
Abstract
Background:
Early detection and monitoring of SARS-CoV-2 infections in animal populations living in close proximity to humans is crucial for preventing reverse zoonosis of new viral strains. Evidence accumulated has revealed widespread SARS-CoV-2 infection among white-tailed deer (WTD), (Odocoileus virginianus) populations in the United States except in the southeast region. Therefore, the objective was to conduct surveillance for evidence of SARS-CoV-2 infection among WTD in Mississippi.
Materials and Methods:
Blood, kidney tissues, and nasal swab samples were collected in 17 counties from hunter-harvested deer during 2021-2022 and 2022-2023.Samples of kidney tissue were collected to evaluate for detecting antibody as a possible alternative to blood that is not always available from dead WTD. Nasal swab samples were tested for SARS-CoV-2 viral RNA by a RT-PCR assay. Sera and kidney tissue samples were tested for SARS-CoV-2 antibody by an enzyme-linked immunoassay (ELISA) and sera by a plaque reduction neutralization test (PRNT80).
Results:
The results of testing sera and kidney homogenate samples provided the first evidence of SARS-CoV-2 infection among WTD in Mississippi. The infection rate during 2021–2022 was 67% (10/15) based on the detection of neutralizing antibody by the PRNT80 and 26%(16/62) based on the testing of kidney tissue homogenates by an ELISA, and viral RNA was detected in 25% (3/12) of nasal swab samples. In 2022 to 2023, neutralizing antibody was detected in 62% (28/45) of WTD serum samples. In contrast, antibodies were not detected in 220 kidney homogenates by an ELISA nor was viral RNA detected in 220 nasal swab samples. Evidence of WTD activity was common in urban areas during the survey.
Conclusion:
Overall, the findings documented the first SARS-CoV-2 infection among WTD in Mississippi and showed that WTD commonly inhabited urban areas as a possible source of acquiring infection from humans infected with this virus.
Introduction
Early detection and surveillance monitoring of SARS-CoV-2 in wildlife populations are essential for understanding the continued risks as we transition from the pandemic to the endemic phase of SARS-CoV-2’s emergence. Abundant wildlife species living in proximity to humans are the obvious choice and primary focus for surveillance studies. Among the wildlife species susceptible to SARS-CoV-2 infection is the white-tailed deer (WTD), Odocoileus virginianus (Chandler et al. 2021). WTD are highly abundant and thrive in both rural and urban landscapes and are a popular game species (Chandler et al., 2021, Hearst et al., 2021, Kuchipudi et al., 2022). Experimental infection of WTD with SARS-CoV-2 causes a subclinical infection, elicits SARS-CoV-2 antibody, and produces infectious virus that is transmissible among WTD via social contacts, through feces, and bodily secretions (Chandler et al., 2021, Palmer et al., 2021, Vandegrift et al., 2022). Surveillance studies across North America have reported SARS-CoV-2 infections among WTD in Canada and the northwest, midwest, southwest, and northeast regions of the United States, thus suggesting that SARS-CoV-2 infection in WTD was occurring nationally (Bevins et al., 2023, Chandler et al., 2021, Cool et al., 2022, Feng et al., 2023, Hale et al., 2022, Kotwa et al., 2022, Kuchipudi et al., 2022, Palermo et al., 2022, Palmer et al., 2021, Roundy et al., 2022, Vandegrift et al., 2022). Reverse transcriptase polymerase chain reaction (RT-qPCR) methods have been utilized to detect WTD SARS-CoV-2 infections but are limited by a short viral detection window (Hale et al., 2022, Pickering et al., 2022). The persistence of antibody long after infection favors serosurveillance as the preferred method to detect evidence of SARS-CoV-2 infection in WTD and other species (Chandler et al., 2021, Zhao et al., 2021). Serosurveillance studies have two major challenges: (1) physically collecting serum from WTD and other wild animals requires expertise, and (2) obtaining representative sample sizes to detect infection is labor intensive. These shortcomings warrant development of improved and/or new WTD SARS-CoV-2 detection methods and surveillance strategies.
The objectives of this study were to conduct surveillance for SARS-CoV-2 infection in Mississippi WTD using traditional methods, to test the efficacy of a new SARS-CoV-2 antibody detection strategy using kidney tissue homogenates, and to identify potential human-to-WTD transmission routes using camera trap data. Kidneys were chosen as a potential sample source to determine the feasibility of collecting organs from deer hunters without the need for any specialized sample collection materials. SARS-CoV-2 antibody was detected in kidney tissue and serum samples of WTD as the first of evidence of infection by this virus among WTD across multiple counties in Mississippi during the 2021–2022 and 2022–2023 deer hunting seasons. In addition, camera trap data documented WTD activity in multiple urban locations, including grazing behavior near wastewater effluents during the SARS-CoV-2 pandemic, thus raising the possibility of exposure to human wastewater as a potential route of indirect transmission of SARS-CoV-2 to WTD.
Methods
White-tailed deer sample collection
Kidney tissues, serum samples, and nasal swabs were collected from Mississippi hunter-harvested deer during the 2021–2022 hunting season in Adams, Amite, Franklin, Jones, Hinds, Holmes, Warren, and Yazoo counties. In 2022–2023, samples were collected from WTD during the Mississippi hunting season in Claiborne, Copiah, DeSoto, George, Hinds, Holmes, Issaquena, Jones, Kemper, Madison, Perry, Tishomingo, Warren, and Yazoo counties. The Mississippi Department of Wildlife, Fisheries, and Parks, Deer Program biologist collected samples for this study beyond the 2022–2023 hunting season ending in May of 2023. Kidney samples were collected from November 2021 to January 2022 for the 2021–2022 cohort and from November 2022 to May 2023 for the 2022–2023 cohort. Blood samples were collected in 15-mL tubes via venipuncture from the cerebral venous and arterial systems for ease of access and to prevent gastrointestinal fluid contamination. Samples were collected during the same time period as kidney samples and were allowed to clot at ambient temperature, and serum was collected after whole blood centrifugation. The nasal cavities of hunter-harvested deer were swabbed using nasopharyngeal polystyrene flocked coated swab (StaLab, McKinney, TX) during the month of January 2022 for the 2021–2022 cohort and from November 2022 to May 2023 for the 2022–2023 cohort. The swabs were placed into sterile 15-mL tubes, containing 3.0 mL of phosphate-buffered saline (PBS). To prevent cross contamination of samples, fresh gloves were used to handle each deer. All samples were labeled by date, location, and sex and then stored at −20°C until further analysis for antibody and/or SARS-CoV-2 viral RNA. In addition to results of testing samples from WTD, the result of testing human samples for SARS-CoV2 was obtained from the Mississippi State Department of Health (https://msdh.ms.gov/msdhsite/_static/14,0,420.html) during the same time (2021–2023) period as the samples were collected from WTD.
SARS-CoV-2 RT-qPCR assay
RNA was extracted from separate WTD nasal swab sample solutions using an Applied Biosystems™ MagMAX™ Viral/Pathogen II (MVP II) Nucleic Acid Isolation Kit (Waltham, MA) according to the manufacturer’s instructions. This method was validated for use in testing human clinical samples for SARS-CoV-2 in the Molecular Pathology Laboratory at the University of Mississippi Medical Center (UMMC). Probes annealing to the region of SARS-CoV-2 genome that encodes the nucleocapsid (NP) protein were used for testing nasal swab samples from WTD for SARS-CoV-2 by RT-qPCR. All primers were provided in the 2019-nCoV RUO kit purchased from Integrated DNA Technologies (Coralville, IA). This kit contained multiple primer/probe sets that targeted regions in the 5′, middle, and 3′ regions of the N gene sequence and was supported by Primer Express software version 3.0.1 (Thermo Fisher Scientific, Waltham, MA USA) (Lu et al., 2020). Three gene regions were selected including regions designated as N1, N2, and N3. The use of the primer/probe set N1 and N2 specifically detected SARS-CoV-2, and N3 universally detected all clade 2 and 3 viruses within the subgenus Sarbecovirus, including SARS-CoV, SARS-CoV-2, and bat- and civet-SARS–like CoVs. Reactions were run on an Applied Biosystems™ QuantStudio 7 Pro or QuantStudio 12K Flex RT-qPCR instrument. Samples with a cycle threshold (Ct) ≤31 were considered SARS-CoV-2 RNA-positive reaction, whereas samples yielding a Ct >31 were considered undetermined or negative reaction.
Illumina-based sequencing of SARS-CoV-2 genomes from deer
Sequencing was performed using the Illumina COVIDseq test and platform by the UMMC Molecular and Genomics Core Facility. COVIDseq is an amplicon-based next-generation sequencing (NGS) test based on 2019-nCoV primers designed to detect SARS-CoV-2 RNA (based on ARTIC multiplex PCR protocol v4, with 98 amplicons), along with internal controls consisting of 11 human mRNA targets. Each sequencing library was prepared according to the manufacturer’s protocol on a PerkinElmer Zephyr NGS workstation, and pooled samples were run on NextSeq500 using MO flow-cell. The DRAGEN COVIDseq Test App (via Illumina BaseSpace Cloud Computing Platform) was used for alignment of reads to confirm positive samples and to identify variants/clades. In silico translate was performed using a tool that allows the translation of a nucleotide (DNA/RNA) sequence to a protein sequence. (Expasy https://web.expasy.org › translate).
SARS-CoV-2 neutralizing antibody test
WTD sera samples were tested for SARS-CoV-2 neutralizing antibody in Vero E6 cells using a plaque reduction neutralization test (PRNT80) previously described (Palermo et al., 2022). Briefly, each serum sample was diluted 1:5 in Eagle’s minimum essential medium and mixed with an equal volume of 40–50 plaque-forming units (PFUs) of SARS-CoV-2 (USA-WA1/2020 strain), incubated at 37°C for 1 h, and inoculated in duplicate onto monolayers of Vero E6 cells. An agarose overlay was added to the cell monolayer, and the cells were incubated for 2 days before staining a neutral red solution. The virus dose was determined as the mean number of PFUs recorded in virus-only negative control wells. Serum samples were considered positive for SARS-CoV-2 neutralizing antibody if a 1:10 serum dilution reduced the virus dose by at least 80% in the number of PFUs of the virus dose. Each test run included known SARS-CoV-2 antibody-positive and antibody-negative sera samples.
SARS-CoV-2 enzyme-linked immunoassay
WTD sera and kidney tissue samples were tested for antibody by an indirect enzyme-linked immunoassay (ELISA). The kidney samples were prepared by removing 4 g of perfused tissue from the center of the kidney and then homogenized in sterile PBS in a 50-mL conical tube using an electric-powered homogenizer. The samples were clarified by centrifugation. Supernatants were stored at −20°C until testing. ELISA was performed as previously described (Bates et al., 2022, Tandon et al., 2020) but using anti-deer IgG conjugated to horseradish peroxidase (VWR, Radnor, PA) as a secondary antibody. Kidney homogenates and serum samples were considered as antibody positive if they had endpoint titers equal to or greater than 1:100 in the ELISA. The antibody titer was calculated as the reciprocal of the sample dilution that yielded an absorbance of 0.2 units or greater above the no-sample control wells. The validity of the performance of the ELISA for detecting SARS-CoV-2 antibody was determined by testing the 45 WTD sera samples that were collected from the 2022–2023 WTD cohort and had been tested by the PRNT80.
Surveying deer population, activity, and movement
Ten camera traps (Model #: 119270CW, Tasco Worldwide, Miami, FL) were placed over active WTD scrapes as previously described (Hearst et al, 2021; Hearst et al., 2023). Scrapes were monitored for deer activity from December 15, 2021, until January 31, 2022, at three separate urban survey sites in Hinds County, MS. Site 1 was located in a neighborhood adjacent to an undeveloped, forested area. Site 2 was located in a populated urban area between Sites 1 and 3. Site 3 was located in a heavily wooded urban area next to local schools and businesses. All neighborhoods within survey Sites 1, 2, and 3 use residential wastewater treatment plants, where septic tank wastewater effluents were spread over the ground using sprinkler systems. Unique male WTD were identified and cataloged to create profiles of each male using parameters such as antler patterns, pelage, body size, and maturity estimation; maturity estimation was based upon body size and neck girth as previously described (Hearst et al., 2021; Jacobson et al., 1997). Deer activity was determined based upon buck and antlerless counts per day and used to produce a heatmap using Office of Government Information Services (QGIS) software as previously described (Hearst et al., 2023). Deer movement was determined based upon unique buck movement between survey sites and was used to determine the population of WTD inhabiting the urban survey location. The analysis was performed using a modified version of Jacobson (Jacobson et al., 1997), where scrapes were used as alternative to bait stations to identify buck-to-doe ratios and estimate the population of WTD.
Results
Using a PRNT80, SARS-CoV-2 neutralizing antibody was detected in 67% (10/15) of serum samples collected from the 2021–2022 WTD cohort in Hinds, Holmes, and Yazoo counties and in 62% (28/45) of serum samples collected from the 2022–2023 WTD cohort in the Hinds, Madison, and Yazoo counties. The sera samples collected from both cohorts were also tested by ELISA to evaluate this assay for detecting SARS-CoV-2 antibody. The results revealed that 60% (9/15) of the 2021–2022 WTD cohort and 62% (28/45) of the 2022–2023 WTD cohort of samples were positive by this assay, thus demonstrating strong agreement between the PRNT80 and the ELISA for detecting SARS-CoV-2 antibody in the WTD serum samples
A total of 62 kidney tissue samples were collected from the 2021–2022 cohort of 62 WTD from eight Mississippi counties. Also, serum samples were collected from 13 of the deer, 9 of which contained SARS-CoV-2-specific antibodies that were detectable by ELISA (Table 1). Antibodies were detected in 8 of the 13 matched kidney homogenates; thus, the results were similar for using either serum or kidney tissue as a test sample. However, mean IgG antibody titer was 6,349 for the 8 antibody-positive serum samples and 546 for the antibody-positive kidney tissue homogenate samples or 11.6 times lower than the titer for the antibody-positive serum samples. On testing the 62 kidney homogenate samples from the 2021–2022 cohort of WTD by ELISA, 26% (16/62) were positive for SARS-CoV-2 antibody, with varying positivity rates across counties. However, 220 kidney tissue samples obtained from the 2022–2023 WTD cohort from 14 Mississippi counties, which also included samples from WTD from Hinds, Holmes, and Yazoo counties, were negative for SARS-CoV-2 antibody (Supplementary Table S1). These results differed from the serosurveillance data obtained using the PRNT80 to test serum samples that indicated 62% (28/45) of the 2022–2023 cohort sampled in Hinds, Holmes, and Yazoo counties was positive for SARS-CoV-2 antibody. The SARS-CoV-2-specific antibody cumulative seropositivity among WTD during the 2021–2022 hunting season was compared with the RT-qPCR positivity rates in humans across the whole state of Mississippi over the same time period (Fig. 1A). The SARS-CoV-2 RT-qPCR test positivity rate for humans increased in late-November of 2021 and reached a maximum of 46% by mid-January of 2022 during the Omicron variant virus wave. This peak in human test positivity rate slightly preceded the spike in WTD seropositivity rates (67%; 10/15) from mid- to late-January of 2022. The SARS-CoV-2-specific antibody cumulative seropositivity was then plotted against the RT-qPCR positivity rates in human based upon their Mississippi county locations over the same time period (Fig. 1B), revealing a correlation coefficient of 0.8393.

WTD SARS-CoV-2 antibody positivity trends and human diagnostic test positivity rates in Mississippi.
Comparison of SARS-CoV-2 IgG Antibody Titers for Kidney Tissue Homogenates and Serum Samples Obtained by an Enzyme Immunoassay (ELISA) for 13 WTD Harvested During the 2021–2022 Survey
ELISA, enzyme-linked immunosorbent assay; ND, indicates that the antibody was not detected in sera and/or kidney homogenate samples; WTD, white-tailed deer.
Nasal swabs were collected from 12 WTD of the 2021–2022 cohort and tested for SARS-CoV-2 infections using RT-qPCR. Among these samples, 25% (3/12) tested positive for SARS-CoV-2. One positive sample was collected in Hinds County and the other two were collected in Yazoo County. As shown in Supplementary Table S2, the CT values were 25 for the positive control 44 for the negative control, and the CT values were 23, 31, and 29 for the three positive deer. Nasal swabs collected from the 2022–2023 WTD cohort (n = 220), which included samples from 14 Mississippi counties, were negative for SARS-CoV-2 (Supplementary Table S1).
Although the kinetics of the seroconversion of the deer population support the hypothesis of rapid spillover from infected people to WTD, the possibility of seasonal peaks of SARS-CoV-2 infection in the deer population occurring independently of the human population could not be excluded. To address this possibility, we attempted to sequence SARS-CoV-2 genomes from the 3 positive nasal swab samples. Only about one-half of the amplicons yielded usable sequences; thus, the sequences were not sufficient to determine the strain of the virus (Supplementary Fig. S1). However, in silico translation of the amplified regions of the genome that encoded the spike protein revealed four mutations (Table 2). Three of these mutations, D614G, N764K, and D796Y, were present in the SARS-CoV-2 Omicron variant (Carabelli et al., 2023), which started circulating in the human population in Mississippi in December 2021. The three mutations, N370K, D614G, and D796Y, are surface-exposed, where the N370K mutation was found to be within the receptor-binding domain of the spike protein structure (Supplementary Fig. S2). The detection of the viral sequence bearing these mutations from the deer population in January 2022 is consistent with rapid widespread increase in the infection rate of the deer population during the peak human Omicron virus wave in Mississippi. Camera traps recorded 55,921 photos of WTD in 3 urban survey sites in Hinds County, Mississippi. These images were used to identify 35 WTD bucks and 96 antlerless does and yearling deer, for a total of 131 WTD inhabiting these urban sites (Fig. 2A). Deer activity analysis showed that WTD were moving across the urban landscape of Hinds County during the Omicron outbreak (Fig. 2B). Site 1 and Site 3 had 12 bucks common to both sites, 8 bucks were common to Site 2 and Site 3, and 2 bucks were common to all 3 sites (Table 3). At Site 1 and Site 3, WTD were observed grazing in urban neighborhoods near septic tank sprinkler heads (Fig. 2C).
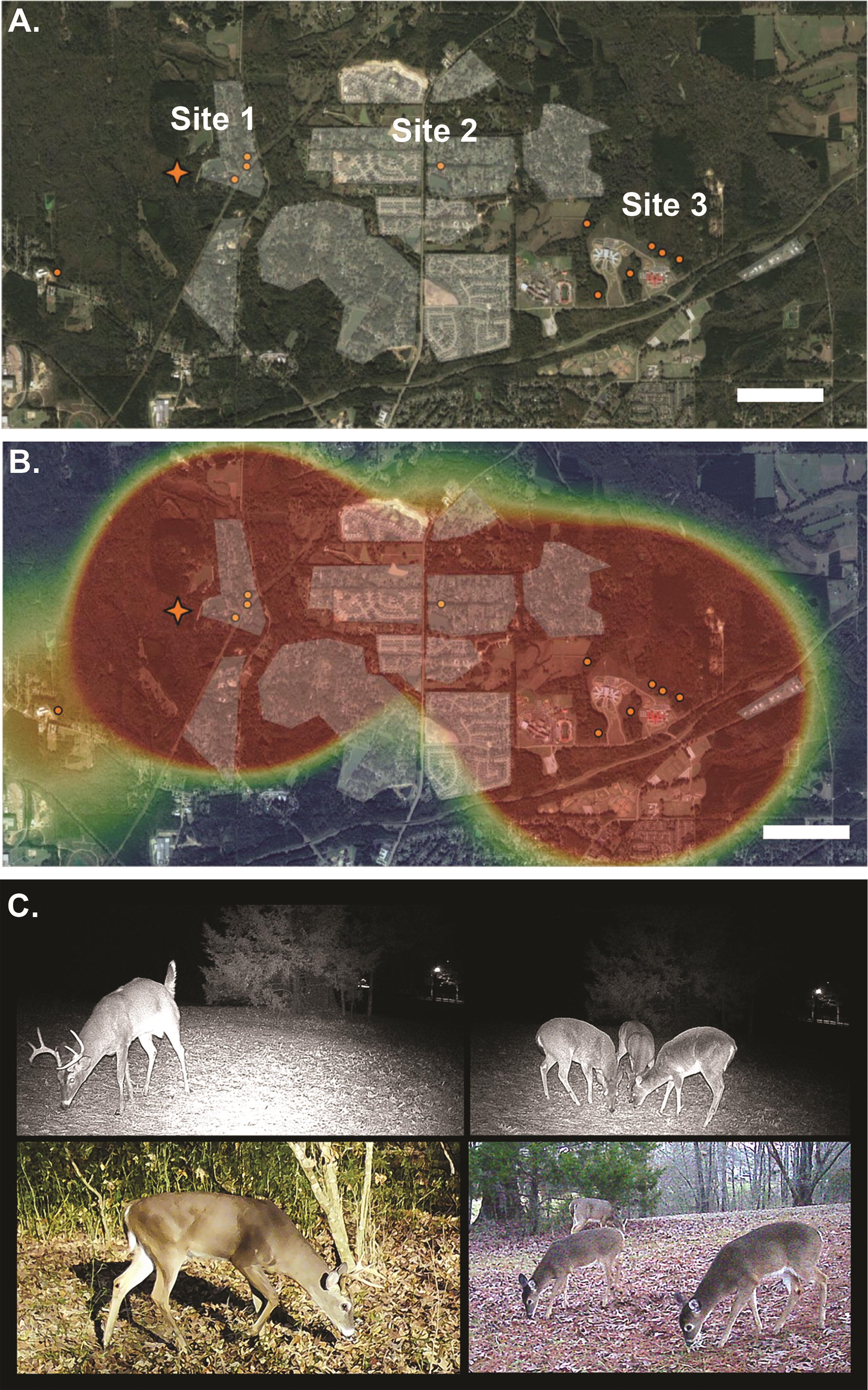
Urban WTD movement and activity during the SARS-CoV-2 Omicron wave in Hinds County, Mississippi.
Encoded Mutations in SARS-CoV-2 Genomes Recovered from Nasal Swabs of WTD
Appearance of Identifiable WTD Bucks at Multiple Observation Sites
Discussion
Although our sample size was small, it is similar to sample sizes from previously published SARS-CoV-2 WTD serosurveillance studies, and our results support a scenario of infection of the WTD population in Mississippi with SARS-CoV-2 following emergence in the human population (Chandler et al., 2021). Here, we report the highest serological positivity results in the United States to date and the first results from the Southeast region of the United States. In 2021–2022 and 2022–2023, we found seropositivity rates in Mississippi WTD to be 67% and 62%, respectively. The overall lower rate of detection of antibody in kidney homogenates using an ELISA relative to the higher rate in sera using the PRNT80 indicated that the use of kidney tissue homogenates was limited for detecting antibody and revealed that serum-based assays provided the most reliable estimate of SARS-CoV-2 antibody among WTD. In addition, given the differential sensitivities of the assays, detection of SARS-CoV-2 antibodies in the serum, but not the kidney homogenates, of deer harvested in 2022–2023 is consistent with waning antibody titers following widespread infection in 2021–2022. These data suggested that SARS-CoV-2 was circulating among WTD during the 2021–2022 survey period but became much less prevalent after the transition to endemic circulation.
Recent 2023 studies by the United States Department of Agriculture Animal and Plant Health Inspection Service (USDA APHIS) reported seropositivity of 40% in WTD across 29 states and Washington, D.C., excluding Mississippi and other states in the Southeast (Bevins et al., 2023, Feng et al., 2023). These reports, in conjunction with our data, support SARS-CoV-2 spillovers from humans into WTD population throughout the United States, including Mississippi. The seropositivity rate among the Mississippi WTD population from mid-November of 2021 until mid-January of 2022 trailed the RT-qPCR testing positivity rate in the human population and then increased abruptly, exceeding the rate in humans by the end of January of 2022. Our observation of relatively short-lived infections in humans closely preceding longer-lived antibody responses in deer supports the hypothesis of rapid spillover from infected humans to WTD and subsequent transmission within the WTD population. Although only partial sequences were obtained from the SARS-CoV-2 genome detected by RT-qPCR in nasal swabs of WTD, the results suggested that WTD were infected by the Omicron virus variant. Once infected, WTD have been reported to shed virus in their fecal matter and nasal secretions, which could be transmitted to other WTD within local social networks, quickly spreading the virus among the WTD population (Chandler et al., 2021, Hale et al., 2022, Hearst et al., 2021, Palmer et al., 2021). However, the route by which WTD were initially infected remains an open question. Identifying transmission routes is needed to develop management strategies to reduce future spillover of this zoonotic virus from humans to wildlife.
Our WTD activity data revealed that WTD move through urban neighborhoods and cohabitated indirectly with humans, where human-to-deer transmission most likely occurs. One possible transmission route resulting from this behavior is through indirect contact of WTD with contaminated wastewater. SARS-CoV-2 also causes gastrointestinal infections, where infected people can shed viral RNA in their feces, and viral RNA, but no live virus, has been detected in human sewage (Cheung et al., 2020, Natarajan et al., 2022). Septic tanks are common in rural and suburban settings, and effluents from septic tanks have the potential to release pathogens via sprinklers and runoff (Amin et al., 2020). During the SARS-CoV-2 Omicron wave in Hinds County, Mississippi (2021–2022), the use of cameras documented WTD grazing near septic tank sprinklers at multiple urban camera sites, raising the possibility of wastewater as a potential anthroponotic transmission route of SARS-CoV-2 to WTD. Although the evidence is limited, this observation warrants further study of wastewater as a possible source of SARS-CoV-2 infection of WTD. Finally, the findings of this study support the potential role of WTD in sustaining the maintenance and transmission of SARS-CoV-2, and therefore, this warrants ongoing surveillance to proactively detect the possibility of newly emerging variants that could pose new threats to human health.
Footnotes
Acknowledgments
The authors acknowledge Dr. Sarika Jain and the UMMC Molecular Pathology Laboratory for their assistance with the RT-qPCR assay. They wish to acknowledge the following Mississippi College students for their efforts in sample collection and preparation: Jeremy Evans, Caroline Armstrong, Jose Alfonso Xavier Fernandez, Chinaza Nwaiwu, Mathew Sinclair, Brandon Welch, James Lock, Breana Chastang, Ridge Futral, Haleigh Hassinger, Bethanie Prine, Daisia Rixter, Emily Rodrigue, Lillian Sisson, Jingxuan Wan, Tyler Welch, and Charles Moore. The authors thank the Mississippi hunters who submitted samples for this study. They also acknowledge the Mississippi College Research Counsel, Bentonia Tire Services, and Walker Farm Services for their support in this study.
Authors’ Contributions
S.H., P.M.P., D.M.W., and J.T.B. conceived the study and co-wrote the article. S.H., K.C., R.I., C.Y., W.Y., E.F., J.S., and G.T.B. collected samples for the study. K.A.M. developed the homogenate ELISA protocol and analyzed samples. P.R., G.C.B., and P.M.P. analyzed samples. All authors reviewed the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
S.H. was funded by the Mississippi IDeA Network of Biomedical Research Excellence (INBRE) (P20GM103476). The work performed through the University of Mississippi Medical Center (UMMC) Molecular and Genomics Facility was supported, in part, by funds from the National Institute of General Medical Sciences (NIGMS), including the Molecular Center of Health and Disease (P20GM144041), Mississippi IDeA Network of Biomedical Research Excellence (INBRE) (P20GM103476), and Obesity, Cardiorenal and Metabolic Diseases COBRE (P30GM149404).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.