
Abstract
Background:
The increasing reports on emerging/re-emerging arboviral disease outbreaks or epidemics in Sub-Saharan Africa have been impacted by factors, including the changing climate plus human activities that have resulted in land cover changes. These factors influence the prevalence, incidence, behavior, and distribution of vectors and vector-borne diseases. In this study, we assessed the potential effect of land cover changes on the distribution and oviposition behavior of some arboviral vectors in Zika forest, Uganda, which has decreased by an estimated 7 hectares since 1952 due to an increase in anthropogenic activities in the forest and its periphery.
Materials and Methods:
Immature mosquitoes were collected using bamboo pots and placed at various levels of a steel tower in the forest and at different intervals from the forest periphery to areas among human dwellings. Collections were conducted for 20 months.
Results and Conclusion:
Inside the forest, 22,280 mosquitoes were collected belonging to four arboviral vectors: Aedes aegypti, Aedes africanus, Aedes apicoargenteus, and Aedes cumminsii. When compared with similar studies conducted in the forest in 1964, there was a change from a sylvatic to a tendency of peridomestic behavior in A. africanus, which was now collected among human dwellings. There was an unexpected change in the distribution of A. aegypti, which was not only collected outside the forest as in previous reports but also collected in the forest. Conversely, A. cumminsii originally collected in the forest expanded its ranges with collections outside the forest in this study. Aedes simpsoni maintained its distribution range outside the forest among agricultural sites. We suspect that land cover changes were favorable to most of the arboviral vectors hence enhancing their proliferation and habitat range. This potentially increases the transmission of arboviral diseases in the area, hence impacting the epidemiology of emerging/remerging diseases in Uganda.
Introduction
The emergence and reemergence of arthropod-borne viruses (arboviruses) over the past two decades has become a global public health (Girard et al., 2020) and socioeconomic burden (Thompson et al., 2020). For example, Zika virus fever whose causative pathogen was first isolated in Uganda in 1947 (Dick et al., 1952), re-emerged and spread to other parts of Africa and beyond (Cao-Lormeau et al., 2014; Heukelbach et al., 2016; Musso et al., 2014). Also, in the recent past, Chikungunya outbreaks have been reported in several African countries beyond rural Tanzania where it was initially detected in 1952 (Bettis et al., 2022). Dengue though more prevalent in Asia, has caused several outbreaks in Africa over the recent past (Mwanyika et al., 2021). Furthermore, yellow fever is another re-emerging arboviral disease that has caused sporadic outbreaks in several Sub-Saharan countries in the past two decades (Oyono et al., 2022).
Other emerging and re-emerging arboviral diseases that have recently impacted the global health system include West Nile fever, Crimean Congo Hemorrhagic fever, Japanese Encephalitis, Rift Valley fever among others (reviewed in Chala and Hamde, 2021).
Several factors have facilitated the emergence of new arboviral diseases and the reemergence of diseases that were previously considered under control in areas where they intermittently reappear in epidemics and beyond known endemic zones. Land cover changes have a significant impact on the distribution and behavioral dynamics of mosquito vectors in general, which ultimately influences epidemiological dynamics in a given region (Afrane et al., 2012; Anyamba et al., 2012; Gould et al., 2017; Oscar Júnior and de Assis Mendonça, 2021; Robert et al., 2020). Anthropogenic encroachment on mosquito habitats especially forests is likely to affect the vectorial traits as well as the interaction between the human population and the vector population of the forest-dwelling mosquitoes (reviewed in Lindahl and Grace, 2015).
Uganda's forests have faced significant encroachment over time due to the need for land for agriculture and settlement for the growing population (Twongyirwe et al., 2018). Consequently, the increasing close human proximity to wild mosquito populations can potentially have epidemiological implications in a country where several index specimens of arboviruses have historically been isolated (reviewed in Mayanja et al., 2021).
In this study, we suggest that landcover changes within the proximity of Zika (among other factors) have a potential effect on the distribution and oviposition behavior of some mosquito arboviral vectors. Zika forest is located within the densely populated central part of Uganda and has consequently reduced in size over the years due to increased anthropogenic activities in the forest and its buffer zone. The forest was extensively used for entomological and arboviral surveillance in the 1960s to 1970s with several arboviral isolations made from the forest, including Zika virus, yellow fever virus, chikungunya virus, rift valley fever virus, and many other arboviral strains (Haddow et al., 1964; Kirya et al., 1977; McCrae and Kirya, 1982). Most of these arboviruses have recently caused outbreaks of arboviral diseases in Africa. For example, yellow fever outbreaks in Kenya (Uwishema et al., 2022), Angola (Barrett, 2016; Rossetto and Luna, 2016), Senegal (Diagne et al., 2021), and Nigeria (Nomhwange et al., 2021) among others. Serological studies have also observed widespread prevalence of Zika virus in Africa (Hill et al., 2019; Kayiwa et al., 2018; Robin and Mouchet, 1975; Sassetti et al., 2018).
Land cover changes, due to encroachment on the forest and its buffer zone, are some of the factors that can potentially modify the ecology and biology of arboviral vectors therein and consequently impact the epidemiology of arboviral diseases in Uganda. Data generated will contribute to information required for effective control and management of emerging and re-emerging arboviral diseases transmitted in Uganda.
Materials and Methods
Study area
Zika forest (32°30′E and 0°7′N) is a raised forest patch (altitude of 3,824 feet) located 11 km North East of Entebbe, Uganda. It is located at the northern crescent of Lake Victoria. It is a field station owned by the Government of Uganda under the Uganda Virus Research Institute (UVRI). Zika forest was originally described as a U-shaped forest covering an estimated 19.2 hectares (Fig. 1A; Buxton, 1952). Zika forest reduced to ∼12 hectares of inverted L-shaped forest (Birungi, 1995; Dranzoa, 1990). The change in shape and size of the forest is partly due to anthropogenic activities visible around the forest (Fig. 1B, C).
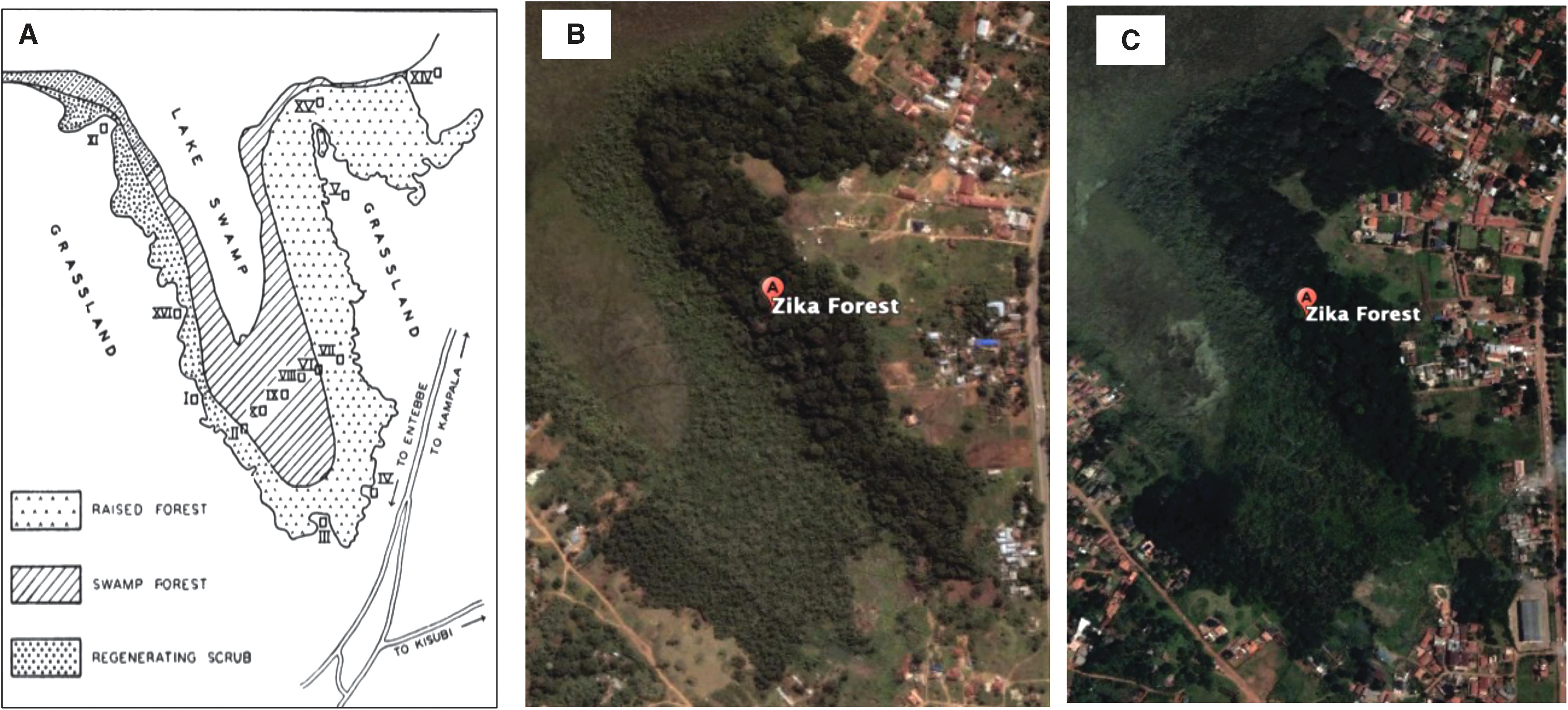
The strip of evergreen moist vegetation is bounded on one side by a papyrus swamp and open grassland on the other. The buffer zone is an influx of increasing anthropogenic activities despite the standing local legal instruments like the Uganda National Forestry and Tree planting act of 2003 (Government of Uganda, 2003) that prevent such activities in forest buffer zones in Uganda.
There is a steel tower 36.5 meters high in Zika forest, placed about 60 meters from the forest edge and ∼120 meters from the swamp. Since 1961, this tower has been used for entomological research, making several important contributions to the field of entomology (Corbet, 1964; Goma, 1965; Haddow et al., 1968).
Mosquito collection and handling
Trimmed bamboo pots measuring an average height of 20 cm, containing water at a height of about 17 cm were used to collect mosquito eggs, larvae, and pupae. Bamboo pots are suitable oviposition sites for tree hole breeding mosquitoes and have been used for mosquito surveys especially in the tropics (Corbet, 1964; Lounibos, 1981; Sempala, 1983). Two sampling designs were used for collecting immature samples inside and outside the forest. Inside the forest, the bamboo pots were placed on the steel tower at ground level and at platforms 6.1 meters (20 feet), 12.2 meters (40 feet), 18.3 meters (60 feet), 24.4 meters (80 feet), and 30.5 meters (100 feet) along the tower. Four bamboo pots were placed at each platform making a total of 24 pots during each collection.
Outside the forest, bamboo pots were placed at the ground level at the periphery of the forest (marginal forest), and in the buffer zone, (open grassland), at distances 5, 60, 120, and 125 meters from the edge of the forest. Four bamboo pots were placed at each of the distance points making a total of 16 pots during each collection. In the buffer zone, collection pots were fixed close to shrubs to avoid being tampered by grazing animals and humans. The pots were topped up with tap water every week after each collection.
Monthly collections of immature samples were conducted inside Zika forest and its buffer zone between July 2011 and July 2013. During this period, a total of 1,392 and 2,088 bamboo pots were exposed for sampling outside and inside Zika forest, respectively.
Collections of immature forms were made weekly from August 2011 to May 2013. During each collection, the immature mosquito forms were emptied into labeled disposable plastic cups. The pots were rinsed twice with tap water and contents were poured into the respective cups, to remove any remaining immature samples. They were then refilled with water and refixed firmly to the collection spots. The plastic cups containing immature forms were placed in secure containers and transported to an insectary at UVRI.
In the insectary, samples were sorted to remove debris and predatory Toxornychites species, counted and morphologically identified to the genus level. The plastic cups were then covered with a net to avoid the escape of emerging adults. The emerging adults were subsequently morphologically identified to species level using standard mosquito identification keys (Edwards, 1941; Gillett, 1972; Hopkins, 1936; Huang, 1990).
Two statistical indices were used in analyzing the abundance of each species: Percentage occurrence (PO), which is the percentage of pots with larvae to the total number of pots exposed (pots at risk); PO = no. of positive pots/total pots at risk × 100 Larvae density (LD), which is the average number of larvae collected from the pots at risk over the study period; LD = total larvae collected/number of pots at risk.
A χ2 test was applied to infer significance in the PO of larvae collected both inside and outside Zika forest. Analysis was done using the R software (R Project, 2013). The study was approved by the UVRI Research Ethics Committee (approval reference: GC/127/223).
Results
A total of 22,280 and 6,142 mosquito larvae and pupae were collected inside and outside Zika forest, respectively (Table 1). The mosquitoes belonged to five species: Aedes (Stegomyia) africanus (Theobald), Aedes (Stegomyia) aegypti (Theobald), Aedes arpicoargenteus, Aedes cumminsii, Aedes simpsoni s.l and Toxorynchites spp.
Total Counts and Larval Densities of Arboviral Vectors Collected Inside and Outside Zika Forest over the Study Period
Larval density is calculated as a ratio of total larvae collected and number of pots exposed (2,088 pots) over the study period for each species.
Inside Zika forest, Aedes apicoargenteus was the most dominant species, with up to 17,738 specimens and a larval density of 8.5 larvae/pot (Table 1). A. africanus was the second most abundant species (n = 3,277; Table 1). Both A. aegypti and A. cumminsii maintained dismal numbers and larval densities inside Zika forest (Table 1). Unexpectedly, a few A. aegypti mosquitoes were collected inside the forest (Table 1). Along the 36.5-meter steel tower in Zika, we observed a significant difference in the POs for most mosquito species with the highest POs observed at the canopy level and below for all the species (Table 2).
Number of Positive Pots for each Arboviral Vector Collected at Various Vertical Heights Along the Zika Tower Inside the Forest
A total of 348 pots were exposed for each level/platform over the study period. Percentage occurrence (in parentheses) is calculated as a percentage of positive pots to the total pots exposed for each species. Significance of differences is assessed with a χ2 test. Significant p-values are in bold.
Outside Zika forest, A. africanus and A. aegypti were the most dominant species with total counts of 2,963 and 2,764, respectively (Table 1). The PO of A. aegypti gradually increased from the marginal forest to distances that are closer to human settlements (60–125 meters from the forest margin) (Table 3). While A. apicoargenteus was abundant inside the forest (Table 1), it had a dismal presence both at the forest margin (PO = 1.15%) and the open grassland (PO = 2.01%, 1.15%, 0.86% at 60, 120, and 125 m, respectively) (Table 3). A. simpsoni, another important vector, was exclusively found outside the forest and in low numbers and larval densities (Table 1). On the other hand, A. cumminsii, like A. apicoargenteus, had its abundance gradually reducing from inside Zika forest to the forest buffer zone (Table 1).
Number of Positive Pots and the Percentage Occurrence (in Parentheses) for each Species for Collections Done at Various Distance Points Outside Zika Forest
A total of 348 pots were exposed for each distance point over the study period. Percentage occurrence is expressed as a percentage of positive pots to exposed points for each species. The significance of differences was calculated, and significant p-values are in bold.
Discussion
Zika forest is a habitat of several medically important mosquito vectors, including arboviral vectors identified in this study, that is, A. africanus, A. aegypti, A. simpsoni, A. cumminsii, and A. apicoargenteus (Corbet, 1964; Haddow et al., 1968; Haddow et al., 1964; Kaddumukasa et al., 2014). Many forests in Uganda, including Zika forest, have remarkably reduced in structure as measured by the Above Ground Biomass index due to continuous disturbance (Bulafu et al., 2013) caused by anthropogenic activities, such as deforestation, agricultural practices, and construction of human settlements. Figure 1 clearly demonstrates that Zika forest has over the years shrunk in size from a U to L shape (a reduction of more than 7 hectares), with increased human developments around the forest (Fig. 1B, C) and reported reduction in species biodiversity (Birungi, 1995; Dranzoa, 1990).
The current study has observed high abundances of A. africanus both inside and outside Zika forest. A. africanus is one of the most important arboviral vectors with up to 21 arboviral strains isolated from field specimens, including yellow fever virus, dengue virus, chikungunya virus, Zika virus, Rift Valley fever virus, and West Nile fever virus among others (Haddow, 1960; Haddow et al., 1964). A. africanus has played a role in recent emerging and re-emerging arboviral disease outbreaks in Africa. For example, A. africanus was found to be relatively abundant in the 2006 chikungunya outbreak in Cameroun and probably played a role in the transmission of chikungunya virus (Demanou et al., 2010). It was also the principal vector during the major yellow fever epidemics in Nigeria in 1987 and Kenya in 1992–1993 (De Cock et al., 1988; Ellis and Barrett, 2008; Reiter et al., 1998).
Entomological studies in several forests in Africa and those conducted in Ugandan forests following the yellow fever outbreak of 1941 indicated that A. africanus was mainly a forest-dwelling mosquito (Ayres, 2016; Benelli and Romano, 2017; Guindo-Coulibaly et al., 2019; Huang, 1990). However, Corbet (1964) working in Zika forest found that A. africanus was not only found in the forest but also at the periphery and buffer zone (Corbet, 1964). Our study concurs with Corbet (1964) in the distribution of A. africanus however, it is worth noting that during Corbet's study, there were no human settlements close to the forest. The increasing anthropogenic activities currently observed in Zika forest and the subsequent land cover changes are a matter of concern as they do influence the microclimatic conditions of mosquito habitats and subsequently the ecology and/or behavior of these vectors (Afrane et al., 2012; Kweka et al., 2016; Munga et al., 2009). With humans invading the vector habitats and conversely the vectors finding refuge in areas of human dwellings, the likely result is the enhancement of arboviral disease transmission between man and other nonhuman primates.
Of concern, is a recent study that observed human blood meal in engorged A. africanus samples collected from within Zika forest (Kaddumukasa et al., 2015), an indication that these mosquitoes are actively biting humans who are either encroaching on the forest or are within its buffer zone. Similar land use and land cover changes have reportedly influenced the emergence and reemergence of malaria epidemics in East Africa (Githeko et al., 2000; Lindblade et al., 2000; Lindblade et al., 1999; Paul et al., 2018).
Furthermore, the ability of A. africanus to oviposit, close to human settlements (as shown in the current study) is typical of a peridomestic form of behavior. Peridomestic forms of A. africanus have been reported in South Eastern Nigeria, where they breed in artificial containers like clay pots, ovipositing as frequently in houses as outdoors (Bang et al., 1980; Omar et al., 2021). The land use and land cover changes due to anthropogenic activities in Zika forest could potentially alter the behavior of A. africanus, which, according to this study, can acquire a peridomestic behavioral form. Interestingly, a similar trend toward peridomesticity has been observed among sylvatic populations of A. africanus near an urban setting in Abidjan, Ivory Coast (Guindo-Coulibaly et al., 2019). Indeed, land cover changes (like deforestation) reportedly increased the vectorial capacity of the malaria vector, Anopheles gambiae, subsequently increasing malaria epidemics in the western Kenyan highlands (Afrane et al., 2008). The observed likely change in behavior in A. africanus could therefore alter the disease transmission dynamics leading to the emergence/reemergence of arboviral epidemics.
Our study has also observed that the vertical height oviposition preference for A. africanus is at the ground level, although there are substantial collections at the tree canopy level. A similar observation was made by Corbet (1964). This is probably not surprising given that A. africanus is predominantly involved in the sylvatic cycle of arboviral transmission (Haddow et al., 1948; Huang, 1990; Mattingly, 1952). It feeds on primates (monkeys) in the forest canopy and oviposits in natural sites, for example, tree holes and stump holes in forests (Huang, 1990).
A. aegypti is another arboviral vector collected from Zika forest in this study. It is a primary vector to humans of the four serotypes of dengue viruses, yellow fever virus (Gubler, 2019; Lwande et al., 2020), and is a known vector of chikungunya virus (Russo et al., 2020). It was recently implicated as the main vector in the chikungunya outbreak in 2016 in Kenya (Konongoi et al., 2018) and it also maintained endemic transmission of dengue and chikungunya in Mozambique (Mugabe et al., 2018).
During previous studies by Corbet (1964) and Haddow et al. (1964) in Zika forest, immature forms or adult A. aegypti were never collected inside the forest. Our data indicate the contrary with a change in the pattern of distribution of A. aegypti, being collected both inside and outside the forest. Therefore, A. aegypti inside Zika forest was an unexpected finding, which is also corroborated in a later adult mosquito sampling effort within Zika by Kaddumukasa et al. (2014). This suggests that ecological conditions or the local climate has become conducive for the proliferation of A. aegypti in the forest. Indeed, the forest canopy has reduced with the increasing human activities involving tree felling in the forest. The presence of A. aegypti, commonly involved in the urban cycle of yellow fever transmission in association with A. africanus (which is mainly involved in the sylvatic cycle of arboviruses), both inside and outside the forest (Table 1 and Fig. 2) is likely to facilitate the spread of arboviruses among communities now living in proximity to the forest. Incidentally, both A. africanus and A. aegypti were vectors in the yellow fever outbreak in Senegal in 2020 (Diagne et al., 2021).
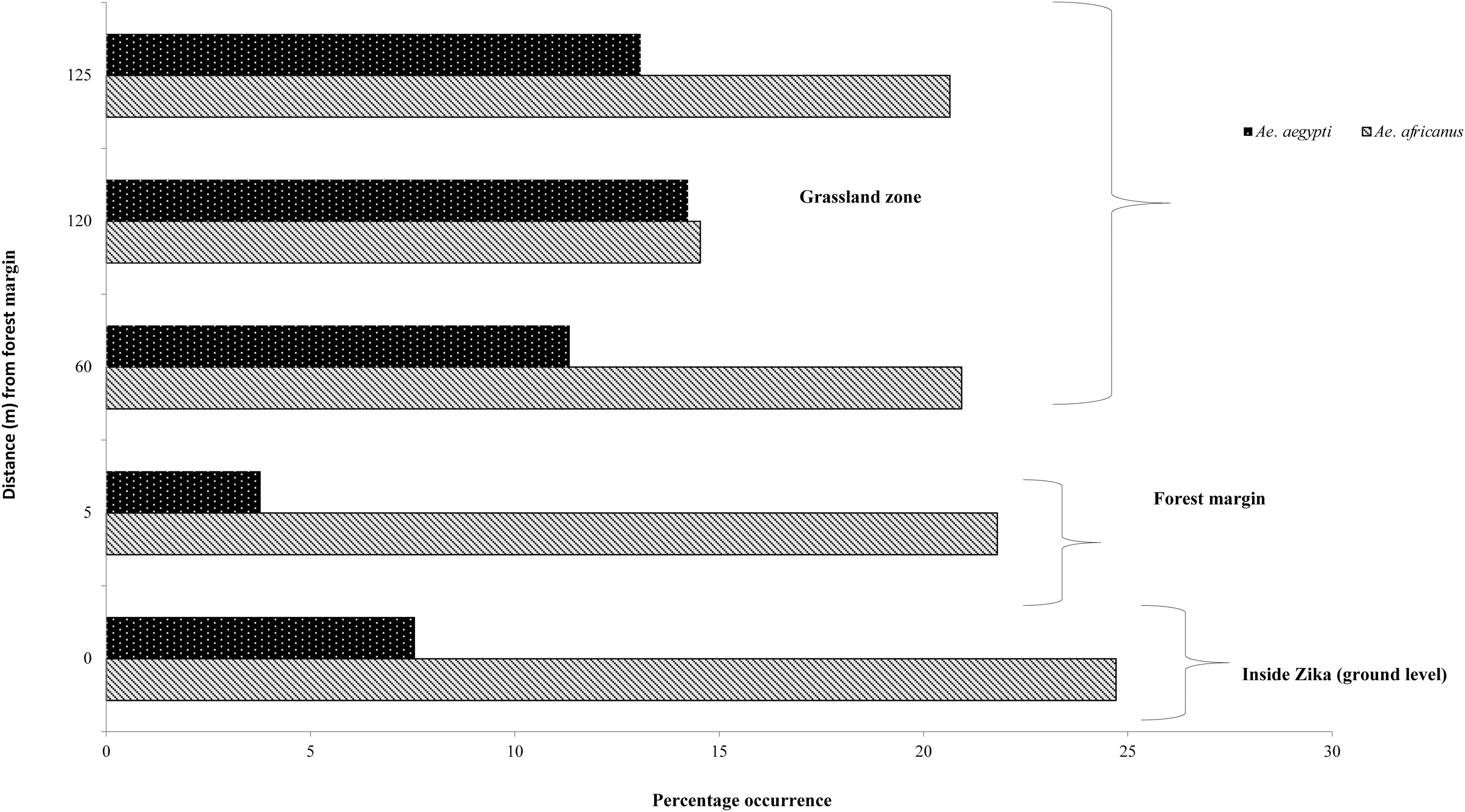
Percentage occurrence of Aedes africanus and Aedes aegypti from the ground level of Zika, through marginal forest and grassland zones.
In Uganda, a high vector competence of A. aegypti in the transmission of Zika virus was reported in a study conducted in Entebbe, Uganda (Mutebi and Lutwama, 2021).
Another vector A. cumminsii is considered an important arboviral vector having isolated 10 viruses, including chikungunya virus, dengue virus, Rift Valley fever virus, and Spondweni virus (Berge, 1975). Previous work in Zika forest indicated that, A. cumminsii adults were caught only inside the forest (Haddow et al., 1964). However, Corbet (1964) working on the oviposition activity in the forest did not record any A. cumminsii collections (Corbet, 1964). Our data, however, show that A. cumminsii oviposited both inside and outside the forest. This is yet another vector that has expanded its range of distribution in the forest.
Other important arboviral vectors, A. simpsoni s.l and A. apicoargenteus, have maintained their distribution over the years (Corbet, 1964; Haddow et al., 1964). A. simpsoni, an important yellow fever vector, oviposits outside the forest both in bamboo and axils of Dracaena ugandensis (plants commonly found in agricultural areas around the forest buffer zone) (Haddow et al., 1948; Pajot, 1983). Aedes bromeliae one of the species in the A. simpsoni s.l complex is a known vector of yellow fever in East Africa (Huang, 1986). Aedes lilii is the nonanthropophilic subspecies within the complex. A. apicoargenteus, a vector of the emerging pathogen, Zika virus and re-emerging arbovirus chikungunya, was frequently found inside and marginally outside the forest (Hayes, 2009; Sempala and Kirya, 1973).
The behavioral changes in the vectorial capacity as reported in A. aegypti, compounded with the changing vector distribution of identified arboviral vectors in and out of Zika forest poses an increased concern in the disease dynamics, prevention, and control of the arboviral diseases. More disease surveillance studies in mosquito vectors, humans, and primates are however required to inform the prevention of disease pathogens that the vectors transmit.
The findings in this study have important implications on (i) the emergence and reemergence of arboviral diseases, and (ii) the management of arboviral outbreaks/epidemics given that the yellow fever virus along with other important arboviruses have been isolated from mosquito vectors, including A. africanus, in Zika forest (Haddow et al., 1964; Kirya and Okia, 1977; Kirya et al., 1977). A more current arboviral surveillance of both mosquitoes and primates in the forest is however required to determine the circulating arboviruses in the forest and its buffer zone to reliably advise on prevention and control strategies especially given the recent reemergence of yellow fever outbreaks in Uganda (Kwagonza et al., 2018; McMullan et al., 2012; Wamala et al., 2012).
Conclusion
In summary, environmental changes due to deforestation and anthropogenic activities probably have had an impact on the distribution and behavior of arboviral vectors in Zika forest. These changes in the microclimate have potentially enhanced the proliferation of some vectors to areas where they previously did not occur. This has led to the association of two major arboviral vectors, A. aegypti and A. africanus, potentially influencing arboviral transmission in the area. These findings guide the appropriate public health management in the area and other regions where forest encroachment has occurred. Noteworthy is that if these findings are extrapolated to the whole country where land use and land cover changes have increased over the years, it is probably no wonder that a yellow fever outbreak did occur in Uganda after over 40 years (Kwagonza et al., 2018; McMullan et al., 2012; Wamala et al., 2012).
So far, sensitization campaigns have been conducted in the area discouraging further anthropogenic activities around the forest to prevent possible transmission of arboviruses in the community. Additional studies to understand the vectorial capacity and predict the risk of arboviral outbreaks/epidemics are required, including (i) the breeding and biting behavior of arboviral vectors, (both indoor and outdoor) (ii) the seasonal population dynamics in relation to rainfall and temperature and their effects on the ecology, and (iii) surveillance of arboviruses in mosquito vectors, the potential reservoir hosts (primates, birds, reptiles) and the human host. Ultimately vaccination against yellow fever is encouraged and for the other arboviruses, all of which have no safe and effective vaccines developed yet, efforts should be made to minimize human–vector contact.
Data Availability Statement
Data are available from the corresponding author upon request.
Footnotes
Acknowledgment
The authors thank the staff of the Division of Entomology (Uganda Virus Research Institute) for their support during the field collections and rearing in the insectary.
Authors' Contributions
Study conceptualization by L.G.M., J.B., and C.M. Field data collection, laboratory work and analysis by M.L. and J.B. Article draft by M.L. and J.B. All authors have reviewed and approved the final article.
Author Disclosure Statement
The authors declare that they have no competing interests.
Funding Information
This work was supported by The Presidential Support to Scientists, an initiative of the President of Uganda to Dr. L.G.M.; International Association of Public Health Institutes (IANPHI) subaward No. 6-38223-G1 to Dr. J.B., and Training Health Researchers into Vocational Excellence (THRiVE) in East Africa, Grant number 087540, funded by Wellcome Trust to Dr. M.L.