
Abstract
Background:
Marsupials and rodents are the most important wild and synanthropic hosts of Trypanosoma cruzi due to the high frequency of infection, maintenance of diverse genetic populations of the parasite, and their close proximity to interact with both transmission cycles, sylvatic and peridomestic. Our aim was to identify the discrete typing units (DTU) of T. cruzi from different wild and synanthropic hosts in two regions of Mexico and to carry out a review of historical data focusing on current knowledge on the diversity and T. cruzi DTUs of host species.
Materials and Methods:
One hundred fifteen samples were obtained from two areas in Tabasco and Nayarit state. The presence of T. cruzi was evaluated by PCR.
Results:
The 12.6% (12/95) of samples from Tabasco and 65% (13/20) from Nayarit were found to be positive for parasite DNA. All the sequences analyzed were grouped in T. cruzi DTU I; low nucleotide diversity was observed in Tabasco (π = 0.00566, and ϴ = 0.00632), while high genetic diversity was observed in Nayarit sequences, up to 8.63 (π) to 11.10 (ϴ) times greater than Tabasco sequences. Genetic flow and migration between Tabasco, and Nayarit were scarce (FST = 0.37329 and Nm = 0.42), and genetic exchange was observed only between nearby areas. The bibliographic review of hosts in Mexico, together with our data, shows a heterogeneous T. cruzi prevalence in Chiroptera and domestic animals. For Atelidae and Canids, prevalence is generally below 25%. However, a high prevalence, greater than 25% and up to 100%, was recorded in Didelphimorphia, and Rodentia. Few studies in regions of Mexico have been described as infected with the parasite; in these, the genetic group with the highest prevalence is the DTU I.
Conclusion:
Marsupials and rodents are important reservoirs of T. cruzi; DTU I was frequently reported; however, recent genetic and reservoir studies have demonstrated the presence of greater diversity of genetic groups.
Introduction
Chagas disease is an infectious disease caused by the hemoflagellate protozoan Trypanosoma cruzi. It is estimated that 6–7 million people are infected with this parasite worldwide, mainly in Latin American countries where it is considered a major public health problem (WHO, 2023).
Wild and synanthropic mammalian hosts play essential roles in the conservation, maintenance, and interconnection of the domestic and sylvatic cycles of T. cruzi. Some species of bats, marsupials, xenarthrans, and rodents are considered important reservoirs of T. cruzi due to their opportunistic food habits, wide geographical distribution, and high rates of T. cruzi infection (WHO, 2012). In Mexico, several species belonging to different mammalian orders have been recorded as T. cruzi hosts. Among them, some species stand out because of their high levels of T. cruzi prevalence, including Artibeus jamaicensis, Didelphis virginiana, Didelphis marsupialis, Mephitis mephitis, Neotoma micropus, Rattus rattus, and Peromyscus melanophrys (Villegas-García and Santillán-Alarcón, 2004; Carnevali et al, 2017; Galaviz-Silva et al, 2017). As synanthropic animals, it refers to a “behavior of free-ranging animals benefiting from the shared ecology with humans” (Klegarth, 2016), they are well adapted to human dwellings and domestic animals, with a constant circulation in domestic and wild environments.
Species of marsupials and rodents share biological attributes that favor parasite maintenance and transmission of T. cruzi, such as a flexible diet, high reproductive rate, and high adaptedness to disturbed habitats (Ruiz-Piña and Cruz-Reyes, 2002; Meerburg et al, 2009; WHO, 2012; Ruiz-Piña et al, 2018); they also play public health and veterinary importance, due to their capacity to cause disease in humans, domestic animals, and wildlife. They are related to the increasing number of infected vectors distributed in the same area, favoring the risk of human infection from those areas where both reservoirs and vectors are present (Ramos-Ligonio et al, 2012; Hernández-Cortazar et al, 2018; Lopez-Vivas et al, 2018; Rosal et al, 2018). Recently, it has been suggested that bats might be relevant harbors and dispersers of T. cruzi discrete typing units (DTUs) (Austen and Barbosa, 2021).
Genetic studies of parasites coupled with host identification at regional or local scales are important to understand host–parasite relationships. However, the role of the species of animals involved in the T. cruzi transmission cycle is poorly understood. In Mexico, reports of T. cruzi prevalence in different species of reservoirs have been documented; however, there are regions where more studies are concentrated than others, such as the southern and central regions of the country compared with the northeastern region. Furthermore, the prevalence reported between groups and regions is heterogeneous. For instance, in the northeastern region, the general prevalence in small wild mammals and dogs was 14.5% (Galaviz-Silva et al, 2017); in the central region, it was of 24% in opossum (Carnevali et al, 2017); and in states located in the southeast region, the prevalence varies between species, 9.05% in white-nosed coati, 21.40–23.39% in nonhuman primates, and 26.6% in raccoon samples (Martínez-Hernández et al, 2014; Rovirosa-Hernández et al, 2021). More detailed studies had been carried out in common raccoons (Procyon lotor) and white-nosed coati (Nasua narica) in Tabasco (southeast region), where the first species had a higher prevalence of T. cruzi and more persistent parasitemia, also a marked heterogeneity in antigen recognition (Martínez-Hernández et al, 2014; Villalobos et al, 2020). The identification of species infected with T. cruzi and the determination of genetic lineages would be steps to follow in establishing the role they play in the transmission and maintenance of the parasite.
On the other hand, six different genetic groups or DTUs of the parasite have been described (DTU I to VI), which are associated with specific hosts, ecotopes/niches, geographical ranges, clinical manifestations, and levels of pathogenicity (Zingales, 2018). An additional DTU, TcBat, has been linked to neotropical bats (Marcili et al, 2013). Particularly, DTU I is the genotype most frequently associated with the sylvatic environment; however, significant intraspecific genetic variability was observed in this group, and this allowed us to clarify the idea that DTU I can participate in other transmission cycles. The characterization of DTU I has separated them into five haplotypes, which are associated with different cycles of infection, vectors, or reservoirs (Herrera et al., 2009; Cura et al, 2010). In Mexico, DTU I is the most prevalent genotype reported, but others (II to VI) have also been identified in humans and vectors mainly (Ramos-Ligonio et al, 2012; Lopez-Cancino et al, 2015; Rangel-Gamboa et al, 2019).
Furthermore, the characterization of parasite genotypes may help to understand the epidemiology of Chagas disease in all transmission cycles. The aim of this work was to identify the T. cruzi genotypes (DTUs) associated with mammalian hosts in Mexico through a laboratory and meta-data approach. For this purpose, synanthropic and wild opossums, rodents, and bats were analyzed in four locations belonging to two states of Mexico. In addition, the flow and genetic variation between the study states were evaluated. Finally, we discuss the role of different groups of wild and synanthropic mammals, as well as the main domestic animals that may play in the maintenance and transmission of T. cruzi.
Materials and Methods
Study area
Areas were selected due to the history of T. cruzi infection in vectors, humans, and/or reservoirs. In Nayarit, nine species of vectors had been documented (Cruz-Reyes and Pickering-López, 2006); particularly, Triatoma longipennis and Triatoma picturata had been found infected with T. cruzi, and associated with wild and domestic habitats in the same areas as our study (Magallón-Gastélum et al. 2001). In nearby states, the infection index in triatomines was 29.3% for T. longipennis and 27.4% for T. picturata (Martínez-Ibarra et al. 2001). In Tabasco, Triatoma dimidiata is the mainly distributed species; in areas near the state, this species has been found to be infected with T. cruzi (38%), and associated with cases of infection in humans (Monteon et al, 2013). In addition, studies in reservoirs had been described (Villalobos et al, 2020). The study was carried out in four locations in two states in Mexico, separated by a distance of approximately 1,300 km. Nayarit is located further north on the west coast, and Tabasco is in the southeast region. The experimental design involved the study of two specific sites within each state. In Nayarit, the analysis included animals from Compostela (21°14’14” N, 104°54’03” W) and Felipe Carrillo Puerto (21°07'50” N, 104°51'37” W) situated 15 km apart. Both sites are warm subhumid areas, with a temperature range of 19.6–25.2°C, relative humidity of 65.3%, and annual rainfall ranging from 780 to 2000 mm, with agricultural economic activity of tobacco, corn, beans, coffee, forage, and grain sorghum, mango, and livestock bovine, pig, and poultry. In Tabasco, sampling was conducted in La Florida (17°27' N, 92°45' W) and Villa Hermosa (18°00′05.39” N, 92°56′02.52” W), separated by 82 km. La Florida is a natural reserve located in the municipality of Tacotalpa; this area is warm and humid, with temperatures ranging from 18.5 to 42.5°C, 80% relative humidity, and annual rainfall ranging from 2900 to 3600 mm, with agricultural economic activity of corn, orange, lemon, cocoa, and sugar cane and livestock of cattle and poultry. DACBiol (Academic Division of Biological Sciences), UJAT (Juarez Autonomous University of Tabasco), located on the outskirts of Villahermosa city, is warm and humid, with temperatures ranging from 15 to 39.3°C, 80% relative humidity, and annual rainfall ranging from 2000 to 4000 mm; its main economic activities are oil mining and trade (INEGI 1993, 2017). All sampling took place in peridomestic environments, except for La Florida, which is a Natural Reserve.
Capture and animal sampling
The animals were captured using different methods according to their size and characteristics. Each night, opossums were captured using 15–20 traps per day (Tomahawk Live Trap Company, Tomahawk, Wisconsin, USA), baited with canned sardines; and rodents were captured using 20 to 40 traps per day (H.B. Sherman Traps, Tallahassee, FL, USA), baited with oats and peanut butter. The traps were baited before sunset and checked early the next morning. Bats were captured using two 12-m mist nets per night. At each location, samples were collected over 5–8 nights between 2015 and 2016. Manual restraint was done, holding an animal by hand for a few moments, while the sample was collected. Once the animal had been immobilized, a blood sample was taken from the coccygeal vein for rodents and opossums, while in bats, it was obtained from the marginal wing vein. For each animal, according to its size, 20 µL to 2 mL of blood was collected using sterile syringes and placed in an Eppendorf® tube with lysis buffer for DNA extraction (Martínez-Hernández et al, 2014). To minimize fear, pain, or stress for the animals, the species and sex were only recorded. Captured animals were identified according to the field identification guides of Emmons and Feer, and Medellin et al. (Emmons and Feer, 1997; Medellín et al, 2008). At the end of sampling, the tail surface was marked with a gentian violet solution for short-term individual identification to avoid repeated sampling of the same individuals. Animals were released from the capture site after processing. The samples were transported under refrigeration and subsequently frozen at −20°C until further analysis.
DNA extraction and PCR assays
DNA extraction was performed using the phenol/chloroform technique (Martínez-Hernández et al, 2014). DNA concentrations between 0.5 to 1 μg were used for each PCR volume of 25 μL. The PCR assay was performed using the primers TCZ_F (5′-GCT CTT GCC CAC AMG GGT GC-3′) and TCZ_R (5′-CCA AGC AGC GGA TAG TTC AGG-3′), which amplify a region of 188 bp of Sat-DNA sequences (Souto et al, 1996), and TC1: 5′-GTG TCC GCC ACC TCC TTC GGG CC-3′; TC2: 5′-CCT GCA GGC ACA CGT GTG TGT G-3′; TCC: 5′-CCC CCC TCC CAG GCC ACA CTG-3′, which amplify a region of 300 or 350 bp for T. cruzi Lineage II and T. cruzi Lineage I, respectively, of Mini-exon (Elias et al, 2005). The master mix contained 2U of Taq polymerase, 12.5 μM of dNTPs, 1 (for the Mini-exon) or 1.5 mM (for the Sat-DNA) of MgCl2, and 0.1 mM of each primer. Amplification conditions were as follows: one cycle at 94°C for 5 min, 35 cycles, including denaturation, annealing, and extension steps at 94°C/30 s, 68°C/30 s (for the Mini-exon), or 56°C/10 s (for the Sat-DNA), and 72°C/30 s, and a final extension step at 72°C for 7 min. PCR products were visualized on 1.5% agarose gels stained with ethidium bromide and observed under UV light. T. cruzi DTU I (MHOM/MX/1994/Ninoa) and DTU II (MHOM/BR/00/Y) were used as a positive control in each amplification reaction; the amplicons were purified using the AxyPrep PCR Cleanup Kit (Axigen Biosciences, CA, USA) and sequenced on both strands by a commercial supplier.
Phylogenetic reconstruction and genetic variation analysis
Sequences obtained were subjected to BLAST (GenBank) searches to confirm the T. cruzi identity; the accession numbers could not be obtained due to the size of the sequence and the policies of GenBank. However, these were attached as supplementary material (Supplementary Data S1). Multiple alignments were performed using the Clustal W in MEGA software version 7.0.26 (Tamura et al, 2011). Phylogenetic reconstruction was conducted using a Bayesian approach (Ronquist and Huelsenbeck, 2003). The analysis was performed for 3,000,000 generations with sampling trees every 100 generations. The following T. cruzi DTU sequences from the GenBank were used as outgroup: KM657844-5, HM015642, and HM015652 as DTU I; AY520061, AY520068, AY520077, and FJ768487 as DTU II; FJ768484, FJ768481, and HM015649 as DTU III; EU178923 as DTU V; and AY520018, FJ768489, and HM015654 as DTU V. The genetic diversity analysis among sequences of locations was performed using DnaSP V6 software (Rozas et al, 2017). A Median Joining Network analysis was performed using NETWORK 4.611 software (fluxusengineering.com) with default settings and assumptions.
Statistical analysis
Prevalence and confidence interval of 95% were calculated for each category (site or species). Contrasts among categories were done by Chi-square test or Fisheŕs exact test. All analyses were done using the software OpenEpi®.
Host records in Mexico
The literature review was carried out during the first month of 2023 (Supplementary Data S2). We searched for records of T. cruzi hosts in Mexico using the PubMed and Web of Science database, entering combinations of the following key words: “DTU,” “genetic characterization,” “lineage,” “genotype,” “isozyme,” “isoenzyme,” “Mexico,” “reservoir,” and “Trypanosoma cruzi.” Articles in English and Spanish were included. Two reviewers worked independent of each other, collecting publications. At the end of the search, the databases were compared, eliminating those repeated publications; the search results were filtered to select only those articles reporting frequencies of infection with T. cruzi, DTUs, hosts, localities, and years of collection. The main data obtained from each article were reservoir species, states of Mexico, and DTUs of T. cruzi. We excluded those articles with case reports or negative infection results.
Ethics statement
Sampling and procedures were in accordance with the provisions of the Regulations of the Environment and Natural Resources Ministry with the license numbers SGPA/DGVS/04726/13 and FAUT-0250. The study was approved by a bioethics committee at UAM.DCBS.CICUAL.008.13.
Results
Trypanosoma cruzi prevalence
One hundred fifteen samples were obtained from four locations. The presence of T. cruzi in the samples was 12.6% (12/95) from Tabasco and 65% (13/20) from Nayarit. In particular, La Florida was 16.17% (11/68) and 3.7% (1/27) in Villahermosa from Tabasco, while Compostela was 80% (8/10) and Felipe Carrillo Puerto 50% (5/10) from Nayarit. By groups of mammals, a higher percentage of infection was observed in marsupials from Nayarit than Tabasco (63.6% vs 19%, respectively), while in rodents, only positive individuals were captured in Nayarit (6/9) (Table 1). Statistical differences among species, states, and regions were detected; prevalence was higher in Nayarit localities for rodents and marsupials than in Tabasco localities (p = 0.048 and 0.002, respectively), particularly for D. virginiana (p = 0.018). Among marsupial species from Tabasco, differences were detected only in D. marsupialis vs P. opossum (p = 0.031).
Mammals Evaluated for Trypanosoma cruzi Infection in Regions of Mexican States
Unidentified species, only genus or family was determined.
Trypanosoma cruzi identification and genetic variation analysis
Twenty five samples were positive by PCR-satellite DNA; however, only 18 could be sequenced, and due to the low sensitivity of PCR-miniexon, two samples were sequenced. The BLAST analysis showed high percentages of homology with T. cruzi, ranging from 97 (177/183) to 99% (181/182) identity with the DNA satellite, particularly with the sequences KM657844, HM015642, and HM015652 from GenBank, all of them corresponding to the DTUI of T. cruzi.
T. cruzi sequences from Tabasco showed low nucleotide (π = 0.00566) and haplotype (ϴ = 0.00632) diversity. In contrast, the parasite sequences from Nayarit sequences showed high genetic diversity (π = 0.04886 and ϴ = 0.07018), which was 8.63 (π) to 11.10 (ϴ) times greater compared with the T. cruzi sample sequences from Tabasco. Expected genetic flow and migration between T. cruzi sequences from Tabasco and Nayarit were scarce (FST = 0.37329 and Nm = 0.42), but some genetic exchange was detected between nearby regions, such as sequences from Felipe Carrillo Puerto and Compostela (FST = −0.01515 and Nm = ∞).
Phylogenetic reconstruction
All the sequences analyzed were grouped into T. cruzi DTU I, regardless of location, habitat type, or host. Almost all sequences from Nayarit (FCP6, FCP7, FCP8, FCP10, COM1, COM2, COM3, COM4, and COM7) were associated with the HM657844 GenBank sequence in a particular subclade (named DTU I subclade 1). In contrast, all sequences from Tabasco and one from Nayarit (FCP3, FLO1, FLO3, FLO4, FLO10, FLO12, FLO21, FLO32, and VIL9) were distributed in another subclade (named DTU I subclade 2), next to the HM015642 sequence (Fig. 1). Haplotype network analyses showed a diversity of sequences in the DTU I clade where Tabasco and Nayarit sequences were included (Fig. 2).
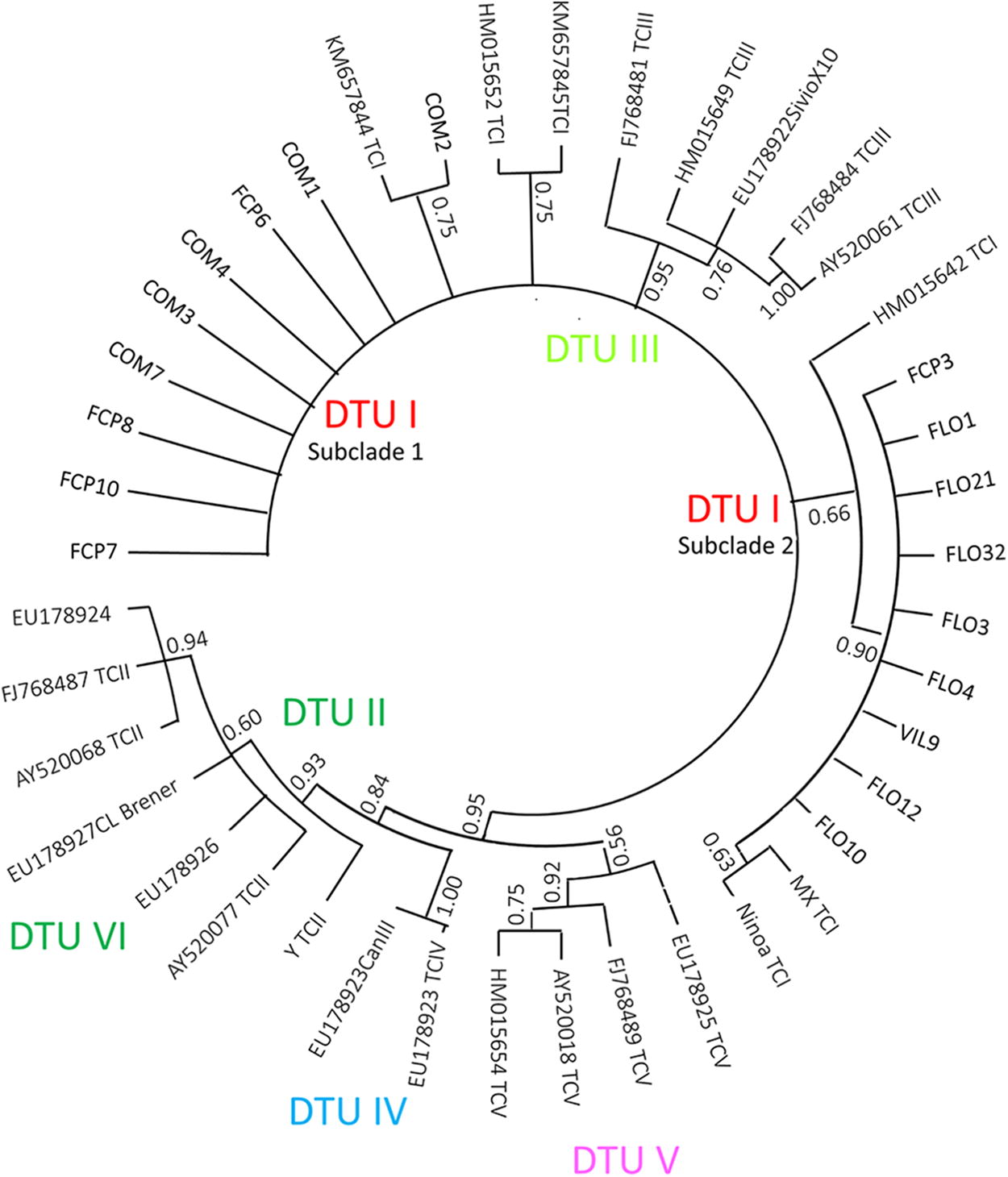
Bayesian phylogenetic tree using the DNA satellite sequences for different DTUs of Trypanosoma cruzi. The numbers of nodes indicate the values of support or posterior probability.
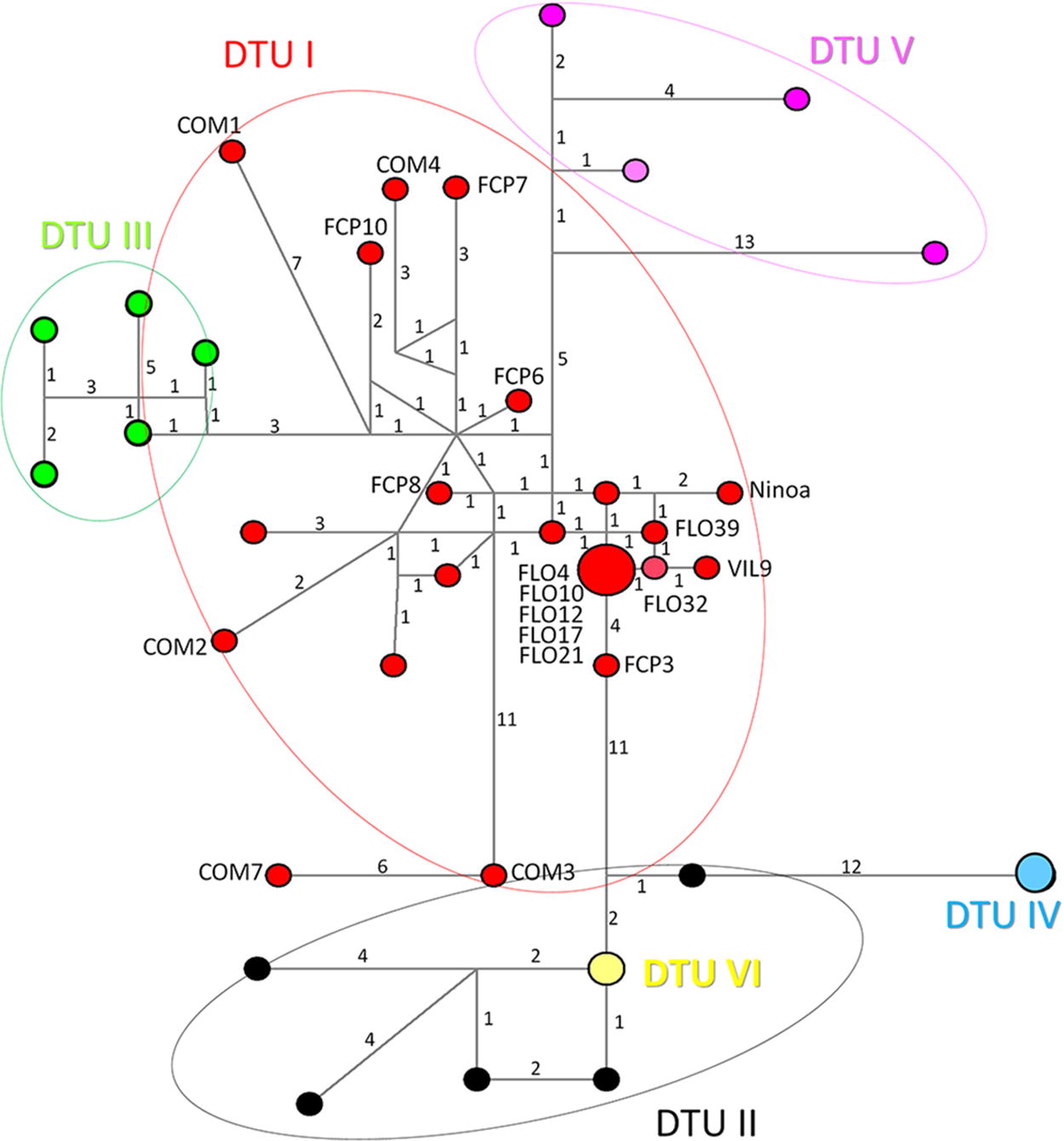
Haplotype networks of different Trypanosoma cruzi populations. The color of circles indicates the DTUs analyzed: Red circles = DTU I and Light Green circles = DTU III; Dark Green circles = DTU II and DTU VI; Blue circles = DTU IV and Pink circles = DTU V. The sequences obtained in this study are indicated next to the red circles.
Trypanosoma cruzi hosts in Mexico
Seventy four articles were recorded with data on T. cruzi and wild reservoirs between 1940 and 2023. Yucatan, Morelos, Jalisco, and Tabasco were the states with the highest number of reservoir studies. Two mammal orders (Rodentia and Chiroptera) and two families (Didelphidae and Atelidae) had the most numerous T. cruzi infection reports. The mammalian hosts with higher T. cruzi prevalence were Artibeus jamaicensis (69.5% to 92.8%), Didelphis virginiana (53% to 76.4%), Neotoma micropus (64%), Rattus rattus (47% to 86%), and Peromyscus melanophrys (50% to 100%). Particular species of carnivores, such as Procyon lotor, Nasua narica, Mephitis mephitis, and Urocyon cinereoargenteus, showed a single T. cruzi register. Domestic animal studies included Canis lupus familiaris, Felis catus, Sus scrofa domesticus, and, some other farm animals (Fig. 3). Studies of T. cruzi infection in dogs, reported a prevalence under 35%; other domestic animals, such as Bos taurus, Capra hircus, Equus caballus, Sus scrofa domesticus, and Ovis aries, showed heterogeneous results in some regions of Mexico (Fig. 3 and 4). There were reports in more than half of the states in the country (20/32), mainly located in the southeast of the country, such as Yucatan, where the largest number of host species was also reported. However, most states had reports before 1980, such as Zacatecas and Michoacan. Besides, few studies have carried out the genetic identification of the parasite (19%, 14/74), with DTU I being the most frequent and other genotypes (DTU II-III, V-VI) being restricted to Veracruz, Tabasco, Morelos, Oaxaca, and Yucatan (Fig. 4).
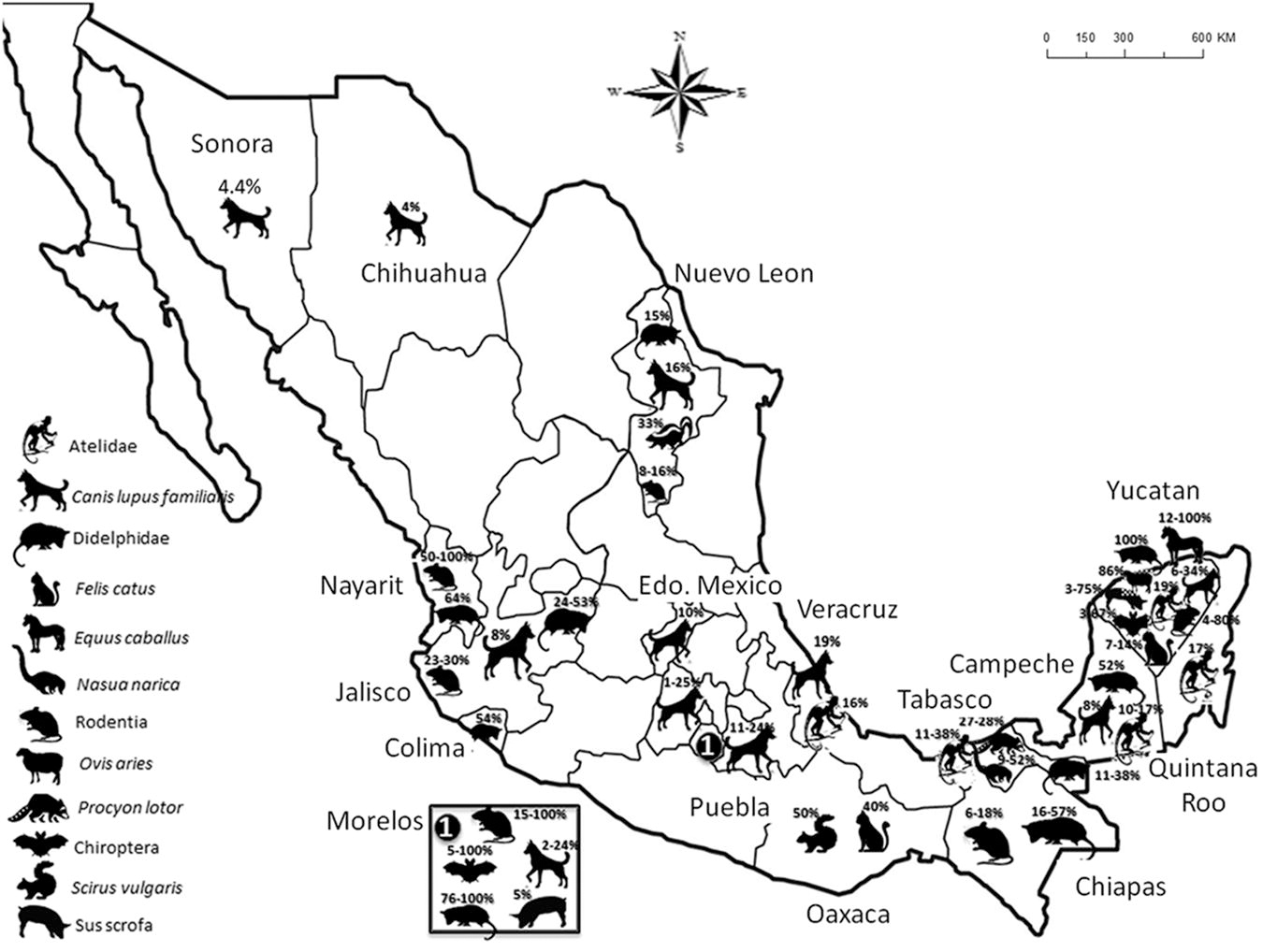
Distribution and percentage of infection of Trypanosoma cruzi hosts in Mexico.
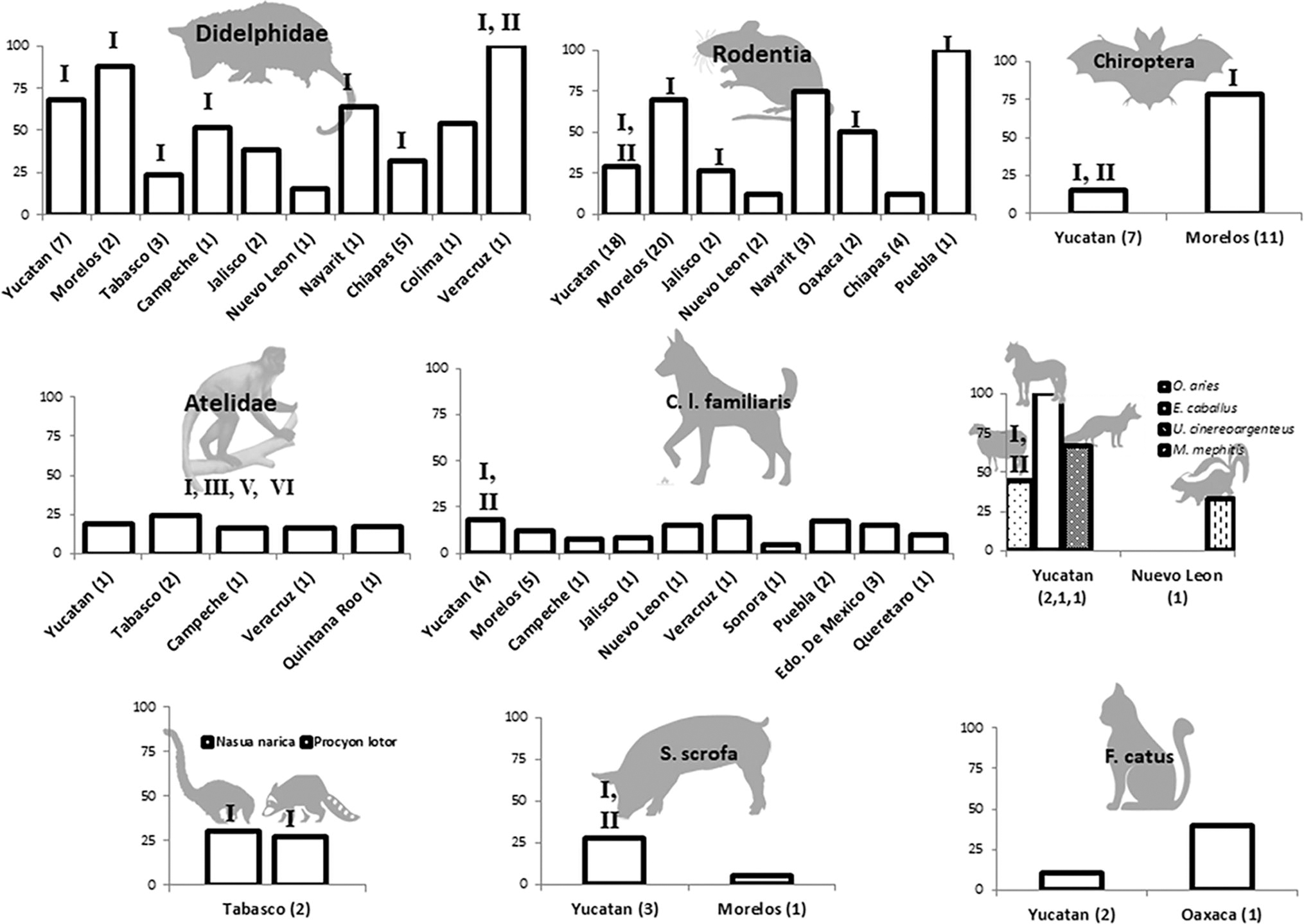
Trypanosoma cruzi in different hosts and regions in Mexico and associated DTUs. The vertical axis indicates the infection percentages. The number in parentheses indicates the number of records for each state. DTU associated by state and host is indicated at the top of the bar.
Discussion
Wild and synanthropic mammals play important roles in the maintenance and the link between the domestic and sylvatic cycles of T. cruzi, with a continuous flow of genetic populations and subpopulations of parasites between the three cycles (Villalobos et al, 2020). The proximity to the human population and the risk of parasite transmission have caused a polarization toward studies on domestic reservoirs. However, in a changing world, human development continuously transforms natural ecosystems, where wildlife is not only exposed to transmitting parasites to humans or to other species but also susceptible to acquiring new agents that may be harmful to its populations (Gibb et al, 2020). In Mexico, limited studies of wild T. cruzi reservoir species have been carried out. For instance, in Brazil, Jansen et al. (2018) carried out a review of T. cruzi infection in free-ranging wild mammalian fauna in eight different biomes; the biome that presented the most species included seven orders, 54 genera, and 67 species, without counting the species of bats. Only the above exceeds the number of species evaluated in the entire country. In this work, our first goal was to contribute to the knowledge of T. cruzi DTUs in wild reservoirs.
Mammals belonging to three different orders were captured: Didelphimorphia, Chiroptera, and Rodentia. Didelphimorphia is described as important in T. cruzi transmission because they can act as reservoirs and vectors, thus contributing to parasite maintenance in the sylvatic and peridomestic environment; and Didelphis, particularly, is a good habitat- and food-resilient genus and can easily adapt to peridomestic areas (Jansen et al, 2020, 2018). In Tabasco state, Didelphis marsupialis, Didelphis virginiana, and Philander opossum were captured, and at least one individual of each species was positive for T. cruzi. Didelphis marsupialis and P. opossum were captured only in sylvatic areas. Although few studies have been carried out for these species, it seems that D. marsupialis acts as a link between the wild and the domestic cycle, as suggested by Zavala-Velázquez et al. (1996), which is why the role of this species as T. cruzi host should be further studied in light of the current human impacts by destruction and invasion of wild environments. Philander opossum shares a habitat with D. marsupialis; as seen in Brazil, this species is exposed to parasite transmission cycles in all forest strata in the wild (Jansen et al, 2020). In Mexico, limited studies of P. opossum have been performed, and different topics, such as parasite prevalence, are necessary to achieve the best understanding of T. cruzi–P. opossum interactions, and in addition to their proximity to the domestic environment, they favor the exchange and contact of parasites with human populations and domestic animals.
On the other hand, in Villahermosa, the only animal captured positive for T. cruzi was D. virginiana; this synanthropic species is considered an ecological generalist well adapted to man-made environments and to a great range of habitats and climates; therefore, it is not uncommon to find them in different geographic areas of the country and infected with T. cruzi (Ruiz-Piña and Cruz-Reyes, 2002; Parada-López et al, 2013; Carnevali et al, 2017; Cab-Romero et al, 2020). The high infection rates observed in this study reinforce the importance of this species in maintaining the parasite and emphasize its great importance among T. cruzi hosts.
Conversely, in Nayarit, D. virginiana was the only Didelphidae species captured; interestingly, the number of captured animals was lower than that in Tabasco, but more infected animals were positive for T. cruzi, as has been observed in other states of Mexico where infection rates are heterogeneous (Ruiz-Piña and Cruz-Reyes, 2002; Parada-López et al, 2013). To our knowledge, there is no study of T. cruzi hosts in Nayarit; reports in near zones such as Jalisco showed D. virginiana with high percentages of seropositivity (53%), suggesting a similar Didelphid-T. cruzi infection rate in western Mexico (Carnevali et al, 2017).
Rodents are the main reservoirs for many pathogens and represent a potential risk to public health. In Mexico, more than 50% of rodent species that have relationships with vector co-occurrences and with T. cruzi reports were considered synanthropic, and more than 20% have been identified as T. cruzi hosts in a vector–host-co-occurrence network study (Ibarra-Cerdeña et al., 2017). In Mexico, the prevalence of T. cruzi in synanthropic rodents has been relatively low (6.2–14.28% for R. rattus and 0–11.1% for M. musculus) (Lopez-Cancino et al, 2015; Panti-May et al, 2017; Hernández-Cortazar et al, 2018), if the rodents are compared to other groups of mammals, such as didelphids. In this study, the rodents R. rattus and other samples classified only up to family level (Heteromyidae) were the only species captured in the peridomestic area from Nayarit, all of them were infected with T. cruzi; their prevalence was higher than that reported in the southern part of Mexico (14.28%; Hernández-Cortazar et al, 2018). These findings highlight the importance of R. rattus in peridomestic cycles of infection. Curiously, in our data on wild rodents, no species was T. cruzi positive from Tabasco. Species such as Heteromys gaumeri and Peromyscus yucatanicus have been reported with prevalence between 3.7 and 2.9%, respectively, in sylvatic habitats from the Yucatan Peninsula (Lopez-Cancino et al, 2015). On the other hand, Hernández-Cortazar et al. (2018) reported 6.25% of T. cruzi positivity in blood samples from four wild rodents, but this percentage corresponded to a single individual of the species Ototylomys phyllotis. However, infection by this parasite was not observed in Mus musculus, Sigmodon toltecus, Heteromys gaumeri, and Peromyscus leucopus, suggesting that some species are less susceptible to infection. For instance, Peromyscus leucopus and Peromyscus yucatanicus or Sigmodon toltecus, and Sigmodon hispidus (Ramsey et al, 2012; Lopez-Cancino et al, 2015). Other factors may be influencing the infection capacity, both intrinsic to the host and external. Because we cannot define the rodent’s state of well-being (nutritional status, age, coinfections, etc.), external factors, such as the habitat, could be influencing the negative result of infection, such as the case of S. hispidus.
The transmission dynamics of T. cruzi appears to be similar in both regions studied, where humans and other vertebrate hosts were found infected; infection frequency in marsupials and rodents between Nayarit (65%) and Tabasco (12.6%) was 5 times more in Nayarit. This results were similar to what was reported by Cruz-Reyes and Pickering-López (2006) in human cases, with 3.24 times more infection in Nayarit (12.19%) than in Tabasco (3.76%).
Chiroptera captures were only possible from the sylvatic area, but no animal tested positive. Our results were similar to those found in Morelos state, a state in central Mexico, by Ramsey et al. (2012), where no bat was T. cruzi positive; the absence of T. cruzi may be due to the small number of animals collected, or the detection methodology than can influence heterogeneous results between populations or species. For example, Sturnira lilium showed 61% of individuals infected with T. cruzi in direct examination of blood smear, while the same individuals, using histopathological examination of different sections, showed 22.6% to 61% of infection (Villegas-García and Santillán-Alarcón, 2004). The same species in Yucatan present range between 0 and 6.5% of T. cruzi infection depending on the habitat (domestic and sylvatic, respectively) using PCR (Lopez-Cancino et al, 2015). On the other hand, in states of the country closest to our study site, such as Campeche and Yucatan, infection frequency in several bat species was 30.2%, but unlike our work, the infection was only detected in renal tissue (Torres-Castro et al, 2021). The contrasting results in this group of mammals reflect that studies are still lacking in the country, despite that bats are considered fundamental in the persistence of T. cruzi and may be important reservoir hosts for human infection (Villegas-García and Santillán-Alarcón, 2004); they also have a key role in the dispersal of a certain DTU (TcVI) from the sylvatic habitat (Lopez-Cancino et al, 2015). Future studies directed to elucidate the transmission dynamics of T. cruzi in bats should be carried out with a large number of specimens from diverse geographical areas and with the use of highly sensitive and specific methods.
Analysis of T. cruzi sequences revealed the presence of DTU I in all samples, confirming the predominance of this genetic group in Mexico. Within the DTU I, we grouped the samples into two subclades (subclades 1 and 2). However, we could not determine, due to the difference in markers, if any of them was similar to TC Ia, using the intergenic region of spliced-leader (SL-IR) genes reported by Cura et al. (2010). TC Ia has been associated with sylvatic cycles in Central and North America, in wildlife and vectors in Mexico (Lopez-Cancino et al, 2015; Lopez-Vivas et al, 2018). On the other hand, Lopez-Cancino et al. (2015) showed that DTU I prevalence decreased according to an increase in habitat modification, from sylvatic to domestic habitats, giving way to other lineage groups. In the genetic tree, a separation between the geographical origin of samples was noted: a single group of parasites infecting and remaining in all hosts in the same region. Studies of mammal species and parasites in all habitats are still required to clarify these findings.
Our meta-analysis of studies in Mexico indicated that 62.5% of the states have records of wild hosts; however, states such as Zacatecas and Michoacan had reported before 1980, and therefore the need to carry out more studies to understand the wild hosts of T. cruzi in the country is a highlight. Yucatan, Morelos, and Jalisco had the highest numbers of records, which is in agreement with the data of the states with the highest numbers of publications about Chagas disease in Mexico (Cruz-Reyes and Pickering-López, 2006). Although the largest number of studies has been registered for those states, several of them evaluate the same areas or regions; for example, in more than half of the studies published for the state of Yucatan, at least seven of them analyzed the same sites or areas (Merida and surrounding municipalities). The above suggests that the study of reservoirs in Mexico is limited to a small percentage of areas in a state.
In wildlife, didelphids, rodents, and bats were still the most representative mammals according to their high infection rates with T. cruzi and the number of studies carried out on them. Mexico has around 550 species of mammals described; rodents and bats are the most diverse, contributing more than 70% of mammal species (Ceballos and Arroyo-Cabrales 2012). The abundance of these animals could be the reason for the greater number of captures and studies on them, which increases their importance as reservoirs of T. cruzi as they are widely diverse groups that occupy a wide range of habitats. Our data on didelphids and rodents add to that of the other authors that emphasize the impact of these animals on the maintenance of the parasite, particularly contributed with new data on rodent species. Both species, synanthropics, increase the zoonotic potential of transmission. On the other hand, other species of wild mammals that were highlighted in the records by the presence of T. cruzi and their increasing proximity to the domestic environment were Urocyon cinereoargenteus, Procyon lotor, and Nasua narica. The need to expand the study of T. cruzi wild reservoirs is decisive in the identification of genetic groups of the parasite; the exchange and maintenance of genetic groups of T. cruzi between domestic and wild environments could be crucial in determinants such as infection capacity or the preferential establishment of lineages. In domestic environments, the dog was the best studied domestic animal; interestingly, its infection percentages did not exceed 35% (Jiménez-Coello et al., 2010), which was below those reported in groups of wildlife animals and even other domestic animals, such as Ovis aries, Felis catus, or Equus caballus. Dogs are considered the most important domestic reservoir of parasites since infected dogs increase the risk of transmission inside human dwellings (Estrada-Franco et al, 2006). Studies on Mexican dogs have demonstrated a direct correlation between seropositivity in humans and dogs. In Mexico, the prevalence of Chagas disease varies widely between reports and places; however, some authors suggest that the national estimated seroprevalence of T. cruzi infection is 3.38% (Arnal et al, 2019). According to Arce-Fonseca et al. (2017), the dogs could act as sentinels for the disease in some regions of Mexico. On the other hand, DTU I was the predominant genotype in both wild and domestic mammals, as reported in humans and vectors; particularly, in Atelidae, an extensive diversity of DTUs (DTU I-III, V-VI) has been identified. Recently, mixed infection between T. cruzi lineages I (phylogenetically homologous to DTU Ia) and II (phylogenetically homologous to DTU VI) was identified in both sylvatic and domestic habitats in a town from Campeche state; particularly for DTU VI, bats would be relevant in sylvatic transmission, while rodent, sheep, pig and dog, even human, in domestic habitats (Lopez-Cancino et al, 2015).
Even though many studies have been performed for T. cruzi wild host identification, more than half of the states of the country remain without data. The study of wild mammals is not easy because the capture effort, animal and sample handling, and ethical issues, among other considerations, represent important challenges for a better understanding of the roles these animals play in the parasite cycle. Recently, the infection dynamics, the parasite’s impact on reservoirs’ health, and the diversity of reservoirs’ immune response have begun to be addressed, demonstrating that some reservoirs are more efficient in the transmission of T. cruzi since differences in blood counts and infection percentages, and recognition of epitopes of the parasite suggest this (Martínez-Hernández et al, 2014; Martínez-Hernández et al, 2016; Villalobos et al, 2020).
Conclusions
This study contributes to the knowledge of genetic populations of T. cruzi in mammals and describes regions that had not been evaluated. Marsupials and rodents continue to be important reservoirs of T. cruzi. Although several species of mammals were evaluated, the DTU I was the only one found. The above relates to the work carried out previously, where the diversity of DTUs has been associated with few species. The difficulties associated with fauna studies are reflected in the limited information available in the country, particularly the genetic populations of parasites, which are poorly described.
Footnotes
Authors’ Contributions
J.A.M.-I.: Conceptualization, methodology, resources, and writing—review and editing. B.O.-M.: Investigation. A.Z.R.-M.: Investigation and writing—review and editing. R.A.-F.: Investigation and writing—review and editing. C.I.M.-G.: Investigation, methodology, and writing—review and editing. E.R.-F.: Formal analysis, funding acquisition, investigation, methodology, and writing—review and editing. G.V.: Conceptualization, formal analysis, investigation, methodology, supervision, writing—original draft, and writing—review and editing. F.M.-H.: Conceptualization, formal analysis, funding acquisition, investigation, methodology, resources, supervision, and writing—review and editing.
Author Disclosure Statement
The authors declare no conflict of interest.
Availability of Data
All data generated or analyzed are included in this article.
Funding Information
No funding was received for this article.
Supplementary Materials
Supplementary Data S1
Supplementary Data S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.