
Abstract
Background:
The growing contact between men and wild animals, caused by the increase in the population in urban centers and the destruction of the habitat of these animals, has been leading to a greater circulation of pathogens between humans and wildlife. Chelonoidis carbonaria, a tortoise found throughout South America, is one of the animals most rescued from animal trafficking and illegal breeding. Considering this situation, this study aimed to verify the occurrence of hemoparasites in C. carbonaria.
Materials and Methods:
Blood samples from 73 C. carbonaria were collected from animals located in (1) a rural commercial breeding unit, (2) an urban zoo, and (3) a center of rescued animal screening. Genomic DNA was extracted from these animals and used in PCRs to detect specific genomic fragments of haemogregarines (i.e., Hepatozoon and Hemolivia), and members of the Anaplasmataceae Family (i.e., Ehrlichia sp. and Anaplasma sp.). Blood samples were screened for hemopathogens by direct microscopy and were used for hematological assays, and serum samples were analyzed to determine the concentration of serum components.
Results:
It was found that 34.2% of the tortoises presented Sauroplasma sp. in their blood samples; these animals showed clinical biochemistry changes that indicate altered liver function. Two zoo animals were positive for Ehrlichia sp. in PCR, and also presented clinical biochemistry and hematological changes.
Conclusion:
The present project is pioneer in the detection of Ehrlichia sp. in C. carbonaria, and was able to identify changes in clinical biochemistry that can be a result of the infection by hemopathogens in this species.
Introduction
Among the protozoan blood pathogens that were already identified in reptiles, the families Haemogregarinidae and Hepatozoidae (Cardoso et al., 2022) are the most studied through molecular and morphological identification, being the genera Haemogregarina and Hepatozoon the most common ones (Telford, 2009). On the contrary, nonpigmented intraerythrocytic inclusions with a spherical or irregular shape, usually seen as vacuoles with peripheral chromatin granules, are also commonly found in reptiles. They have been assigned to the genus Sauroplasma (Piroplasmida: Haemohormidiidae), with diagnosis based on morphometric and morphological characteristics, and no molecular sequences available (Telford, 2009).
Anaplasmosis is a disease caused by bacteria of the genus Anaplasma, with species such as A. platys infecting dog platelets, and A. phagocytophilum found in human leukocytes (MacQueen and Centellas, 2022). However, there is still a lack of research on the importance of these bacteria in reptiles. The Ehrlichia sp. that infect domestic mammals are the most studied ones, such as Ehrlichia canis, Ehrlichia chaffeensis, and Ehrlichia ewingii. Morphologically, they are identified within cytoplasmic vacuoles in mammalian leukocytes, but in reptiles there has not yet been any molecular and microscopic identification, only in vectors collected from these animals (Andoh et al., 2015). All the hemopathogens described above require vectors, such as ticks, for vertebrate infection (Melo et al., 2015). Furthermore, these animals have a significant close contact with humans and little is known about the pathogens they can carry.
Considering the above, the objective of this work was to identify and to classify morphological and molecular hemopathogens in tortoises Chelonoidis carbonaria, which are broadly found in South America in periurban forest fragments and are not only bred as companion animals but used as food by many populations.
Materials and Methods
Structure of the studied population
Samples were collected from 73 animals, of both sexes, with the following median parameters: weight of 2.8 kg, 32.15 cm wide, and 35.5 cm long. All of them were considered clinically healthy according to clinical evaluations carried out by veterinarians specialized in wild animals. These animals were kept in different captivities, being one a rural commercial breeding unit-commercial resources (CR), a center for screening and rescue of animals from illegal property or trafficking-center of triage (CT), and a zoo-center of zoosciences (CZ).
The collection of blood from these animals was allowed by the Authorization and Information and Biodiversity System (SISBio), protocol number 64936-1. An IRB approval and a collection permission was also obtained from the Ethics Committee for the Use of Animals in Research, the School of Veterinary Medicine and Zootechnics of the Federal University of Bahia, under the protocol number 59/2019.
Sample collection
Two milliliters of blood was collected from the animals through jugular venipuncture, using a 3 mL syringe and a 25 × 0.7 needle. Five hundred microliters of blood was placed in a tube with ethylenediaminetetraacetic acid anticoagulant for genomic DNA obtaining, and 500 μL was placed in a tube containing the heparin anticoagulant for hematological analyses; the use of heparin as an anticoagulant was based on the fact that it is more suitable for the morphological analysis of reptile blood cells (Martínez-Silvestre et al., 2011). Another 1 mL of blood was placed in a tube without an anticoagulant for clinical biochemistry analyses. Previous antisepsis was performed at the puncture site with cotton soaked in 70% ethanol.
PCRs
Genomic DNA samples were extracted from the blood of the 73 animals using the PureLink Genomic DNA Mini Kit extraction kit (Thermo Fisher Scientific, Waltham, MA, USA), according to the manufacturer's instructions. DNA samples were quantified and analyzed for purity in a NanoDrop device (Thermo Fisher Scientific).
Genomic DNA samples were tested for the presence of specific genomic material from Hepatozoon sp. (Inokuma et al., 2002), Hemogregarines (Perkins and Keller, 2001; Ujvari et al., 2004), Ehrlichia sp. (Almeida et al., 2013), and members of the Anaplasmataceae family (Breitschwerdt et al., 1998; Spolidorio et al., 2010), using primers and PCR conditions already described in the literature (Supplementary Table S1). PCRs were performed in a final volume of 25 μL, with 1 × buffer, 1.5 mM magnesium chloride, 0.2 mM of each deoxynucleoside triphosphate, 1.0 μM of each primer, 1 U of Taq DNA Platinum (Invitrogen, Carlsbad, CA, USA), and 10 ng of DNA. Amplification took place over 35 cycles with varying annealing temperatures for each primer set.
In all reactions, positive controls (DNA from cell cultures of the pathogen studied, or from confirmed infected animals) and negative controls (ultrapure water, without DNA) were used. PCR products (2 μL) were visualized on a 1.5% agarose gel stained with SYBR Green to confirm the amplification and the presence of a single specific product.
Direct microscopy analysis
For each blood sample collected from each animal, five blood cytology slides were made, which were fixed with methanol and stained with a rapid hematology dye (Panotic) or May–Grunwald–Giemsa. These smears were analyzed under an optical microscope (Nikon, Tokyo, Japan) at 1000 × magnification for morphological and morphometric analyses.
The slides were analyzed focusing on the observation of intracellular structures, based on Telford (2009). The evolutionary forms of Hepatozoon spp. were identified according to O'Dwyer et al. (2013) and Telford (2010). Sauroplasma sp. was identified as described by Svahn (1976) and Telford (2009). In the morphometric calculations, measurements of the length, width, and area of the structures found in the blood cells were made.
Hematological and clinical biochemistry analysis
The hemogram (total red blood cell counts, hemoglobin concentration, and globular volume) and leukogram (total white blood cell counts and differential counts) techniques were performed within a maximum period of up to 8 h after collection, aiming at preserving the cellular characteristics of erythrocytes and leukocytes. The techniques were performed manually, taking into account the difficulty of calibrating automated devices for analysis of wild animal blood samples, especially those that have nucleated erythrocytes.
To determine the total and differential cell counts in each whole blood sample, the following techniques were performed: (1) the number of erythrocytes and leukocytes were obtained using a Neubauer chamber and the Natt-Herrick solution 1:100 as a diluent solution; (2) determination of globular volume, using the microhematocrit technique, with sealed 70-mm-long and 1-mm-diameter capillary tubes, which were centrifuged for 5 min at 12,000 × g; (3) the hemoglobin concentration was determined by the cyanmethemoglobin method, using the Drabkin's solution; and (4) the calculation of the absolute hematimetric indices, such as mean corpuscular volume, and mean corpuscular hemoglobin and mean corpuscular hemoglobin concentrations were obtained by mathematical calculations resulting from the correlation of values of red blood cell counts, concentration of hemoglobin, and determination of the globular volume.
For the leukocyte differential counts, as well as for the evaluation of the morphotinctorial characteristics of the red blood cells, blood smears were done right after the blood collection. Then, the smears were stained with May–Grunwald–Giemsa or Panotic.
The determinations of serum biochemistry parameters were performed using commercial kits (Wiener, Rosário, Argentina). The methodologies were performed according to the manufacturer's recommendations in an automated system (Cb 350i—Wiener, Rosário, Argentina). The parameters evaluated were alkaline phosphatase, serum glutamic oxaloacetic transaminase (SGOT), lactate dehydrogenase (LDH), creatinine, urea, iron, calcium, phosphorus, glucose, albumin, uric acid, cholesterol, and triglycerides.
Statistical analysis
All data were analyzed using the statistical software GraphPad Prism version 6. The first step consisted of descriptive statistics analysis, with the determination of the maximum and minimum values, quantification of the medians and identification of the values present between the second and third quartiles (25–75%), for the hematological and clinical biochemistry results. To determine whether the data followed a Gaussian distribution, the Shapiro–Wilk test was performed with a confidence level of 95% (p < 0.05). Outliers were identified by the Tukey's t-test and Rout's test and, when necessary, were discarded for statistical calculations (values greater than three times the value of Q3). With the objective to determine reference values, the results between the 25% and 75% quartiles were used. All results were evaluated and analyzed as described in the American Society for Veterinary Clinical Pathology standards (Friedrichs et al., 2012).
Results
Molecular and microscopic diagnosis
In the peripheral blood of C. carbonaria, Sauroplasma sp. structures were found, with frequencies of 35% (9/26) in the CR group, 36% (9/25) in the CZ group, and of 32% (7/22) in the CT group (Fig. 1). These parasites presented means of 1.72 μm width and 2.93 μm length, and no changes were observed in the morphology of the host cells. The parasitic forms were observed as small inclusions that were amoeboid-shaped and with no pigments, and they were associated with the presence of peripheric chromatin granules and often ring-shaped with a central vacuole (Fig. 2).
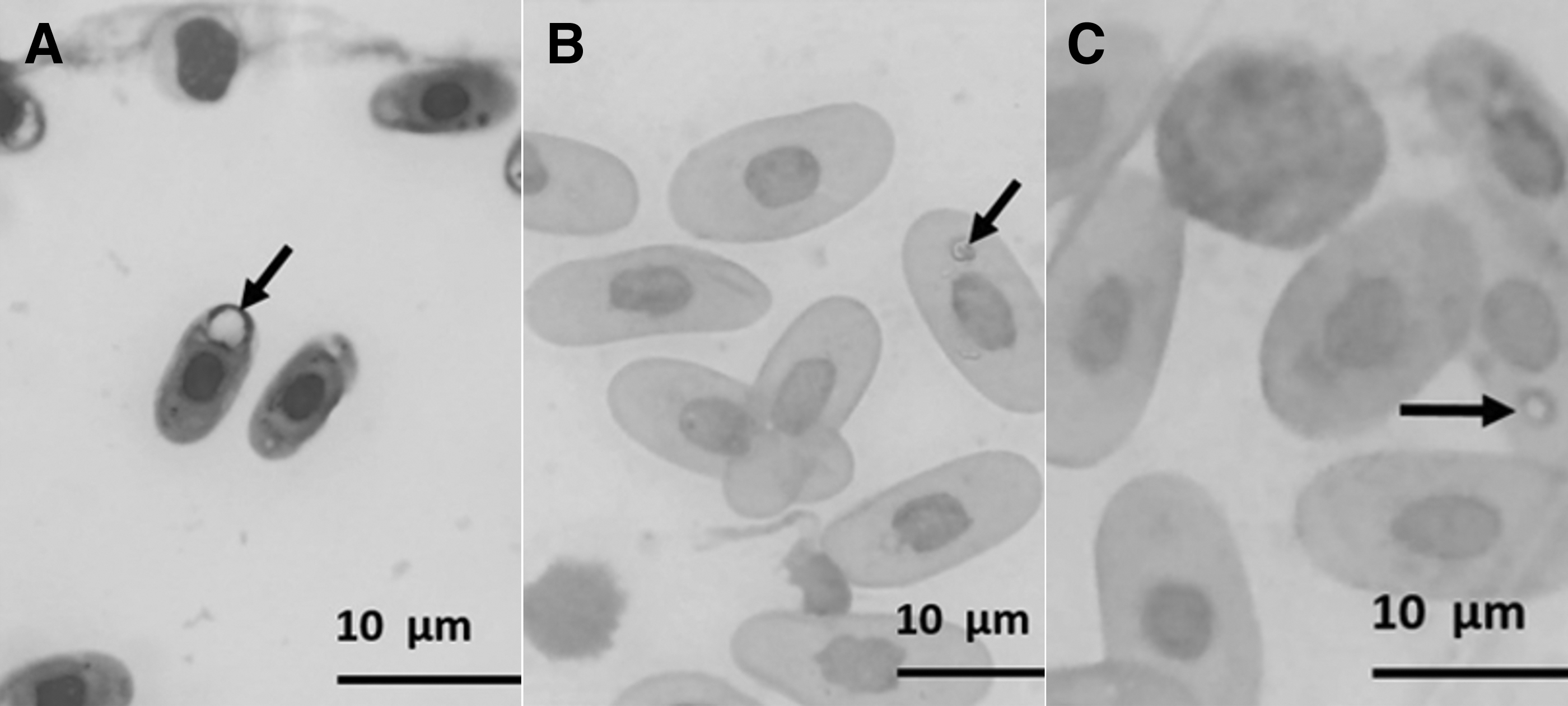
Presence of Sauroplasma sp. in the erythrocytes of Chelonoidis carbonaria (red tortoise). Blood smears stained with Panotic. The structures are indicated by the black arrows.
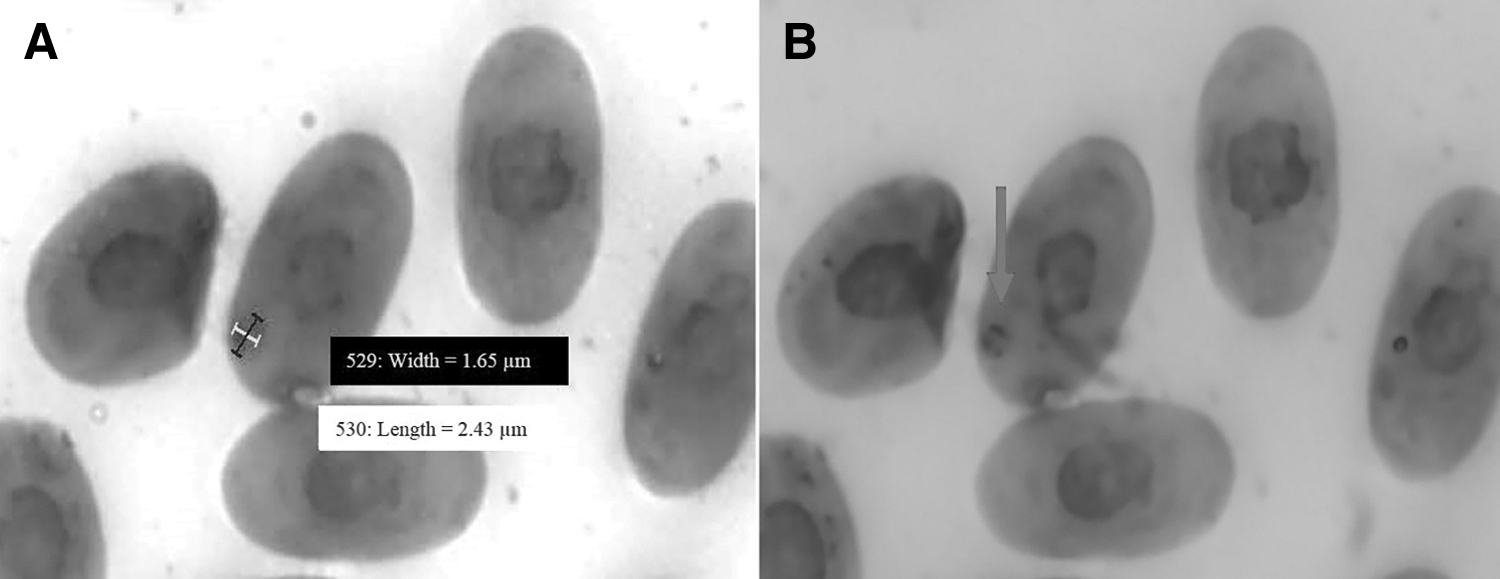
Morphometry of the Sauroplasma sp. structures found in the erythrocytes of Chelonoidis carbonaria (red tortoise). Blood smears stained with Panotic. Measurement of width and length
In the molecular diagnosis of hemopathogens, no positive results were found for the presence of specific genomic material of haemogregarines and bacteria of the Anaplasmataceae family. However, two positive results were found, with amplification of a specific genomic fragment of Ehrlichia sp. in two samples from the CZ group (Supplementary Fig. S1). The bands presented the expected size (400 bp).
Hematological and clinical biochemistry parameters
In this work, hematological and clinical biochemistry values were compared between 25 tortoises positive for Sauroplasma sp. and 48 negative animals. In the erythrogram and leukogram analysis, there were no statistically significant differences in the comparison between positive and negative animals; however, in the clinical biochemistry values, there were significant differences. SGOT, LDH, creatinine, albumin, uric acid, and urea levels were lower in positive animals, when compared with the negative ones, and serum iron presented an opposite result. Quantitative assessments of biochemical parameters are shown in Figs. 3 and 4.

Serum component concentrations in Chelonoidis carbonaria (red tortoise) presenting or not blood Sauroplasma sp. Serum levels of SGOT
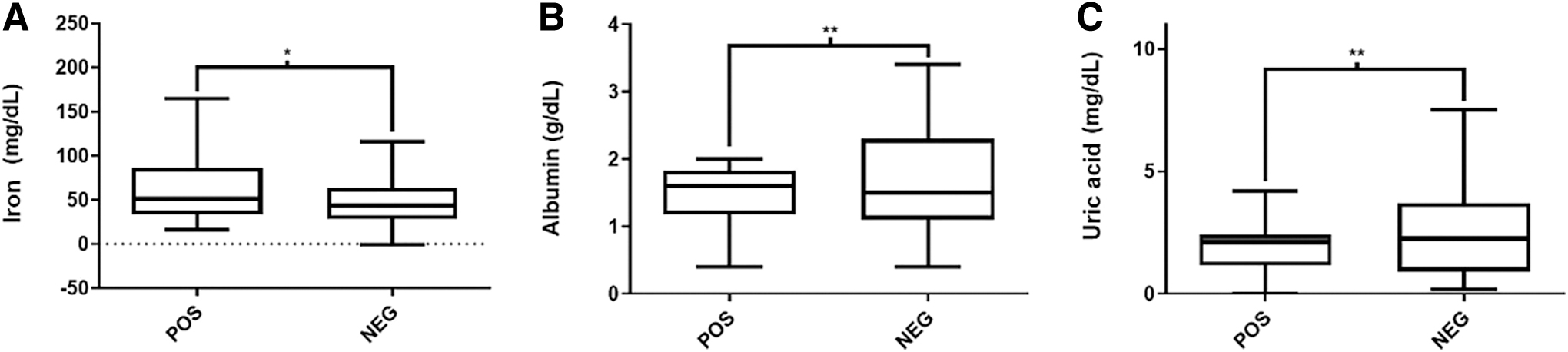
Serum levels of iron
When the hematological values of the two Ehrlichia sp.-positive tortoises were compared with the reference intervals obtained herein for the zoo animals and another one already reported in the scientific literature (Bergamini, 2016), it was found that both animals presented high levels of hemoglobin and high red blood cell counts. The animal 20 presented high counts of total leukocytes and heterophils, but the animal 24, on the contrary, showed low counts of these two cells (Table 1). Regarding clinical biochemistry, the animal 20 presented high levels of all serum elements measured here, with the exception of urea and alkaline phosphatase (Table 2). The animal 24 presented high urea levels only.
Hematological Parameters in Two Chelonoidis carbonaria Infected by Ehrlichia sp.
The reference interval of the zoo group and the reference values determined by Bergamini (2016) are listed.
The symbol “*” stands for altered values, when compared with both the reference intervals.
MCH, mean corpuscular hemoglobin; MCHC, mean corpuscular hemoglobin concentration; MCV, mean corpuscular volume.
Clinical Biochemistry Values in Two Chelonoidis carbonaria (Red Tortoises) Infected by Ehrlichia sp.
The reference interval of the zoo group and the reference values determined by Pereira (2015) and Pereira et al. (2019) are listed.
The symbol “*” stands for altered values, when compared with both the reference intervals.
LDH, lactate dehydrogenase; SGOT, serum glutamic oxaloacetic transaminase.
Discussion
There are few published works on the identification of hemopathogens in C. carbonaria, and it is important to carry out this identification so that possible environmental and even zoonotic risks can be evaluated (Kenny et al., 2004). The tortoise C. carbonaria is a species that has a very proximity to humans, because they are animals that may be consumed as food or bred as companion animals. Some hemopathogens have already been described in Testudines, including viruses (Mashkour et al., 2021) and bacteria (Marenzoni et al., 2015), with protozoa of the order Adelina, families Haemogregarinidae and Hepatozoidae (Goes et al., 2018), and of the order Piroplasmorida, family, Haemohormidiidae (Picelli et al., 2016), the most studied ones.
The detection of Sauroplasma sp. in reptiles has already been described in freshwater chelonians Podocnemis expansa from Brazil (Carvalho et al., 2019, Picelli et al., 2016). In Carvalho et al. (2019), haemogregarine-like structures with a mean size of 1.72 μm width and 2.93 μm length were visualized in erythrocytes, as previously described by Halla et al. (2014). Even considering the fact that the animals did not present clinical signs of any infectious disease, we must correctly diagnose the infection, because factors such as stress, poor diet, and environmental changes can trigger possible pathologies. These parasites have been identified in reptiles for a long time, but they were initially mistaken for artifacts (Telford, 2010), and works such as those by Carvalho et al. (2019) and others describe piroplasm-like parasites recorded in chelonian hosts only morphologically, failing to develop a molecular identification (Picelli et al., 2016).
The absence of studies describing the complete life cycle of these parasites is an aggravating factor for their identification and possible perception of their pathogenicity. Carvalho and Malvasio (2017) noted the possibility that the vectors may be leeches, since in his study the Sauroplasma-positive freshwater chelonians P. expansa were infested by these worms. However, for terrestrial hosts, that is, turtles, snakes, and lizards, the transmission by ticks is more plausible.
There are reports of haemogregarines and Hepatozoon sp. in tortoises and/or sea turtles (Pessoa et al., 2016). In our work, it was not possible to detect these protozoa through the search for gamont in blood smears, even when a sensitive and specific molecular technique was used to detect these pathogens. The same occurred for the detection through PCR of members of the Anaplasmataceae family. This may have occurred due to the difficulty of finding these pathogens in the hosts, despite their easy detection in ectoparasites (Fonseca et al., 2020, Sumrandee et al., 2015). Haemogregarine and Hepatozoon sp. hardly cause clinical signs in reptiles, and the parasitemias may be so low that it is not possible to visualize them in blood smears (Telford, 2010).
It was possible to detect in a pioneer way Ehrlichia sp. in blood samples from two C. carbonaria through PCR using primers for specific amplification of part of the dsb gene. In general, the studies were associated with the finding of Ehrlichia in ticks, without examining the host. Andoh et al. (2015) and Kenny et al. (2004) diagnosed Ehrlichia sp. in an Amblyomma sparsum tick that was parasitizing a leopard tortoise Geochelone pardalis from Africa. Qiu et al. (2021) found Ehrlichia sp. in Amblyomma geoemydae that was parasitizing a Chinese tortoise Cuora flavomarginata.
Our work was able to detect Ehrlichia sp. in a possible vertebrate host, thus generating a new factor to be considered when studying reservoirs of this bacterium, and even guiding further works on the possible transmission to other animals and humans through vectors that are not so specific in relation to the parasitized species (Mubemba et al., 2022, Palomar et al., 2012).
No significant changes in hematological parameters were observed for animals showing blood structures suggestive of Sauroplasma sp., but there were significant changes in clinical biochemistry values. SGOT, creatinine, uric acid, LDH, albumin, and urea were all present in reduced amounts in the sera from positive animals. This result may first appear to be some kind of contamination of the samples with lymph, and the consequent dilution of the sample, but this is not the case, since blood cell counts were normal. This is a profile very similar to chronic liver failure, mainly due to hypoalbuminemia, and the decrease of other molecules that are markers of liver activity (Rallis et al., 2005). Anyway, the results suggest that the possible infection by Sauroplasma sp. has a decisive influence on the liver function of animals.
There are no studies correlating the infection by Ehrlichia sp. in reptiles with their hematological values, considering that there has never been this identification in animals, only in their ectoparasites. Thus, inferences about the pathophysiological consequences of the disease are difficult to make, as we only have dogs to compare, as canine ehrlichiosis is highly studied and prevalent (Diniz and Moura, 2022). The tortoises that were positive for Ehrlichia sp. presented variable hematological alterations. In animal 24, it was possible to observe a leukopenia associated with a decrease in heterophils, and this decrease may be associated with a hyperacute inflammatory disease, mainly due to their relocation to the tissues and the slow mobilization of blood cells from the bone marrow (Thanasak et al., 2022). The animal 24 also presented polycythemia, suggesting a dehydration profile. Ehrlichia sp.-infected animals may present different hematological alterations (Kakoma et al., 2000).
In dogs, classic hematological alterations occur in the infection by Rickettsiales (Almeida et al., 2013), such as anemia, leukopenia, and/or thrombocytopenia (Diniz and Moura, 2022). In contrast, when considering the clinical biochemistry analysis, the animal 24 only showed changes in serum urea levels, which may also be associated with dehydration; still, a hyperacute phase can be also suggested, on which there was not enough time for the development of significant changes in different parameters.
The animal 20 presented a mild leukocytosis, and an elevation in heterophil counts was also observed. Leukocytosis in reptiles may be associated with stress (Scheelings and Rafferty, 2012) and is seen in acute infectious processes, such as those of bacterial origin (Pessoa et al., 2016). The high concentrations of LDH, iron, phosphorus, glucose, albumin, uric acid, cholesterol, and triglycerides in the serum of the animal 20 may be associated with an acute phase of the disease (Rosser, 2022). We also observed polycythemia, which is usually related to dehydration, as seen for the animal 24.
Conclusions
The present study identified, in a pioneer way, Ehrlichia sp. infecting C. carbonaria chelonians. It also identified Sauroplasma sp. forms in the blood cells of these animals, and we were able to identify clinical biochemical changes in these animals, mostly of hepatic origin. Further studies should focus on a better identification of these Ehrlichia, with isolation in cell culture and the consequent sequencing of the entire genome.
Footnotes
Acknowledgments
The authors are grateful to Francisca Soares (LABIMUNO ICS-UFBA) and Leane Gondim (HOSPMEV UFBA) for technical assistance.
Authors' Contributions
M.A.M.: Conceptualization, experimental design, sample obtaining, and microscopic and molecular analysis. G.C.P., G.M.R., and P.O.M.S.: Sample obtaining and article writing and editing. M.S.F.: Molecular analyses. F.S.M. and M.C.C.A.: Clinical biochemistry analyses. L.A.V.: Data curation and critical revision of the article. R.M. and R.D.P.: Resource obtaining, conceptualization, critical revision of the article, and article editing. All authors read and approved the final article.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This study was funded by the Fundação de Apoio à Pesquisa e Extensão (FAPEX), through continuous resources obtained by extension projects. Marcos Antonio Mendonça, Gustavo Macedo Rodamilans, and Gabriela Cruz Piedade are PhD fellows from Coordenação de Aperfeiçoamento de Pessoal de Ensino Superior (CAPES). R.D.P. is a Technical Development fellow from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq - Proc. 313350/2019-1). The funding agencies had no influence on the development of the studies and the results presented herein.
Supplementary Material
Supplementary Figure S1
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.