treatments for ABLV Infections
ABLV in bats
Preliminary findings indicate that there is a high potential for cross-species transmission within bat populations (Weir et al., 2014a). Infected bats may be either symptomatic or asymptomatic. However, if an infected bat is showing symptoms, it is common to observe abnormal vocalization, sudden and unpredictable bouts of aggression, muscle spasms, impaired breathing, convulsions, paralysis, neurological disease, and death (Merritt et al., 2018; O'Connor et al., 2022). As symptoms progress, infected bats may struggle to eat, roost, and maintain weight (O'Connor et al., 2022; Weir et al., 2014a).
Although surveillance efforts are being made in relationship to better understand the extent of ABLV in bats, the disease progression associated with ABLV in bats has been determined based on observing sick, orphaned, or injured bats reported by the Australian public, veterinarians, or animal-handlers (Merritt et al., 2018; O'Connor et al., 2022).
ABLV in horses
Horses are not currently considered natural reservoirs of ABLV; instead, horses are simply recognized as susceptible hosts from a single known spillover event. In May 2013, two horses in Queensland, Australia were diagnosed and euthanized from an infection caused by an ABLVs variant (Annand and Reid, 2014; Weir et al., 2014a). The first case involved an 18-month-old female which initially struggled with coordination and movement of her hindlimbs. Over the next few weeks, her initial ataxia progressed; she became hypersensitive to stimulation and less inclined to interact with other horses. By 54 h of symptom onset, she was unable to stand and exhibited hypersalivation, rapid eye movement, thrashing of head and neck, and uncontrollable seizing (Annand and Reid, 2014; Weir et al., 2014a).
The second case, involving a close contact of the first case, began showing signs of hindlimb ataxia and behavioral changes the morning after the first horse exhibited symptoms (Annand and Reid, 2014; Weir et al., 2014a). Within 12 h of symptoms onset, this 18-month-old male horse displayed head pressing (an indication of neurologic damage in horses), fear of light, fever, dilated pupils, rapid heartbeat, and an abnormally wide stance. Within 54 h of symptoms onset, his symptoms mirrored the first case. Due to the significance and severity of their disease, both horses were euthanized.
ABLV in humans
Similar to RABV, it is believed that ABLV cannot be contracted via bat feces, urine, or blood, although further research may help with solidifying these beliefs (Merritt et al., 2018). Instead, ABLV is transmitted directly through either a bat bite or scratch that punctures the skin. There is also potential for ABLV transmission should infected bat saliva come in direct contact with mucosal membranes lining the eyes, nose, or mouth. Although there is potential for cross-species transmission, ABLV is currently not known to be transmitted from human-to-human, and based on our current understanding of its well-studied, serologic relative, it is highly unlikely for human-to-human transmission to occur in the future of ABLV (Francis et al., 2014a; Merritt et al., 2018).
Due to the similarities between RABV and ABLV, researchers believe that the incubation period for ABLV in humans mimics that of RABV, ranging anywhere between 3 and 8 weeks (Merritt et al., 2018). The three recorded human ABLV infections suggest that there is potential for the incubation period to fall anywhere between 3 weeks and 27 months (Allworth et al., 1996; Francis et al., 2014b; Hanna et al., 2000). Although the reason as to why the incubation period ranged so drastically for the three reported human cases is currently unknown, it may pertain to either the inexperience of ABLV adapting to human hosts, as ABLV is only an emerging pathogen, or the misidentification of the infection's source in the case of the 27-month-long incubation.
Although the primary location of ABLV replication is not known, it is likely that ABLV travels to the host's central nervous system (CNS) via motor neurons or sensory nerves; ultimately, ABLV may enter the nervous system of the host by either crossing the neuromuscular junction or nerve spindles (Lafon, 2005). Once ABLV has reached the nervous system, the disease symptoms associated with prolific infection of the CNS are observed. For ABLV to be successful in its replication, infection, and spread, it must use certain mechanisms to evade the host's immune response. Although the specific mechanisms ABLV uses are unknown, if ABLV pathogenesis resembles RABV pathogenesis, perhaps ABLV is able to efficiently destroy helper T cells arriving at the primary site of infection, preventing infected cells and neurons from inducing apoptosis (Lafon, 2005). This would encourage ABLV to propagate within the host without being disturbed or halted by host defense mechanisms.
Because of the existing genetic and serologic similarities between RABV and ABLV, it is believed the disease associated with ABLV, acute nonsuppurative encephalitis, clinically manifests itself similar to that of RABV (Guyatt et al., 2003). Based on what has been observed through the three recorded human cases of ABLV, ABLV infections begin with experiencing nonspecific signs and symptoms such as fever, headache, vomiting, fatigue, and dizziness (Francis et al., 2014a; Merritt et al., 2018; Weir et al., 2014a). Within a few days, these symptoms may progress into severe muscle spasms, difficulty swallowing and speaking, general discomfort, and cyclic bouts of aggression, confusion, and lucidity.
Shortly thereafter, the infected person may experience convulsions, paralysis, ventilator-dependence, severe neurological damage, encephalitis, and death (Allworth et al., 1996; Francis et al., 2014a; Hanna et al., 2000; Merritt et al., 2018; Weir et al., 2014a). Unfortunately, once symptoms associated with infection of the CNS begin, the disease progresses rapidly, and doctors are unable to offer more than palliative care. Death is inevitable at this point of the disease.
There are no therapeutics or vaccines explicitly used against ABLV today (Francis et al., 2014a). Proper management and treatment of potential exposure to ABLV are determined based on the individual's current immunization status, or lack thereof, against RABV, and the extent of exposure (Merritt et al., 2018). Upon suspected exposures to bats, it is recommended that the wound be washed with soap and water for at least 5 min after exposure, applying a virucidal antiseptic (i.e., alcohol or povidone-iodine) afterward (Francis et al., 2014a). Once the wound has been sufficiently cleaned, postexposure prophylaxis (PEP) should be administered. PEP consists of inoculating the wound multiple times, as much as possible, with either human rabies immunoglobulin (HRIG) or equine rabies immunoglobulin (ERIG) in hopes of neutralizing the present virus (Francis et al., 2014a; Weir et al., 2021).
HRIG and ERIG are antibodies known to successfully inhibit RABV, and due to the serological commonalities between RABV and ABLV, they have also been used in attempts to minimize the threat of ABLV after potential exposure. If these antibodies are administered appropriately and readily after ABLV exposure, the hope is that the virus will not be able to reach the host's CNS and ultimately result in the infected individual's death. In addition to the use of HRIG and ERIG, the current rabies vaccine is believed to be efficacious in treating ABLV infections and should be administered on days 0, 3, 7, and 14 postexposure for unvaccinated individuals, on days 0, 3, 7, 14, and 28 for immunocompromised individuals, and on days 0 and 3 for previously vaccinated individuals (Francis et al., 2014a; Guyatt et al., 2003; Hooper et al., 1997; Merritt et al., 2018).
Given the high number of confirmed exposures each year to bats, which have the capacity to harbor ABLV, it is believed that these precautionary measures have been largely effective at preventing other fatal infections in humans.
Ecology and the Potential for Evolution of ABLV
Before the emergence of ABLV in 1996, Australia and Antarctica were the only two continents without RABV-like viruses (Field, 2018). Since its emergence, ABLV has consistently been detected within Australian bat populations, resulting in its current endemic status (Iglesias et al., 2021; Merritt et al., 2018). Through surveillance testing, researchers have found that ABLV is more prevalent in younger, vulnerable bat populations as opposed to adult, healthy populations (Field, 2018; Merritt et al., 2018; Weir et al., 2014a; Weir et al., 2013). Researchers estimate that <1% of healthy reservoirs are affected by ABLV, whereas anywhere between 5% and 17% of injured, sick, or orphaned bats succumb to ABLV infections. Although only recently have people become aware of ABLV, it is believed that ABLV has maintained a historic presence in Australian bat populations (Field, 2018).
Despite efforts made toward increasing awareness on ABLV among the Australian public, one surveillance study illustrates a four-fold increase in administering PEP in New South Wales between 2007 and 2011; the increase in PEP administration alludes to the potential increase in exposure to ABLV in recent years, which is concerning in relationship to rates of transmission and spread throughout the years to come (Kardamanidis et al., 2013; Quinn et al., 2014). Although extensive surveillance testing has not been conducted in other Australian territories to determine the prevalence of human-bat interactions, it can be assumed that human–bat interactions have been maintained among veterinarians, bat handlers/caretakers, wildlife officers, and ABLV researchers, which may influence the incidence and prevalence of ABLV in humans in the future (Merritt et al., 2018).
There are well-documented observations of indigenous Australians hunting and eating flying foxes en masse (Tidemann et al., 1997). Although there are no reported rabies-like illnesses associated with the hunting, preparing, and consumption of flying foxes among this population inhabiting northern Australia, there is potential some of the prepared bats were infected with or maintained ABLV due to its historic presence in Australia (Field, 2018). These common, historic human–bat interactions over time might have provided ABLV with the opportunity to evolve and adapt in ways that allow for its transmission to humans. With time, continued exposure to humans, and each viral replication cycle, ABLV will continue to explore its genetic space and may find mutations which dramatically impact its tropism or pathogenesis.
From what is known about the potential host receptor of ABLV, there remains the possibility of ABLV acquiring the right mutation to jump into humans due to potentially high conservation among mammalian species (Weir et al., 2014b; Weir et al., 2013).
In addition to increased risk of exposure, human environmental impacts such as deforestation and urbanization are also likely to influence the future ecology of ABLV-infected bats and their interactions with humans (Tait et al., 2014; Tidemann et al., 1997). Clearing vegetation reduces familiar, available food sources and space for bat roosting. When the bats are displaced from their original habitats, they are forced to migrate and settle elsewhere, resulting in their increased presence within urban, residential areas highly populated with humans. The displacement of flying fox camps has vast potential to increase human-bat interactions which in turn increases the potential of humans contracting ABLV, especially since displaced bats have a higher chance of becoming orphaned, injured, or sick due to the heightened stress and pressure they experience through this displacement.
Over the last few decades, changes in bat habitats have changed drastically due to human interference, and these unsolicited, forced lifestyle changes have induced grave stress in bats, providing ABLV with an opportunity to increase its potential prevalence by preying upon vulnerable, susceptible bats with strained immune systems (Tait et al., 2014; Tidemann et al., 1997).
Final Thoughts
There remain significant gaps in our current knowledge and understanding of ABLV biology and ecology. Much of our efforts to understand and treat ABLV remain centered around our knowledge of RABV. While there has remained only three confirmed human cases to date, suggesting that current approaches may be working, clinicians and health care providers may be limited in their ability to treat current infections and monitor/prevent the potential of future infections should sudden changes in the tropism or pathogenesis of ABLV were to occur. There also remains limited active, targeted surveillance research done to better understand the extent of ABLV reservoirs and hosts within, and especially beyond, Australia. Perhaps, ABLV is more prevalent than initial research suggests due to the high potential of going unnoticed by health officials in countries with less access or awareness (Tidemann et al., 1997).
What research has been done reveals the presence of neutralizing antibodies in a variety of bat species, illustrating the potential expansive presence of ABLV in both Australia and neighboring countries (Field, 2018; Iglesias et al., 2021). Surveillance efforts made to date suggest the potential of exposing further diversity among ABLV isolates with more focused efforts (Field, 2018). These findings warrant the need for more research tailored toward understanding the extent of past, current, and future ABLV infections.
Finally, it is essential for public health that the Australian public be aware of Australian bats as reservoirs to a myriad of harmful pathogens, how to appropriately and safely handle bats, and what to do if bitten or scratched by a bat (Quinn et al., 2014). Although one research study shows that volunteer bat handlers are highly educated on these topics, another research study exposes the lack of understanding within the general public (Quinn et al., 2014; Sánchez and Baker, 2016). Some of interviewed participants from the general public displayed an overconfidence in their ability to handle wild bats (despite being scratched or bitten during their well-intentioned endeavors), an awareness that bats were “germy” without knowing what pathogens bats carry and their potential to cause disease in humans, and an empathetic desire to help any wounded animal (Quinn et al., 2014).
Despite this research being done with a small sample size (n = 16), these findings suggest that there is a need to increase awareness among the general public of how to appropriately and safely address animals maintaining and propagating lethal pathogens. Should the general public remain unaware of the dangers associated with interacting with bats, there may be an increase in recorded ABLV infections and thus an increase in harvesting already limited HRIG and ERIG antibodies, administering PEP, and utilizing already depleted hospital spaces for treatment. It is imperative that education efforts increase in the coming years.
Despite the numerous concerns associated with the future of ABLV mentioned above, one redeeming aspect is that lyssaviruses maintain a slow rate of evolution (Badrane and Tordo, 2001; Calisher and Ellison, 2012). A slower rate of evolution provides the scientific community with more time to wholesomely understand ABLV and its future relationship with current and potential hosts. Lower evolution rates and more time to enhance our knowledge would also result in the virus' decreased potential to acquire favorable mutations, thus lowering the potential for host range expansion, increased pathogenicity or virulence, and heightened transmission rates among the same or different species. The future of ABLV can be concerning, but efforts toward understanding this emerging pathogen will be essential to viral maintenance, treatment, and peace of mind.
Footnotes
Authors' Contributions
Both A.R.Y. and C.C.S. shared equal roles in the conceptualization, writing, review, and editing of this article.
Author Disclosure Statement
The authors declare no conflicts of interest related to the content of this review.
Funding Information
This work was not supported by any external grants or funding sources. This work was supported by a Holcomb Awards Committee (HAC) Grant of Butler University (C.C.S.), and funding and support from both the Butler University Department of Biological Sciences (C.C.S.) and the Butler Summer Institute (BSI; A.R.Y.).