
Abstract
Background:
Clonorchiasis remains a serious public health problem. However, the molecular mechanism underlying clonorchiasis remains largely unknown. Amino acid (AA) metabolism plays key roles in protein synthesis and energy sources, and improves immunity in pathological conditions. Therefore, this study aimed to explore the AA profiles of spleen in clonorchiasis and speculate the interaction between the host and parasite.
Methods:
Here targeted ultrahigh performance liquid chromatography multiple reaction monitoring mass spectrometry was applied to discover the AA profiles in spleen of rats infected with Clonorchis sinensis. Kyoto Encyclopedia of Genes and Genomes pathway enrichment analysis (KEGG) was performed to characterize the dysregulated metabolic pathways.
Results:
Pathway analysis revealed that phenylalanine, tyrosine, and tryptophan biosynthesis and β-alanine metabolism were significantly altered in clonorchiasis. There were no significant correlations between 14 significant differential AAs and interleukin (IL)-1β. Although arginine, asparagine, histidine, lysine, methionine, phenylalanine, proline, serine, threonine, tryptophan, tyrosine, and valine were positively correlated with IL-6, IL-10, tumor necrosis factor (TNF)-α as well as aspartate aminotransferase and alanine aminotransferase; β-alanine and 4-hydroxyproline were negatively correlated with IL-6, IL-10, and TNF-α.
Conclusion:
This study reveals the dysregulation of AA metabolism in clonorchiasis and provides a useful insight of metabolic mechanisms at the molecular level.
Introduction
C
Metabolomics provides insights into underlying reactions toward genetic and environmental perturbations in a biological system (Schrimpe-Rutledge et al., 2016). Targeted metabolomics is the absolute quantification of chemically characterized and biochemically annotated metabolites (Roberts et al., 2012). Metabolomics has been widely used in exploring pathogenic mechanisms, screening potential biomarkers and early diagnosis of parasitic disease (Wu et al., 2010; Yue et al., 2020). Thus, profiles of metabolism could be contributed to understand the pathogenesis of clonorchiasis.
Metabolites included amino acids (AAs), nucleic acids, lipids, and other products. Because AAs are required for protein synthesis and energy sources in all living cells (Kelly and Pearce, 2020), AAs play a critical role in organ protection as well as energy metabolism (Chen et al., 2018). Previous studies have shown that AA metabolism could affect the immune response, inflammation, and disruption of the tricarboxylic acid (TCA) cycle (Nguyen et al., 2018). The production of the profibrotic cytokines such as TGF-β and monocyte chemoattractant protein 1 (MCP-1) has been identified to be associated with AA metabolism and TCA cycle (Gong et al., 2019; Li et al., 2017). Multiple mechanisms could sense AAs to control immune cell metabolism (Kelly and Pearce, 2020). Alterations of AA metabolism could reflect the impact of infection on metabolism, but also reveal host immune response to infection and/or tissue injury and repair (Chandler et al., 2016; Kato et al., 2018).
Abnormal AA metabolism was involved in the pathogenesis and progression of many diseases. A previous study showed that serine, glycine, alanine, and histidine were significantly higher in C. sinensis–infected bile (Li et al., 2020), which serve as important energy source for C. sinensis (Li et al., 2020). Although we identified some endogenous metabolites in clonorchiasis, little is known about the mechanisms involved in the host–pathogen interaction, especially AAs metabolic profiling and related metabolic pathway. Therefore, the AA profiles of spleen in C. sinensis–infected rats were investigated by a targeted ultrahigh performance liquid chromatography multiple reaction monitoring mass spectrometry (UHPLC-MRM-MS/MS). The relationships among the AAs and some inflammatory factors as well as progression (aspartate aminotransferase [AST] and alanine aminotransferase [ALT]) were further analyzed. The exploration of AAs metabolism and related biochemical pathways could be useful for enhancing our understanding of the immunoregulatory mechanisms in clonorchiasis.
Materials and Methods
Animal infection
C. sinensis metacercariae were collected from Pseudorasbora parva in the Songhuajiang River of Heilongjiang Province. The collection and preparation of metacercariae were as follows. The fish was washed with tap water, broken up in a Waring Blender, and then digested with a pepsin-HCl (0.6%) artificial gastric juice at 37°C for 12 h. The digested mixture was passed through three sieves with mesh sizes of 1000, 300, and 106 μm in sequence. Finally, metacercariae were harvested by centrifugation and stored at 0.1 M phosphate-buffered saline (pH = 7.4) at 4°C until use. Male Wistar rats (5–6 weeks old) were purchased from the Harbin Medical University Laboratory Animal Center. Rats were randomly divided into two groups: infected group (n = 16) and control group (n = 16). In the infected group, rat was infected orally with 50 metacercariae, and control rat was fed with 50 μL of sterile normal solution. All rats were observed daily throughout the experiment.
Tissue collection and detection of infection
Feces were collected weekly and microscopically examined by the Kato-Katz method to determine whether the rats had been successfully infected with C. sinensis (Hong et al., 2003). C. sinensis adults develop matured, start producing eggs, and cause obvious inflammatory cell infiltration, fibrocyte accumulation, and serious collagen deposition from 4 weeks postinfection (wpi) (Lun et al., 2005; Xu et al., 2016). Our study found hepatocyte apoptosis index increased from 4 wpi and reached a peak at 8 wpi (Zhang et al., 2008), hepatic injury and serum metabolimics perturbations in 4 wpi (Han et al., 2023), as well as liver iron deposits apparently at 8 wpi (Han et al., 2017). Similarly, some research also selected 4 and 8 wpi to explore the pathogenic mechanism of clonorchiasis (Lee et al., 1987; Lun et al., 2005; Uddin et al., 2012; Xu et al., 2016; Yan et al., 2015; Zhang et al., 2008).
Thus, based on these significant characteristics of the worm growth, pathologic changes, and disease progress, rats were killed at 4 and 8 wpi for analysis. The spleens were rinsed with saline solution (0.9% sodium chloride w/v), and stored at −80°C until analysis. Control rats were killed at both time points.
Metabolites extraction
First, each spleen sample was weighed and transferred to an Eppendorf tube. Then two little steel balls and 1000 μL of extract solvent (precooled at −20°C, acetonitrile–methanol–water, 2:2:1, containing isotopically labeled internal standard mixture) were added in the tube. Second, after vortexed (30 s), the samples were homogenized (45 Hz, 4 min), sonicated (5 min, ice water bath), and repeated three times. Then the mixture was incubation at −40°C for 1 h and centrifugation at 12,000 rpm, 4°C for 15 min. Finally, the supernatant was transferred into an autosampler vial for UHPLC-MS/MS analysis.
Standard solution preparation
After dissolving or diluting each standard substance, it was transferred to a 10 mL flask to mix working standard solution. A series of calibration standard solutions were prepared by stepwise dilution of this mixed standard solution (containing isotopically labeled internal standard mixture in identical concentrations with the samples).
UHPLC-MRM-MS analysis
The LC-MS/MS analyses were performed in Shanghai Biotree Biotech Co., Ltd. The UHPLC separation was running an Agilent 1290 Infinity II series UHPLC System (Agilent Technologies), equipped with a Waters ACQUITY UPLC BEH Amide column (100 × 2.1 mm, 1.7 μm). The injection volume was 1 μL. The mobile phase A and B were 1% formic acid in water and acetonitrile, with the column temperature (35°C) and autosampler temperature (4°C), respectively. An Agilent 6460 triple quadrupole mass spectrometer (Agilent Technologies), equipped with an AJS electrospray ionization interface, was used for assay development. Typical ion source parameters were as follows: capillary voltage = +4000/−3500 V, nozzle voltage = +500/−500 V, temperature = 300°C, flow = 5 L/min, sheath temperature = 250°C, sheath gas flow = 11 L/min, nebulizer = 45 psi. Agilent MassHunter Work Station Software (B.08.00; Agilent Technologies) was used for MRM data acquisition and processing.
Calibration curves
Calibration solutions were subjected to UPLC-MRM-MS/MS analysis. Least squares method was used for the regression fitting. 1/x weighting was applied in the curve fitting because it provided highest accuracy and correlation coefficient (R 2). The level was excluded from the calibration if the accuracy of calibration was not within 80–120%. The detailed calibration curves, limit of detection (LOD), and limit of quantitation (LOQ) for individual analyses are given in Supplementary Fig. S1 and Supplementary Table S1.
The calibration standard solution was diluted stepwise, with a dilution factor of 2. These standard solutions were subjected to UHPLC-MRM-MS analysis. The signal-to-noise (S/N) ratios were used to determine the lower LODs (LLODs) and lower LOQs (LLOQs). The LLODs and LLOQs were defined as the analyzed concentrations that led to peaks with S/N ratios of 3 and 10, respectively, according to the U.S. FDA guideline for bioanalytical method validation.
Precision and accuracy
The precision of the quantitation was measured as the relative standard deviation, determined by injecting analytical replicates of a quality control (QC) sample. The accuracy of quantitation was measured as the analytical recovery of the QC sample determined. The percent recovery was calculated as [(mean observed concentration)/(spiked concentration)] × 100%.
Detection of inflammatory factors, ALT and AST in serum
The level of inflammatory factors was detected by Luminex 200 liquid suspension microarray and Bio-Plex Pro Rat Cytokine 1 10PLX EXP Kit. Samples were processed following manufacturer's instructions. The experimental procedure is briefly described as follows: 50 μL diluted mixed micro-beads was added into wells, then 50 μL of standard products and samples were added corresponding to each well (diluted twice with assay buffer in advance). The plate was incubated with shaking at room temperature for 60 min and washed three times. After that, it was reincubated with 25 μL biotin-labeled detection antibody for 30 min and washed three times. After washing, the samples were reincubated in 50 μL diluted temperature for 10 min at room temperature. Following another wash, the magnetic beads were resuspended with 125 μL assay buffer. Each sample was measured in duplicate. Data were analyzed and calculated with GraphPad Prism 4 software (San Diego, CA).
Correlation analysis among AAs, inflammatory factors and progression
To investigate the relationship between metabolites and inflammatory factors as well as progression (ALT and AST), Spearman coefficients were calculated between spleen AAs and serum factors (interleukin [IL]-1β, IL-6, IL-10, TNF-α, ALT, and AST). Correlation coefficients (r) greater than 0.500 or less than −0.500 were deemed moderately strong correlations. Clustering correlation heatmap with signs was performed using the OmicStudio tools at www.omicstudio.cntodisplaythestrengthofallmetabolitescorrelations
Statistical analysis
Data were presented as mean ± SD. SPSS 13.0 software (SPSS, Inc., Chicago, IL) was used for statistical analysis. Continuous variables were analyzed by Student's t-test. Value of p < 0.05 was considered significant.
The AAs data set containing the information of peak number, sample name, and normalized peak area was imported to SIMCA 16.0.2 software package (Sartorius Stedim Data Analytics AB, Umea, Sweden) for multivariate analysis. The principle component analysis (PCA), an unsupervised analysis that reduces the dimension of the data, was used to visualize the distribution and the grouping of the samples. The orthogonal projections to latent structures-discriminant analysis (OPLS-DA), a supervised multivariate data analysis tool, was applied to evaluate the variation of metabolites at different infection groups. The Variable Importance in the Projection (VIP) of OPLS-DA model was >1, so as to identify the metabolites expressed differently. Log2 fold change was used to assess the variation of the metabolites. Volcano plots and heatmaps were applied to describe the unbalanced metabolic profiles among C. sinensis–infected and control rats.
The corresponding Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways were matched to each metabolite in KEGG online (http://geneontology.org/). Further analysis on the pathway affected by C. sinensis infection was used by MetaboAnalyst 3.0 (www.MetaboAnalyst.ca/).
Ethics statement
The animal research protocol has been reviewed and assigned by the Medical Ethics Review Committee of Harbin Medical University that approved our animal experiments (Animal Use License SYXK-2019-001). All animal experiments were carried out based on the Guide for the Care and Use of Laboratory Animals published by the Ministry of Science and Technology of the People's Republic of China. In the study, we made great efforts to reduce animal suffering and the number of animals.
Results
AA metabolite profiles
A total of 25 AAs were established and further evaluated in the spleen by the targeted UHPLC-MRM-MS/MS quantitative analysis. The quantitative results of AA profiles are given in Supplementary Table S2. Then PCA was performed to analyze and visualize the AA profiles in all samples (Fig. 1). In addition, the two components of the OPLS-DA explained 80.7% and 93.7% (R2X = 0.807, R2Y = 0.937) of the total variances and predicted 90% of the variance (Q2 = 0.9) at 4 wpi (Fig. 2 and Supplementary Table S3). The samples of different infection groups and control groups were clearly separated in the OPLS-DA models, which indicated the excellent discriminatory power in spleen metabolites. The permutation test demonstrated the robustness of the OPLS-DA model (Fig. 2). The volcano plot of AAs metabolites is given in Fig. 3. A clear clustering and difference were illustrated between different time-specific metabolic phenotypes by heatmap analysis (Fig. 3). These results indicated that significant AA metabolic alterations occurred in clonorchiasis.
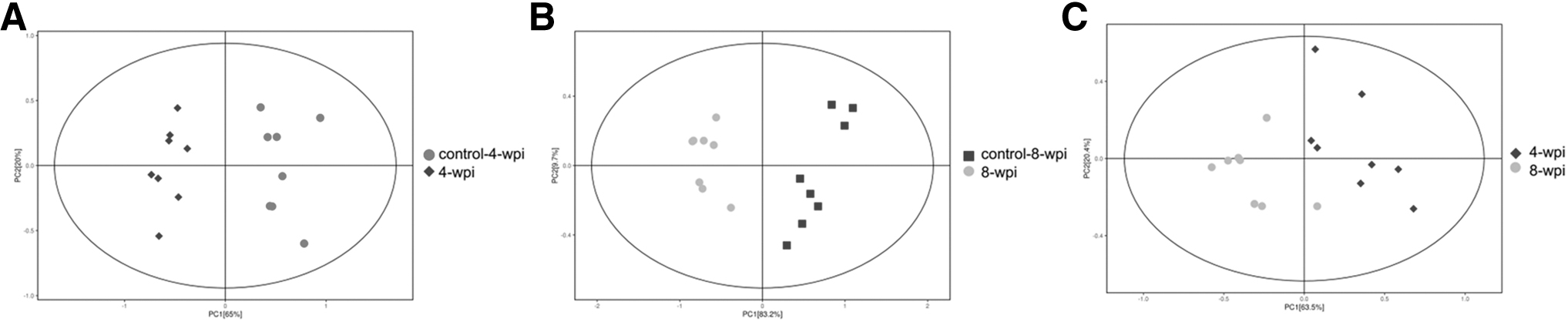
Principal component analysis score scatter plots of amino acids metabolites in spleen during Clonorchis sinensis infection.

Multivariate statistical analysis of AAs metabolites in spleen during C. sinensis infection. OPLS-DA score plots of AAs metabolites in 4 wpi vs. control

Differential AAs metabolites were identified among different groups during C. sinensis infection. The volcano plot displays the differentially expressed AA metabolites in 4 wpi vs. control
Potential important metabolites
Based on VIP >1 and p < 0.05, a total of 14 AAs were used for screening the potential biomarkers at both 4 and 8 wpi. Among them, the concentration of arginine, asparagine, histidine, lysine, methionine, phenylalanine, proline, serine, threonine, tryptophan, tyrosine, and valine were significantly upregulated, whereas β-alanine and 4-hydroxyproline were found to be dramatically downregulated (Table 1).
Identification of the Potential Amino Acid Biomarkers Discovered by Variable Importance in the Projection Values in Clonorchiasis
FC, fold change; Q, Q-values were p-values adjusted by false discovery rate method across the metabolites within each comparison; VIP, variable importance in the projection based on orthogonal projections to latent structures-discriminant analysis model; wpi, weeks postinfection.
AAs metabolic networks
The exploration of biological pathways could be helpful in clarifying the mechanisms of clonorchiasis. Based on the value of p < 0.05 and pathway impact value, some significant differential AA metabolic pathways involved in clonorchiasis were estimated by KEGG annotation and MetaboAnalyst. As given in the bubble chart (Fig. 4), phenylalanine, tyrosine, and tryptophan biosynthesis, β-alanine metabolism, arginine biosynthesis, pantothenate and coenzyme A (CoA) biosynthesis, phenylalanine metabolism and arginine and proline metabolism were enriched at both 4 and 8 wpi (Supplementary Table S4). Our results suggested that there were significantly changed metabolic biological networks (Supplementary Fig. S2).

Pathway analysis of the identified differential AAs metabolites for 4 wpi vs. control
Correlation analysis between AAs and inflammatory factors
IL-6, IL-10, and TNF-α were found to significantly increase at 4 and 8 wpi (p < 0.05), although there was no significant change in IL-1β both at 4 and 8 wpi (p > 0.05) (Fig. 5). The correlation heatmap showed no significant correlations between 14 AAs and IL-1β, whereas arginine, asparagine, histidine, lysine, methionine, phenylalanine, proline, serine, threonine, tryptophan, tyrosine, and valine were positively correlated with IL-6, IL-10, and TNF-α, β-alanine and 4-hydroxyproline were negatively correlated with IL-6, IL-10, and TNF-α (Fig. 5E).
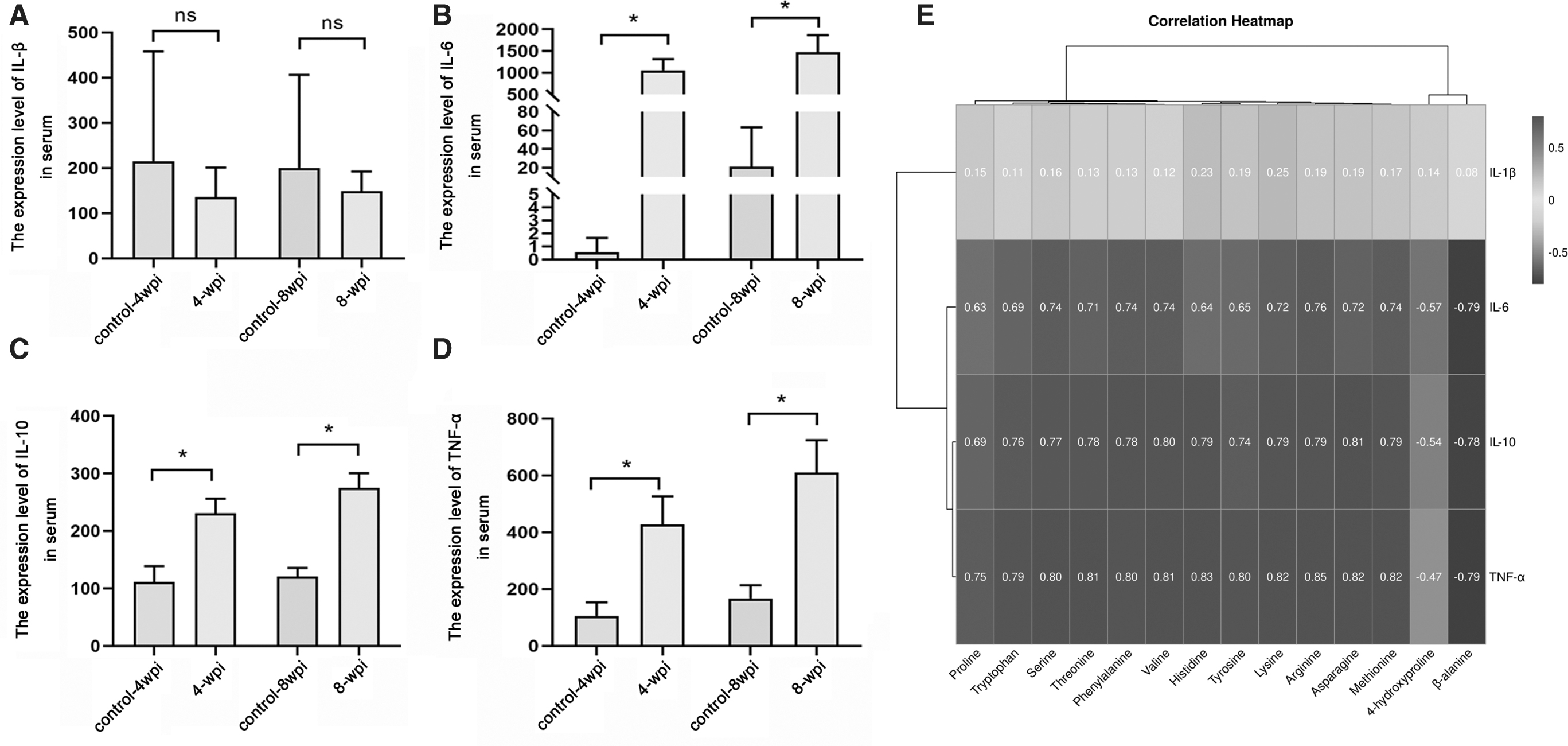
Association of AAs level and inflammatory factors. The serum levels of IL-1β
Correlation analysis among AAs, AST, and ALT
AST and ALT were found significantly increase at 4 and 8 wpi, respectively (p < 0.05) (Fig. 6). We analyzed correlation between progression (AST and ALT) and significant differential AAs. Proline, serine, tyrosine, histidine, phenylalanine, threonine, tryptophan, asparagine, lysine, arginine, methionine, and valine were positively correlated with AST and ALT, respectively, whereas 4-aminobutyric acid and 4-hydroxyproline were negatively correlated with AST and ALT, respectively (Fig. 6).

Association of AA level and aminotransferase (ALT and AST). The serum levels of ALT
Discussion
Previous studies on the interactions between C. sinensis and hosts have mainly focused on variation in protein or genes of the liver (Han et al., 2021; Han et al., 2016). Except for hepatic injury, C. sinensis infection also causes remarkable immune activation of cell-mediated immunity and humoral immunity (Na et al., 2020). The parasite utilizes kinds of immunoregulatory mechanisms to fight host defenses, including dysregulation of the expression of cytokines, alteration in the architecture of spleen, and developing antiparasitic humoral immune response (Xu et al., 2016; Zheng et al., 2017). Spleen is a critical immune organ having an important role in the innate and adaptive immune responses (Chen et al., 2017). In addition, the white pulp possesses B cell and T cell zones, and develops specific immune responses to protect host against microbial infection (Chen et al., 2017). Therefore, in-depth understanding of the role of spleen could contribute to understand the pathogenesis of clonorchiasis.
It was reported that metabolites play important roles in cell differentiation, inflammation and tumorigenesis, and it is involved in host–pathogen interactions (Muxel et al., 2019; Nawaz et al., 2019; Wu et al., 2018). Of interest, the interaction between immunity and metabolism also plays a key role in the pathogenesis of clonorchiasis. C. sinensis infection can evoke significant serum metabolic reprogramming of host (Han et al., 2023). Meanwhile, immunoinflammation is a common feature associated with C. sinensis infection in the peripheral circulation (Sriraj et al., 2016). Therefore, because immunity is closely related to metabolism, as well as the importance of the spleen in the immune response, the metabolic profiles of spleen in C. sinensis–infected rats were investigated (Zhang et al., 2020). However, the literature did not carefully analyze the role of AA metabolism, and did not combine cytokines to analyze the correlation between AA metabolism and immunity. To investigate the spleen AA profiles and related metabolic pathways in clonorchiasis, a targeted metabolomic approach was applied.
In this study, most AA levels were found to be altered in C. sinensis–infected rats (Supplementary Table S2), indicating that clonorchiasis was accompanied by changes in AA metabolism. For example, the concentration of 4-hydroxyproline and β-alanine were decreased, suggesting the utilization of these two AAs. 4-Hydroxyproline plays crucial roles in protecting mammalian cells from oxidative stress and injury (Wu et al., 2019). Furthermore, 4-hydroxyproline could promote the immunological defense of humans against bacteria, fungi, parasites, and virus infection by enhancing the metabolism and functions of monocytes, macrophages, and other cells of the immune system (Wu, 2020). It was reported that host gain the required energy by converting 4-hydroxyproline to glycine during C. sinensis infection (Wu et al., 2019). Some functional effect of some identified amino acids on immune cells were present in Supplementary Table S5.
Of note, several AAs significantly increased during C. sinensis infection, such as histidine, valine, serine, and tryptophan. Histidine is an important AA, which is associated with protein energy, wasting, inflammation, and oxidative stress (Watanabe et al., 2008). Histidine supplementation could reduce the concentration of TNF-α (Nasri et al., 2020). TNF-α is a strong proinflammatory cytokine that plays an important role in the immune system during inflammation, cell proliferation, differentiation, and apoptosis (Son et al., 2005). In addition, the levels of proinflammatory cytokines such as TNF-α, IL-1β, and IL-6 were differentially upregulated in C. sinensis–infected sera (Maeng et al., 2016). Thus, we speculated that C. sinensis may influence TNF-α secretion by upregulating histidine expression.
The deficiency of valine could inhibit the differentiation of monocytes into mature dendritic cells (Chen et al., 2017; Kakazu et al., 2007), although the upregulated of valine suggests its potential immunomodulatory metabolite enhanced host immunity against infection (Moura et al., 2017). Serine is a mediator of macrophages activation, which elevates expression of genes IL-1β and IL-8 in the spleen and plays a role in innate immunity (Du et al., 2017). In addition, the change of tryptophan levels is associated with T cell function and apoptosis in HIV-infected patients (Ghannoum et al., 2013). Taken together, these AAs dysregulation could modulate host immunity against infections in clonorchiasis.
In comparison, 1-methyl-
In addition, research in the variations of biological pathways could further explore the pathogenesis of clonorchiasis. Compared with control groups, several metabolic perturbation pathways, such as phenylalanine, tyrosine, and tryptophan biosynthesis, β-alanine metabolism, arginine biosynthesis, pantothenate and CoA biosynthesis, phenylalanine metabolism, and arginine and proline metabolism were found disturbed. Further investigation on the mechanisms behind these abnormal metabolic pathways could be helpful for exploring the etiology and making prevention strategies for clonorchiasis.
This study showed the AA metabolism and some biological pathways are involved in clonorchiasis. However, there are some limits of this study. First, owing to the complex and dynamic cellular heterogeneity of the spleen, the altered metabolite may be the result of differential expression of spleen cell. In the future, an individual cell population in the spleen will be selected for metabolomics or multi-omics joint analysis. Second, the sample size was small, AA changes in spleen needed to be validated by large-scale samples and further estimated in human subjects. Finally, the functional effect of some identified AAs on immune cells (Supplementary Table S5) should be further identified and analyzed.
Conclusions
In conclusion, this study presents the spleen AA metabolic signature and several metabolic perturbation pathways in clonorchiasis. These findings not only deepen our understanding of metabolic alterations, but also offer new insights into the complex regulation of metabolism in clonorchiasis. Future analysis is needed to characterize the correlation between splenic metabolites and its immunoregulation during C. sinensis infection.
Footnotes
Authors' Contributions
J.W. and J.D.: Data curation; formal analysis; methodology; writing—original draft. X.Z.: Data curation; resources. X.H.: Formal analysis; methodology. R.C.: Data curation; resources. S.H.: Conceptualization; data curation; review and editing; funding acquisition; methodology; supervision.
Disclaimer
The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the article.
Author Disclosure Statement
The authors declare that they have no competing interests. All authors have given their permission to publish data presented in this article. All the data supporting our findings are contained within the article.
Funding Information
This work was supported by National Natural Science Foundation of China (81971958), Natural Science of Heilongjiang province (YQ2020H006).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.