
Abstract
Background:
The control and prevention of rodent-borne diseases are mainly based on our knowledge of ecology and the infectious status of their reservoir hosts. This study aimed to evaluate the prevalence of Francisella tularensis, Yersinia pestis, and arenavirus infections in small mammals and to assess the potential of disease occurrence in East Azerbaijan, northwest of Iran, in 2017 and 2018.
Methods:
Spleen and lung samples were obtained from all trapped small mammals. The real-time quantitative PCR (qPCR) method was used to detect nucleic acid sequences of F. tularensis, Y. pestis, and arenaviruses. Serum samples were tested for antibodies indicating the host response to F. tularensis and Y. pestis infections using the standard tube agglutination test and enzyme-linked immunosorbent assay (ELISA), respectively.
Results:
A total of 205 rodents, four Eulipotyphla, and one carnivore were captured. The most common rodent species captured (123 of 205 rodents, 60%) belonged to the genus Meriones (mainly Persian jird, Meriones persicus). In total, 317 fleas were removed from trapped animals. Flea species belonged to Xenopsylla buxtoni, Xenopsylla nuttalli, Stenoponia tripectinata, Paraceras melis, Ctenophthalmus rettigi smiti, Rhadinopsylla bivirgis, Paradoxopsyllus grenieri, and Nosopsyllus iranus. Using the qPCR tests, five spleen samples from M. persicus were positive for F. tularensis. The qPCR tests were negative for the detection of Y. pestis and arenaviruses. Finally, all serum samples tested were negative for antibodies against Y. pestis and F. tularensis.
Conclusions:
F. tularensis was the only zoonotic agent detected in rodents captured in East Azerbaijan. However, the diversity of trapped rodents and fleas provides the potential for the spread of various rodent-borne viral and bacterial diseases in the studied areas.
Introduction
The control and prevention of rodent-borne diseases largely depend on our understanding of the ecology and infection status of these animals. Climatic and ecological fluctuations are associated with the demographic dynamics of rodent populations, changing the route of pathogen transmission, and potentially increasing the risk of their transmission to humans (Mills et al., 1999). Therefore, it is of crucial importance to periodically monitor reservoir populations and their infection status, especially in the regions where rodent-borne diseases were detected in the past.
The East Azerbaijan Province is located in the northwest of Iran, a region known as a historical focus for two major rodent-borne diseases, that is, plague and tularemia (Arata et al., 1973, Hashemi Shahraki et al., 2016, Karimi, 1980). This province has also experienced several rodent-borne disease outbreaks in the past (Rabiee et al., 2018). For instance, in 2018, a significant outbreak of hepatitis E was reported in East Azerbaijan, affecting more than 10 provinces (Rabiee et al., 2018). Another notable outbreak was the Crimean–Congo hemorrhagic fever, which was reported in the same year (Rabiee et al., 2018). These outbreaks, along with the presence of various rodent species in Iran, suggest a complex landscape for rodent-borne diseases in the region. The fauna and flora of the region could be affected by recent environmental changes such as massive deforestation, Uremia Lake drying up, lowering of groundwater, and human economic development. These ecological and anthropogenic changes may cause fluctuations among rodent populations, as well as among hosts and vectors of zoonotic agents (Mills et al., 1999). Investigating the species composition and infection level of rodents in this region will provide an early warning for the increased risk of transmission of rodent-borne infections, such as plague, tularemia, and arenaviruses to humans.
Plague is a life-threatening zoonotic disease caused by the Yersinia pestis bacterium (Bendiner, 1989). More than 350 mammalian species can be infected with Y. pestis, among them rodents being the primary hosts (Mahmoudi et al., 2021). This bacterium is transmitted to different animals and humans in various ways, including the bite of infected fleas, contact with contaminated fluid or tissue, inhalation of infectious droplets, and transmission from corpses and carcasses of infected animals (Perry and Fetherston, 1997; Steele, 1979). The most prominent arthropod vector for Y. pestis is the genus Xenopsylla, in particular X. cheopis (Perry and Fetherston, 1997). Plague is endemic in certain regions of Africa, Asia, and North and South America (Dubyanskiy and Yeszhanov, 2016), where the infection is enzootic in rodent populations (Perry and Fetherston, 1997). From 1943 to 1965, nine plague outbreaks have been reported in western Iran. For these outbreaks, rodents and hares were considered the principal reservoirs of Y. pestis (Hashemi Shahraki et al., 2016). The last report of Y. pestis among rodents in the Eastern Azerbaijan Province dates back to 1978 (Karimi, 1980). Over recent years, several plague outbreaks have been reported in Iran’s neighboring countries, such as Afghanistan, Saudi Arabia, and Jordan (Bin Saeed et al., 2005).
Tularemia is a zoonotic disease caused by the bacterium Francisella tularensis. The disease is transmitted to humans through various modes, including direct contact with infected animals (such as handling of infected animals), bites from arthropods (ticks, flies, and mosquitoes), consumption of contaminated water or food, and inhalation of contaminated dust or aerosols (Hestvik et al., 2015). F. tularensis affects more than 200 species of mammals, birds, reptiles, and fish (Hestvik et al., 2015). This disease is common in the Northern Hemisphere in particular in the United States, Eastern Europe, Scandinavia, Russia, Kazakhstan, Turkmenistan, and Turkey, a country neighboring Iran (Akalın et al., 2009; Erdem et al., 2014; Tarnvik et al., 2004; Zargar et al., 2015). The first reports of tularemia in Iran date back to 1969 and 1970 in domestic and wild animals in the Azerbaijan Province (northwest) and Zabol County (southeast of Iran) (Arata et al., 1973). The first human case of tularemia in Iran was reported in 1980 in the Marivan district, Kurdistan Province (Zargar et al., 2015). Seroprevalence studies performed from 2011 to 2017 in Kurdistan, Sistan and Baluchistan, Ilam, and Lorestan Provinces showed that 14.4%, 6.52%, 2.78%, and 3.8% of studied human serum samples were positive for F. tularensis antibodies, respectively (Esmaeili et al., 2019a; Esmaeili et al., 2019b; Esmaeili et al., 2014a; Esmaeili et al., 2014b). Other studies from 2013 to 2015 from different provinces of Iran showed that 1.91% to 11.11% of trapped rodents were positive for F. tularensis by the serum agglutination test (Mostafavi et al., 2018; Zarger et al., 2015; Pourhossein et al., 2015). In 2015, 22.22% of tested surface water samples from northwest Iran were positive for Francisella spp. (Rohani et al., 2019).
Arenaviruses (family Arenaviridae, genus Mammarenavirus) are RNA viruses with mammalian hosts responsible for several emerging zoonosis diseases (Hallam et al., 2018; Korsman et al., 2012). Mammarenaviruses are divided into two serogroups, including the Old World (Lassa fever, lymphocytic choriomeningitis [LCM]) and the New World (Junin, Machupo, Guanarito, and Sabia) viruses, based on genetic differences and geographical distribution (Hallam et al., 2018). Muridae rodents (rats and mice) are the main reservoirs of arenaviruses in nature (Jay et al., 2005). Some arenaviruses are associated with person-to-person spread (e.g., Machupo), while transmission of others is related to contact with rodent excretions. The exact transmission route from rodents to humans is obscure, but transmission possibly happens through inhalation of aerosolized viruses or direct contact with viruses through mucosal or cutaneous routes. Infected rodents shed viruses in their urine, saliva, and other body excretions and secretions, which may deposit infectious material into the environment (Ölschläger and Flatz, 2013). Lassa virus is responsible for 300,000–500,000 infections and 5000–10,000 deaths annually in the endemic areas of West Africa. LCM virus is found worldwide because its natural host, Mus musculus, has an extended distribution (Shao et al., 2015). There is no information about the epidemiology of arenavirus in Iran and the neighboring countries.
Although studying the potential for transmission of the abovementioned agents in each geographic region is essential for public health, little is known about the current biodiversity, the host range of these pathogens, and the infection status of the animal host populations in East Azerbaijan. Therefore, the present study aimed to evaluate the diversity within small-mammal communities in the eastern Azerbaijan Province, detect their ectoparasites, and then the rodent infection rate with Y. pestis, F. tularensis, and arenaviruses.
Materials and Methods
Study area
East Azerbaijan Province is bordering the Republic of Armenia, the Republic of Azerbaijan, and Ardabil Province of Iran in the north, West Azerbaijan Province in the west, and Zanjan Province in the south. To obtain an unbiased estimate, seven counties, including Sarab, Ahar, Azar Shahr, Kaleybar, Bonab, Ajabshir, and Maragheh (Fig. 1) were randomly selected to study.
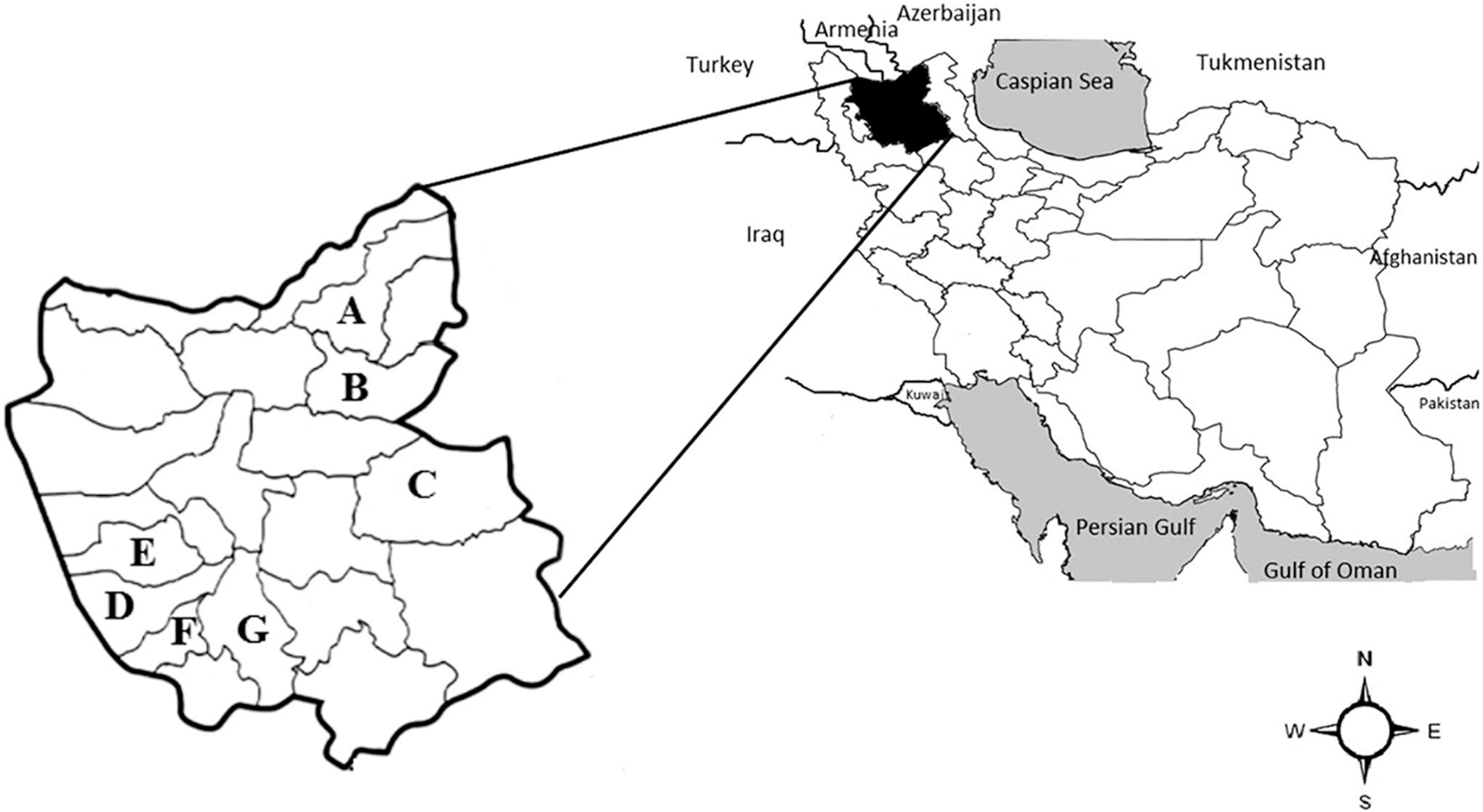
East Azerbaijan Province and its counties. The sampling of rodents was conducted in Kaleybar
Trapping and identification of small mammals
Rodent sampling was done in two stages. The first phase of this study was conducted in the fall of 2017, and the second phase was in the spring of 2018. Trapping was conducted using traditional live traps with bait. Spatial information on rodent trap lines was recorded using the Global Positioning System.
External morphology, standard external measurements (head and body length, tail length, hindfoot length, and ear length), weight, and gender of each trapped animal were recorded. Species identification was primarily based on available morphological criteria (Darvish et al., 2014; Kryštufek and Vohralík, 2001).
Collection and identification of ectoparasites
Captured small mammals were handled with long forceps on top of a basin filled with water. Ectoparasites were released into the water by slowly brushing and blowing in the rodent fur. They were collected from the water with forceps and placed into individual microtubes filled with 70% ethanol. Animals were euthanized following ethical rules (Sikes and Animal Care and Use Committee of the American Society of Mammalogists, 2016). The samples were sent to the Research Centre for Emerging and Re-emerging Infectious Diseases (RCERID) at the
Tissue and serum sample collection from small mammals
After euthanizing the rodent and collecting blood following the standard guidelines for working with animals, as outlined by the American Veterinary Medical Association (2016), the rodent was immediately dissected, and the spleen and lung tissues were harvested by a pathologist using an aseptic technique. Serum samples of small mammals were obtained by centrifuging 5 mL of clotted whole-blood specimens and investigated for serological analysis of Y. pestis and F. tularensis.
Spleen tissues were harvested from small mammals for molecular identification of Y. pestis and F. tularensis. Lung tissues were collected for arenavirus detection and submerged in microtubes containing RNAlater solution (Qiagen, Germany; each 200 mg sample in about 1.5 mL of RNAlater solution) for cellular RNA stabilization, protection, and storage.
All samples were sent to the RCERID laboratory in the
Serological survey of antibodies against Yersinia pestis and Francisella tularensis
An enzyme-linked immunosorbent assay (ELISA) was used to detect antibodies against the capsular F1 antigen of Y. pestis. All antigens, positive and negative control serums, antirodent secondary antibodies, and solutions necessary for ELISA were obtained from the Pasteur Institute of Madagascar (World Health Organization’s reference center for plague). All steps were done according to the protocol of the Pasteur Institute of Madagascar (Chanteau et al., 2003).
The serum sample of each rodent was tested for serological detection of F. tularensis using the standard tube agglutination method. Whole-cell antigen and control sera were prepared from Bioveta, Inc. (Ivanovice, Czech Republic). The test procedure was carried out according to the manufacturer’s instructions. Briefly, the sera were serially diluted using the diluent solution (1:10, 1:20, 1:40, etc.). Half a milliliter of diluted sera was mixed with an equal volume of diluted antigen and then incubated at 37°C for 20 h. Agglutination was then assessed after subsequent incubation at room temperature for 1 h. According to the protocol, visible agglutination at titers ≥1:80 was considered positive, while a titer of 1:40 was recorded as suspect (Gyuranecz et al., 2011).
DNA extraction and real-time quantitative polymerase chain reaction (qPCR) for identification of Yersinia pestis and Francisella tularensis
DNA extraction from spleen samples was done using the High Pure PCR Product Purification Kit (Roche, Germany). Extracted DNA was tested by qPCR for the detection of F. tularensis (ISFtu2 and fopA genes) and Y. pestis (yihN, caf1, and pla genes) using the specific primers and probes that are shown in Table 1 (Francy et al., 2009; Rohani et al., 2018; Stewart et al., 2008; Versage et al., 2003). The qPCR mixtures contained 4 μL of extracted DNA, 10 μL of 2X Real Q Plus Master Mix for Probe (Ampliqon, Denmark), and 1 μL of a mixture of 200 nM probe, 900 nM forward, and 900 nM reverse primers that was finally volumed to 20 μL with 5 μL of double-distilled water (DDW).
Primer and Probes Used for Detection of Francisella tularensis and Yersinia pestis
Amplifications were carried out on the Corbett 6000 Rotor-Gene system (Corbett, Victoria, Australia) with the following program: 10 min at 95°C, followed by 45 cycles of 15 s at 94°C and 60 s at 60°C. The qPCR results were interpreted using Rotor-Gene® Q 2.3.5 software (QIAGEN).
DDW was used as a negative control in the molecular detection methods. The Y. pestis positive control was a cloned plasmid (pUC 57, provided by the Pasteur Institute of Iran). The F. tularensis positive control was DNA from the F. tularensis subsp. holarctica NCTC 10857.
For quality and quantity examination of extracted DNA samples, the absorbance at 260, 280, and 230 nm was measured using a microplate spectrophotometer (Epoch BioTeK, USA). Absorbance ratios of 260/280 nm ∼1.8 and 260/230 nm ∼2.0–2.2 were considered good quality.
Arenavirus RNA extraction and quantitative real-time reverse transcriptase-PCR (RT-qPCR)
Total RNA was extracted from homogenized lung tissues of collected rodents using a High Pure Viral RNA kit (Roche, Germany) according to the manufacturer’s instructions. The quantity of extracted RNA was determined by spectrophotometry (Epoch BioTek, USA). A ratio of absorbance at 260/280 nm ∼2.0 and 260/230 nm ∼2.0–2.2 was accepted as pure for RNA. In addition, RNA integrity was observed by running the RNA on a 1% standard agarose gel.
The RNA was immediately used for the qPCR assay. The Pan-arenavirus quantitative real-time RT-qPCR was performed (Vieth et al., 2007) using QIAGEN One-Step RT-PCR Kit reagents (Qiagen, Hilgenberg, Germany). The 25 µL mixture contained 5 μL of 5x Qiagen OneStep RT-PCR Buffer, 0.3 μM of each primer (5′-AGAATYAGTGAAAGGGARAGYAAYTC-3′ and 5′-CACATCATTGGTCCCCATTTACTRTGATC-3′) (Atkinson et al., 2014), 1 μL of Qiagen OneStep RT-qPCR Enzyme Mix, 100 μM each of the four dNTPs, 1U/reaction RNase inhibitor, 3 μL of extracted RNA (1 pg–2 μg/reaction), and 13.25 μL of RNase-free water.
Synthetic plasmid and RNase-free water were used as positive and negative controls for the RT-qPCR, respectively.
The amplification reaction was performed in a BioRad thermocycler (BioRad, CA, USA) using the following temperature profile: 50°C for 30 min and 95°C for 15 min, followed by 45 cycles of 95°C for 30 s, 55°C for 60 s, and 72°C for 10 min. The PCR products (395 bp) were visualized by electrophoresis on a 2% agarose gel stained with DNA-safe stain (Yekta Tajhiz Azma, Iran).
Results
Identification of small mammals and ectoparasites
A total of 205 rodents, four Eulipotyphla, and one carnivore were captured from seven districts of the East Azerbaijan Province (Sarab [n = 50], Ahar [n = 45], Maragheh [n = 38], Bonab [n = 34], Azar Shahr [n = 27], Kaleybar [n = 9], and Ajabshir [n = 2]; Fig. 1). Captured rodents belonged to 10 genera, including Meriones, Microtus, Rattus, Mus, Apodemus, Nothocricetulus, Arvicola, Dryomys, Chionomys, and Scarturus. Most of the captured rodents belonged to the jirds of the genus Meriones (123 out of 205), with Meriones persicus being the most prevalent captured species (112 out of 123). Eulipotyphla belonged to three genera, including Crocidura, Erinaceus, and Hemiechinus, and the Carnivora belonged to the genus Mustela (Table 2).
Captured Small Mammal Species and Their Associated Flea Species, and the Localities and Geographic Coordinates of the Animal Capture Sites in East Azerbaijan Province, Iran
Positive samples of Francisella tularensis with ISFtu2 and fopA. No positive cases of Yersinia pestis or arenavirus infections were detected.
A total of 317 fleas were removed from all animals. In contrast, no ticks were collected. Flea species belonged to Xenopsylla buxtoni (53.63%; 170/317), Xenopsylla nuttalli (26.18%; 83/317), Stenoponia tripectinata (3.47%; 11/317), Paraceras melis melis (7.26%; 23/317), Ctenophthalmus rettigi smiti (1.89%; 6/317), Rhadinopsylla bivirgis (2.21%; 7/317), Paradoxopsyllus grenieri (4.10%; 13/317), and Nosopsyllus iranus iranus (1.26%; 4/317). All fleas were collected from M. persicus, Meriones vinogradovi, Apodemus witherbyi, Microtus socialis, Microtus mystacinus, and Mustela nivalis (Table 2).
Detection of Yersinia pestis, Francisella tularensis, and arenavirus
Serum samples from all captured animals were negative for antibodies against the capsular F1 antigen of Y. pestis and F. tularensis antigen. The qPCR and RT-qPCR tests were negative for Y. pestis and arenaviruses, respectively. Five M. persicus tested positive for F. tularensis (ISFtu2 and fopA genes) in the qPCR results. Four of these positive samples were collected from the Maragheh County and one from the Azar Shahr County.
Discussion
In this study, F. tularensis infection was detected in rodents from northwest Iran. The study did not find any Y. pestis and arenavirus infection in these animals using the molecular detection method. However, most of the collected small animals (rodents and Eulipotyphla) had the potential to carry Y. pestis and F. tularensis infections.
Reporting the F. tularensis-positive samples from animals in the northwestern and southwestern parts of Iran confirms that tularemia is extensively scattered in Iran.
An investigation conducted from 1969 to 1970 on serum samples from more than 4,600 small mammals and 200 cattle and sheep showed that F. tularensis is present in the northwest of Iran and at one location in the very east of the country (Arata et al., 1973). Further studies have confirmed that F. tularensis is also present in other parts of Iran. The first human case of tularemia was reported in 1980 in Marivan, the Kurdistan Province of Iran (Karimi et al., 1981). More recent studies carried out from 2011 to 2017 have shown F. tularensis antibodies in human blood samples from various regions of Iran (Esmaeili et al., 2014a; Esmaeili et al., 2014b; Khoshdel et al., 2014). In a study conducted in 2013, infected rodents were reported from the Sistan and Baluchistan Province (southeastern Iran) (Pourhossein et al., 2015). In another study in the Kurdistan Province, 4.8% of the 140 collected rodents were seropositive for F. tularensis, including Microtus spp., Meriones spp., Mus macedonicus, Dryomys nitedula, and Calomyscus spp. (Zarger et al., 2015). Several studies conducted between 2014 and 2015 revealed the presence of F. tularensis in different Iranian provinces. Positive samples for F. tularensis were observed in three rodents (Apodemus uralensis, Mus musculus domesticus, and Chionomys nivalis) and two hares (Lepus europaeus and Lepus sp.) in the north, southeast, and southwest of Iran (Mostafavi et al., 2018).
Tularemia outbreaks following increases in rodent populations have revealed the important role of rodents in the transmission of F. tularensis to humans and the importance of monitoring these animal reservoirs as an early warning system (Rodríguez-Pastor et al., 2017). In our study, qPCR-positive samples for the F. tularensis ISFtu2 gene were detected in four M. persicus in Maragheh (4/27) and one M. persicus in Azar Shahr (3.7%). Most rodent species with a significant role in the transmission of F. tularensis throughout the world are belonging to the genera Microtus (vole), Arvicola (water vole), Apodemus (field mice), and Clethrionomys (red-backed vole) (Gyuranecz et al., 2010). In the present study, all positive samples belonged to the Persian jird (M. persicus), which is mainly distributed through the Iranian plateau and adjacent territories (e.g., Afghanistan, Armenia, Azerbaijan, Iraq, Pakistan, Turkey, and Turkmenistan). Therefore, M. persicus may represent a major threat as a reservoir of F. tularensis in the aforementioned countries as well.
Ticks act as the major natural vector for F. tularensis and may transmit the infection to humans and animals (Sjöstedt, 2007). The prevalence of F. tularensis infection in ticks varies worldwide and has been reported in diverse tick genera and species such as Dermacentor, Hyalomma, Haemaphysalis, Ixodes, and Rhipicephalus (Rahravani et al., 2022). These species have been observed in livestock in various parts of Iran (Fayazkhoo et al., 2017). In a recent study in Iran on ticks collected from sheep and goats, 1.22% of Dermacentor marginatus (D. marginatus) ticks were infected with F. tularensis (Rahravani et al., 2022). The genus Dermacentor is a primary vector for F. tularensis in the United States (Zellner and Huntley, 2019). In southwestern Europe, F. tularensis were detected in 25.7% of Rhipicephalus sanguineus and 2.4% of D. marginatus (de Carvalho et al., 2016). However, the transmission of F. tularensis via Xenopsylla cheopis has also been reported under experimental conditions (Prince and McMahon, 1946). Since 2000, many human cases have been reported for tularemia every year in Turkey (Kutlu et al., 2021; Sahin et al., 2007; Zargar et al., 2015), the country that borders the west of Iran; therefore, more attention should be given to this disease in Iran, particularly in rural areas (Esmaeili et al., 2021). Although the sampling area in the present study is an endemic region for tularemia in Iran and is located in the neighborhood of Turkey (an endemic country), we found a low detection rate of F. tularensis. Considering ticks are the main vector of F. tularensis (Sjöstedt, 2007), the failure to find ticks (possibly for climate or seasonal reasons) in the geographical area and period of the study can explain this low prevalence. On the contrary, other sources of infection may play a role in tularemia outbreaks in Iran. For instance, water contamination was presumed as the source of the tularemia outbreak in northwestern Iran in 2016–2018 (Esmaeili et al., 2021). Therefore, further assessment of potential F. tularensis reservoirs and vectors is a key forward step in predicting the dynamics of the infection in Iran.
Plague outbreaks were reported regularly in the past in Iran (Hashemi Shahraki et al., 2016). During 1772–1773, one of the biggest epidemics of plague in the world happened in Iran and the neighboring regions. This outbreak caused approximately two million deaths (Hashemi Shahraki et al., 2016). The last human plague case was reported in 1965–1966 in the West Azerbaijan Province (North West) and Kurdistan Province (west) of Iran (Mohammadpour et al., 2018). Recent serological studies in 2011 and 2012 showed antibodies against Y. pestis in 1% (1/98) of rodents and 3.5% (4/117) of dogs in the Kurdistan-Hamadan border, which would imply the circulation of plague infection in these regions (Esamaeili et al., 2013). Despite the existence of active sylvatic foci in the western and northwestern parts of the country, no human case has been reported for several decades.
In natural foci of plague in Iran, four species of the genus Meriones, including M. libycus, M. persicus, M. vinogradovi, and M. tristrami, play a significant role as reservoirs and in the transmission of this disease (Hashemi Shahraki et al., 2016). The most abundant rodent species collected in this study were M. persicus (94/200, 47%), M. socialis (21/200, 10.5%), and Apodemus witherbyi (18/200, 9%). M. persicus is a well-known reservoir for both Y. pestis and F. tularensis in Iran (Mostafavi et al., 2017a). Since it carries vectors for both diseases, our study highlights that there is a high potential for the occurrence of human plague and/or tularemia cases in East Azerbaijan. Fleas are the principal vectors of Y. pestis. In the present study, several fleas were isolated from different mammal species, including M. socialis, M. mystacinus, A. witherbyi, M. nivalis, M. persicus, and M. vinogradovi. The most prevalent flea was X. buxtoni, which is known as a significant vector of Y. pestis in Iran (Baltazard et al., 1960; Karimi et al., 1981). Our failure to find evidence of Y. pestis in the studied areas could be related to the reservoir population fluctuations and dynamics/movements under ecological changes and also the small sample size of analyzed animals. Comprehensive sampling of the potential reservoirs and vectors is needed to get a better overview of this disease in the studied area.
In the present study, although RNA extraction and RT-qPCR were performed according to the standard protocols, arenavirus infection was not detected in the East Azerbaijan Region in the collected small mammals. Our negative results can be explained for the following reasons: (1) collection of samples from areas that were not endemic for this infection, and (2) different reservoirs of this disease in Iran. In fact, the reservoirs of arenaviruses in Iran might not include the specific species of rodents that we sampled, or the virus might not be present in the same abundance as in other regions. For instance, the presence of specific rodent species that are known reservoirs of arenaviruses in other parts of the world might be absent or less prevalent in Iran. This could result in a lower prevalence of the virus, leading to negative results in our study.
Various human illnesses are caused by rodent-borne arenaviruses. Arenavirus infections contribute significantly to the human disease burden in Africa and the Americas, but little data exist from Asia. To date, only two arenaviruses, the broadly dispersed LCM virus and the newly reported Wenzhou virus, have been recognized in Asia (Blasdell et al., 2016). There are not any reports of arenaviruses in Iran and the neighboring countries despite arenavirus infections continually being observed worldwide. For instance, several imported Lassa fever cases with hazardous outcomes have been reported in different parts of the world, including North America, Europe, and Asia (Mazzola and Kelly-Cirino, 2019). So, there is a need for more advanced screening of animal reservoirs and carriers of the virus in currently nonendemic areas. The present study has several limitations. First, the absence of arenavirus rodent-borne infections in the study location in northwestern Iran does not imply the absence of this infection in Iran. Therefore, further investigation with a larger animal sample size should be performed to examine this hypothesis. Second, the study did not investigate the prevalence of Y. pestis, F. tularensis, and arenavirus infections among the collected ectoparasites. Future research could simultaneously investigate rodents and vectors. Third, the study did not consider the effect of seasonal patterns and mating/fighting on the prevalence of disease within the study population. These factors could significantly influence the spread and prevalence of diseases among rodents, and should be considered in future studies.
Since we detected F. tularensis in some captured rodents, it seems important to recommend conducting a further study by testing surface water, insect, and livestock samples collected from the surveyed areas to further explore reservoirs of F. tularensis and the modes of transmission of this pathogen to humans.
Conclusions
In conclusion, this study highlights for the first time molecular detection of F. tularensis among small mammals in northwestern Iran, indicating the endemic nature of tularemia in the country. Therefore, health officials, physicians, and health care workers must be informed of the public health threat associated with rodent-borne diseases. The diversity of trapped rodents and fleas and various climatic conditions in these study areas indicate the potential for the presence and spread of various rodent-borne diseases, especially plague and tularemia. Therefore, larger scale studies on animal reservoirs and potential vectors of Y. pestis and F. tularensis in other regions of northwestern Iran are needed to deepen our knowledge of the current status of these two extremely dangerous diseases, considering the influence of seasonal patterns, mating, and fighting behaviors.
Footnotes
Acknowledgments
We sincerely thank Hamed Hanifi, Dr. Hossein Ahangari Kohan, Dr. Majid Hemmati, Alireza Moradi, Hesam Zali, and Seyyed Adel Hosseini for their help in the sampling.
Authors’ Contributions
E.M, S.Es, M.SV, S.Ey, Ch.D, M.M, V.N, AL, and JP.H.: Conception and design of the work. E.M: Acquisition. S.E, A.Ma, M.SV, A.Gh, M.R, and A.Mo.: Investigation. E.M, S.E, and S.Ey.: Analysis and interpretation of data. R.M.: Writing—the original draft. E.M, N.B, S.Es, Ch.D, M.M, V.N, A.L, and JP.H.: Writing—editing. N.B: response to reviewers’ comments. All authors reviewed the article.
Ethics Approval and Consent to Participate
All animal experimental protocols in this study were complied with standard guidelines and approved by the Ethics Committee of the National Institute for Medical Research Development, Iran: approval number IRNIMADREC1396079. All methods were performed in accordance with relevant guidelines and regulations and the American Veterinary Medical Association (AVMA) guidelines (2020). The animals were not owned by individuals, and therefore, document informed consent from the client or owner was not applicable.
Author Disclosure Statement
The authors declare that they have no competing interests.
Funding Information
The National Institute of Medical Sciences Research Development (grant no. 957145) and the Pasteur Institute of Iran (grant no. 1050).