
Abstract
Background:
Severe fever with thrombocytopenia syndrome (SFTS) is an emerging tick-borne disease caused by Bandavirus dabieense. Initially identified in China, this disease has spread throughout Asian countries via tick bites and animal-to-human transmission. However, reports of the prevalence of SFTS virus (SFTSV) in cattle in Korea are lacking. This study aimed to investigate SFTSV infections in grazing cattle in the Republic of Korea (ROK).
Materials and Methods:
In total, 845 grazing cattle serum samples were collected over 2 years (2019 and 2020) in the ROK, and viral RNA was extracted using a kit. One-step RT-nested PCR was performed to amplify the S-segment of SFTSV. Positive serum samples were used to isolate SFTSV in Vero E6 cells, and the full sequences were analyzed. A phylogenetic tree was constructed using the maximum-likelihood method with MEGA X. In addition, immunoglobulin G antibodies against SFTSV were investigated using an enzyme-linked immunosorbent assay.
Results:
Here, 4.0% of serum samples (34/845) were positive for SFTSV S-segments, and one virus isolate was cultured in Vero E6 cells. Phylogenetic analysis based on the partial S-segment classified 4 SFTSV isolates as the B-2 genotype, 9 as the B-3 genotype, 18 as an unclassified B genotype, and 3 as the D genotype. One cultured virus was classified as the B-2 genotype based on SFTSV L-, M-, and S-segments. Antibody detection results showed that 21.1% of serum samples (161/763) were positive for SFTSV.
Conclusion:
To the best of our knowledge, this is the first study performed to identify the prevalence of SFTSV in grazing cattle in the ROK. Our findings indicate the necessity for more intensive and continuous SFTSV monitoring, not only in cattle but also in other animals, to comprehend the genetic diversity of the virus and its potential eco-epidemiological impact on human health.
Introduction
Severe fever with thrombocytopenia syndrome (SFTS) is a tick-borne infectious disease caused by the SFTS virus (SFTSV) (Bao et al., 2011), of the Bandavirus genus, Phenuiviridae family, and Bunyavirales order, which has been officially named Bandavirus dabieense by the International Committee on Taxonomy of Viruses (https://talk.ictvonline.org) in their 2022 taxonomic update. SFTSV is an enveloped virus with a single-stranded, negative-sense RNA genome, segmented into large (L), medium (M), and small (S) sections (Yu et al., 2011). The L-segment, comprising 6,368 nucleotides, encodes an RNA-dependent RNA polymerase (RdRp) via an open reading frame. The M-segment, containing 3,378 nucleotides, encodes a glycoprotein precursor, which is later cleaved into Gn and Gc glycoproteins within the cell. The S-segment, an ambisense RNA of 1,746 nucleotides, encodes two proteins in opposite orientations, a nonstructural protein (NSs) and a nucleocapsid protein (N) (Yu et al., 2011). Various studies have documented at least six distinct genotypes of SFTSV (genotype A–F) across East Asian nations, with the prevalence of each varying among these countries (Fu et al., 2016; Yoshikawa et al., 2015; Yun et al., 2020).
The primary clinical manifestations of SFTSV infection include hemorrhagic fever, thrombocytopenia, leukocytopenia, and elevated hepatic enzymes (Liu et al., 2014; Yasukawa and Azuma, 2019). These can become fatal, with mortality rates ranging from 5.0% to 19.3% in China, Japan, and the Republic of Korea (ROK) (Crump and Tanimoto, 2020; Huang et al., 2019; Kim et al., 2022). In addition, SFTSV infections have been documented in dogs and cats, with various disease signs, including fever, anorexia, vomiting, leukopenia, and thrombocytopenia (Han et al., 2022a; Han et al., 2022b; Ishijima et al., 2022; Matsuu et al., 2019; Park et al., 2019). Furthermore, animal-to-human SFTSV transmission has been documented (Oshima et al., 2022; Tsuru et al., 2021), increasing public health concerns and indicating a potential risk of transmission from other animal species.
This disease was first identified in China in 2009 (Yu et al., 2011), followed by the ROK, Japan, (Kim et al., 2013; Takahashi et al., 2014), and other Asian countries, such as Vietnam, Thailand, Pakistan, Myanmar, and Taiwan (Lin et al., 2020; Ongkittikul et al., 2020; Rattanakomol et al., 2023; Tran et al., 2019; Win et al., 2020; Zohaib et al., 2020). The expanding geographical distribution of SFTSV has increased concerns regarding its global prevalence. Therefore, many researchers have reported its prevalence in not only humans but also ticks and companion, livestock, and wildlife animals (Ishijima et al., 2023, Kimura et al., 2018; Kirino et al., 2022; Wang et al., 2015). As in other countries, many reports have documented SFTSV prevalence and infection rates in various animals, including ticks, dogs, cats, horses, goats, poultry, and wild animals (Chae et al., 2017; Han et al., 2023; Kang et al., 2020; Kang et al., 2018a; Lee et al., 2018; Oh et al., 2016; Rim et al., 2023), but there are few reports of its prevalence in cattle in Korea (Lee et al., 2017; Lee et al., 2016). Cattle interact closely with humans and are significant contributors to zoonotic pathogen transmission (Klous et al., 2016), and a previous study reported a correlation between the cattle breeding density and human SFTSV infections (Liu et al., 2015), suggesting the importance of cattle when investigating SFTSV. Therefore, it is imperative to include cattle when monitoring SFTSV transmission. This study comprises an investigation of the prevalence, isolation, and molecular characterization of SFTSV in grazing cattle within the ROK.
Materials and Methods
Ethics statement
This study was approved by the Seoul National University Institutional Animal Care and Use Committee (IACUC no. SNU-190524-2-1) and Jeonbuk National University Institutional Biosafety Committee (IBC no. JBNU2022-03-002) and performed in strict accordance with the recommendations of the national guidelines.
Sample collection
To investigate SFTSV prevalence in cattle, blood samples were taken from grazing cattle raised in four provinces in the ROK, Gyeonggi-do, Gyeongsangbuk-do, Incheon Metropolitan City (Incheon), and Jeollanam-do, between 2019 and 2020. Samples were transported to the laboratory at 4°C. Upon arrival, they were centrifuged at 3,000 rpm to obtain serum, which was stored at −80°C until further analysis.
Detection of SFTSV S-segment
Viral RNA was extracted from 200 μL aliquots of serum using the Gene-spin Viral DNA/RNA Extraction Kit (iNtRON Biotechnology), as per the manufacturer’s guidelines, which was stored at −80°C until further use. Subsequent testing of each RNA sample was conducted using nested RT-PCR to detect the S-segment. The primary PCR was performed in a 30 μL reaction mixture containing 1 μL (10;pmol) of each forward (5′-CATCATTGTCTTTGCCCTGA-3′) and reverse primer (5′-AGAAGACAGAGTTCACAGCA-3′), 4 μL of extracted RNA, 9 μL of Tris-EDTA (TE) buffer, and 15 μL of one-step RT-PCR premix (Solgent) (Yoshikawa et al., 2014). The reaction began with a 30 min step at 50°C and a 15 min denaturation at 95°C, followed by 40 cycles of 20 s at 95°C, 40 s at 52°C, 30 s at 72°C, and a final 5 min extension at 72°C. Nested PCR was performed in a 30 μL reaction mixture containing 1 μL (10;pmol) of each forward (5′-AAYAAGATCGTCAAGGCATCA-3′) and reverse primer (5′-TAGTCTTGGTGAAGGCATCTT-3′), 1 μL of the primary PCR product, 12 μL of TE buffer, and 15 μL of the HiPi PCR premix (Biofact) (Oh et al., 2016). Nested PCR comprised 25 cycles, each consisting of 20 s at 94°C, 40 s at 55°C, and 30 s at 72°C. Secondary PCR products were analyzed via gel electrophoresis with a 1.2% agarose gel. Bands were visualized through poststaining with the EcoDye™ Nucleic Acid Staining Solution (Biofact) and finally purified with the DNA Gel Extraction Kit (Bionics). PCR partial S-segment (346 bp) amplicons were directly sequenced using dideoxy termination with an automatic sequencer (3730xl Capillary DNA Analyzer; Applied Biosystems).
Cell culture and virus isolation
Vero E6 cells were prepared to isolate SFTSV from positive serum samples. Cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM; Sigma-Aldrich) with 10% heat-inactivated fetal bovine serum (FBS) and antibiotics (penicillin/streptomycin or kanamycin) at 37°C. SFTSV-positive samples from cattle were selected for virus isolation. Briefly, Vero cells were adjusted to 1 × 105 cells/mL in 15 mL of DMEM (Sigma-Aldrich) with 10% heat-inactivated FBS and antibiotics (penicillin/streptomycin or kanamycin) and seeded in a T75 flask (Sumitomo Bakelite). When cells reached 80–90% confluence, the cell culture medium was changed to DMEM with 2% heat-inactivated FBS and antibiotics (penicillin/streptomycin or kanamycin) with 200 μL of SFTSV-positive serum after three PBS washes. Cells were cultured for 4–5 days after inoculation. Detection of a cytopathic effect and RT-PCR targeting the S-segment were performed to confirm virus isolation. Virus culturing was performed for three times. For the molecular characterization of final virus isolates, viral RNA was extracted from infected cells using QIAamp Viral RNA Mini Kits (QIAGEN) according to the manufacturer’s instruction. cDNA was synthesized using the PrimeScriptTM first-strand cDNA synthesis kit (TaKaRa) according to the manufacturer’s instructions. The three genome segments were amplified via RT-PCR using the primer pairs listed in Supplementary Table S1. The 5′ and 3′ sequences of each SFTSV were confirmed via rapid amplification of the cDNA ends using PCR. The genome sequences covering all three segments were generated via de novo assembly using DNAStar SeqMan version 7.1 (Lasergene).
Serological investigation of SFTSV antibody
To detect SFTSV antibodies in cattle serum samples, an indirect enzyme-linked immunosorbent assay was performed, as previously described with slight modifications (Han et al., 2023; Rim et al., 2023). Ninety-six-well plates (Thermo Scientific) received a coating of 100 ng/well of purified recombinant N and were left overnight at 4°C. For the recombinant protein production, the N gene from an SFTSV isolate was inserted into the pET28a(+) vector (Novagen), followed by transfection into Escherichia coli BL21 cells. Postinduction with 0.1 mM isopropyl β-D-thiogalactoside, HisTrap HP columns (GE Healthcare) were used for purification of the recombinant protein as per the manufacturer’s guidelines. The plates, postcoating with recombinant N, were blocked using 5% skim milk in Phosphate-Buffered Saline with Tween 20 (PBST) with 0.05% Tween 20 for 2 h at room temperature. This was followed by an 1-h incubation at room temperature with 1 μL/well of bovine serum in 5% skim milk. Detection of binding antibodies was achieved with horseradish peroxidase-labeled goat anticow immunoglobulin G (ab102154, Abcam). A solution with 3,3′,5,5′-tetramethylbenzidine was used for color development, and the reaction was allowed to proceed for 10 min at room temperature. The reaction was halted using 2 N hydrogen sulfide. Optical density readings were taken at 450 nm using a microplate reader (BioTek Instruments). The cutoff value was established as the mean absorbance of 30 negative control bovine serum samples plus three standard deviations (SD) (mean absorbance + [3 × SD]) (Yoo et al. 2016).
Molecular characterization and phylogenetic analysis
Sequences were subjected to a Basic Local Alignment Search Tool analysis with the National Center for Biotechnology Information (NCBI) database. Sequence alignment was accomplished using Clustal X (Version 2.0), followed by examination based on a similarity matrix. Phylogenetic analysis was performed using the maximum-likelihood method, based on the Kimura two-parameter model and the used nucleotide alignments. Bootstrap analysis was conducted with 1,000 replicates using MEGA software X.
Results
Detection of SFTV S-segment and phylogenetic analysis
In total, 845 cattle serum samples (300 in 2019; 545 in 2020) were collected (Table 1). Regarding regional differences, in Gyeonggi-do, 201 serum samples were collected in 2020; in Gyeongsangbuk-do, 251 samples were obtained, with 107 in 2019 and 144 in 2020; in Incheon, 104 serum samples were collected in 2020; and in Jeollanam-do, 289 samples were secured, 193 in 2019 and 96 in 2020. All cattle were older than 7 months; however, some ages were not recorded. Of the 845 samples, 34 were SFTSV-positive (4.0%, 34/845) (Table 1). In 2019, six samples, among all total tested (2.0%, 6/300), were positive in Jeollabuk-do. In 2020, positive samples included three from Gyeonggi-do (1.7%, 3/181), one from Gyeongsangbuk-do (0.7%, 1/144), 22 from Incheon (21.2%, 22/104), and two from Jeollanam-do (2.1%, 2/96), with total infection rates of 5.1% (28/545). Nucleotide sequences were submitted to the GenBank database under accession numbers OR987939–OR987972.
Description of SFTSV Antigen and Antibody Detection from Grazing Cattle in the Republic of Korea
—, Not calculated.
ELISA, enzyme-linked immunosorbent assay; SFTSV, severe fever with thrombocytopenia syndrome virus.
An analysis of the 34 sequences obtained showed homology ranging from 94.2% to 100%. In addition, for the genotype analysis, comparisons were made with the genetic sequences of each genotype previously reported in the NCBI database. The phylogenetic analysis revealed that among the sequences obtained, four were classified within SFTSV genotype B-2, nine within genotype B-3, and three within genotype D (Fig. 1). Interestingly, 18 sequences were categorized within SFTSV genotype B, yet they formed a distinct cluster, diverging from other sequences classified as the same genotype (Fig. 1).
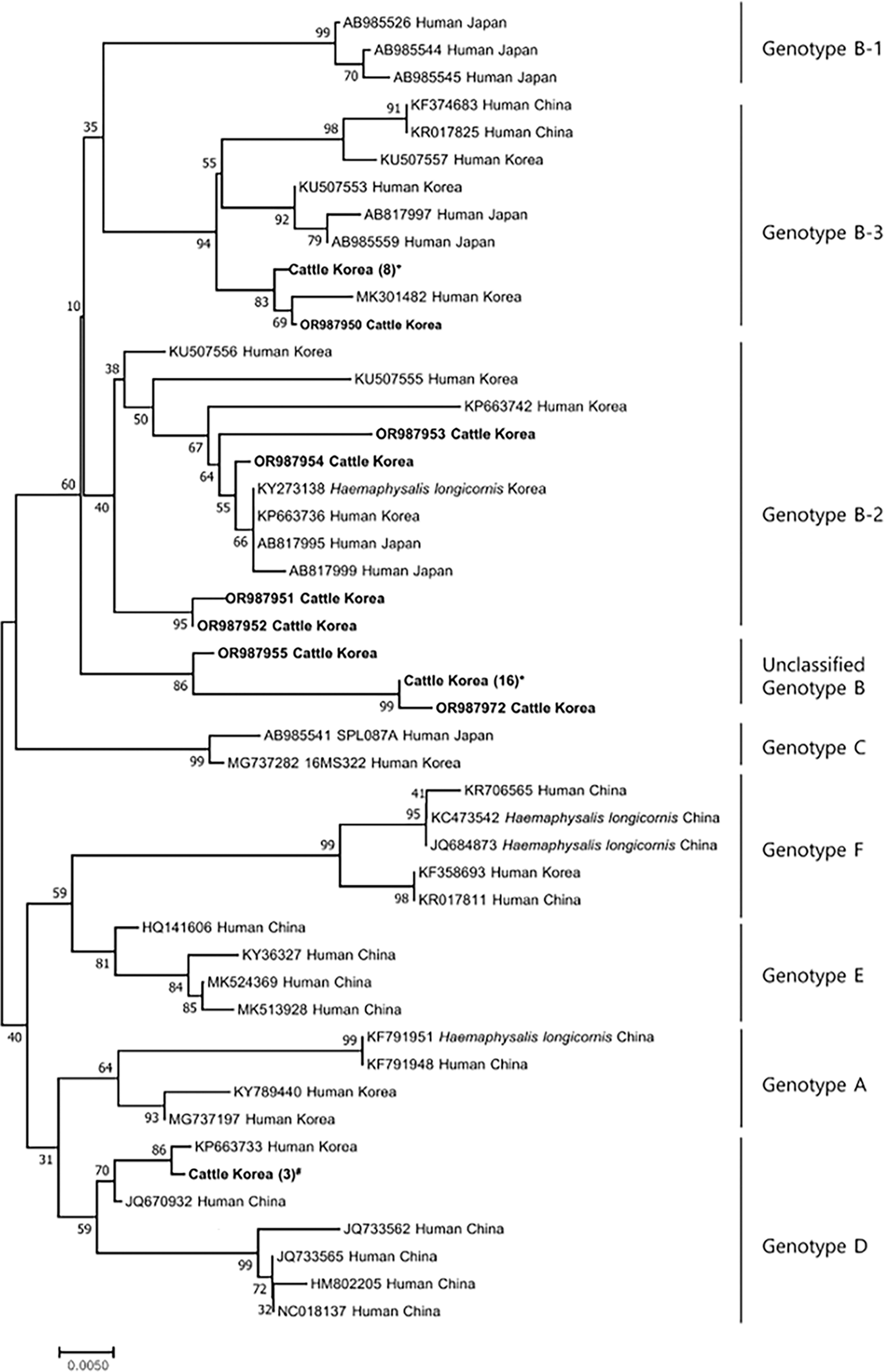
Phylogenetic tree and genotypes of SFTSV based on analysis of partial sequences (346 bp) of the small (S) segment. The sequences identified in the current study are indicated in bold. The maximum-likelihood method was used to construct the phylogenetic tree based on the Kimura two-parameter model (1,000 bootstrap replicates). The percentage of trees in which associated taxa clustered is shown next to the branches. Bold letters indicate the sequences from serum samples positive for SFTSV in this study. The number inside the brackets indicates the identical sequences. The submission numbers for identical sequences were described as follows: +OR987942-987949; *OR987956-987971; #OR987939-987941. SFTSV, severe fever with thrombocytopenia syndrome virus.
SFTSV isolation from cattle and whole-genome analysis
Among 34 SFTSV-positive serum samples, only one virus was cultured in Vero E6 cells. After three passages, SFTSV was identified via RT-PCR and subjected to full-sequence analysis. L-, M-, and S-segment lengths of the isolate were 6,368, 3,378, and 1,746 nucleotides, respectively, and the guanine and cytosine (G + C) content of each segment was 48.4%, 49.3%, and 49.1%, respectively. Nucleotide sequences obtained were submitted to the GenBank database under accession numbers OR987936 (L-segment), OR987937 (M-segment), and OR987938 (S-segment). Through phylogenetic analysis, the SFTSV isolate in this study clustered with B-2 for all segments (Fig. 2). A comparative analysis of each segment was performed against the reported SFTSV B-2 genotypes from various origins in the NCBI database (Table 2). Among isolates, our SFTSV isolate showed the highest similarity with SFTSV isolates from humans in Korea, exhibiting 100% homology for the L-segment, 99.8% for the M-segment, and 100% for the S-segment.
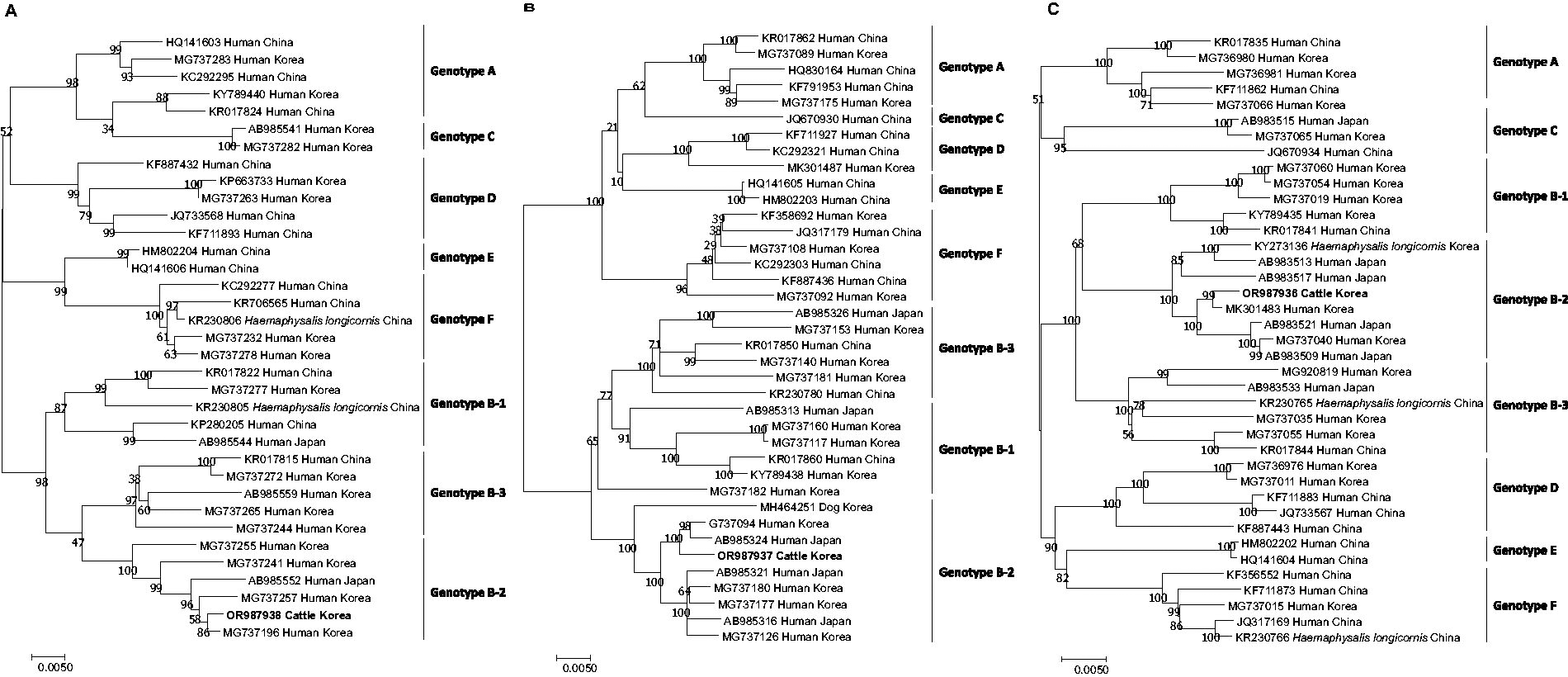
Phylogenetic tree and genotypes of SFTSV based on analysis of the full sequence of the L
Nucleotide Identities of SFTSV Isolates in This Study Compared with Different SFTSV Genotype B-2 from Various Origins
Serological investigation of SFTSV antibody
Among 845 samples, 763 available serum samples were subjected to a serological SFTSV antibody investigation. Among the 763 samples tested, 161 samples were SFTSV-positive (21.1%, 161/763) (Table 1). Specifically, Gyeonggi-do had an 11.0% infection rate (20/181) in 2020; Gyeongsangbuk-do had a total infection rate of 43.2%, with 39/46 positive samples in 2019 (84.8%) and 43/144 in 2020 (29.9%); Incheon had a 10.7% infection rate (11/104) in 2020; and Jeollanam-do had a total infection rate of 16.6%, with 30/193 positive samples in 2019 (15.5%) and 18/96 in 2020 (18.8%) (Table 1).
Discussion
Here, the SFTSV prevalence in grazing cattle in the ROK was investigated by detecting corresponding antigens and antibodies. The overall infection rate based on antigens was 4.0% (34/845). Comparing by region, Incheon exhibited a high infection rate of 21.2%, whereas that in other regions, it ranged between 0.4 and 2.8%. Unfortunately, a further investigation of the causes of the abnormally high infection rate in Incheon was not possible. There may be differences in tick density and virus infection rates depending on the pastures in different regions, and the timing of sample collection may also vary. However, this study could not further investigate the regional differences in results, such as the high antigen detection rate in Incheon and the high antibody detection rate in Gyeongsangbuk-do. Few studies have been performed to detect SFTSV antigens in cattle in other countries; however, much higher infection rates have been reported, specifically 23.2–27.5% in Taiwan (Kuan et al., 2023) and 18.75% in China (Huang et al., 2019). This could be due to differences in national breeding environments, farming practices, and cattle ages.
We focused on grazing cattle as SFTSV hosts in the ROK. However, the prevalence of SFTSV in various animals in the ROK has been reported. Reported infection rates include 0.5–17.5% in cats (Hwang et al., 2017; Kang et al., 2020; Lee et al., 2017), 0.2–3.1% in dogs (Han et al., 2022b; Lee et al., 2017), 2.0–2.4% in goats (Kang et al., 2018a; Yu et al., 2018), 1.7% in domesticated pigs (Kang et al., 2018b), 2.5% in various poultry species (Rim et al., 2023), and 2.3–5.2% in wild animals (Lee et al., 2020; Oh et al., 2016; Rim et al., 2021), similar to the infection rates in cattle reported herein. Currently, owing to a paucity of such research, the details regarding natural SFTSV infections in animals and natural hosts remain unclear. Whereas raising domestic animals is known to be related to a risk of human SFTSV infection, further research on animal-to-animal transmission and virus amplification in animals is needed to enhance the overall understanding of natural SFTSV infection mechanisms (Sun et al., 2015).
A phylogenetic analysis revealed four sequences classified within the SFTSV genotype B-2, nine within genotype B-3, and three within genotype D (Fig. 1). Interestingly, 18 obtained sequences were categorized within SFTSV genotype B, yet they formed a distinct cluster. Recent studies have classified SFTSV genotypes as A–F, with genotype B identified as the most prevalent in the ROK (Yun et al., 2020). Genotype B was previously divided into three subgenotypes (B-1, B-2, and B-3) (Yun et al., 2020). However, in this study, the isolated strains formed a distinct cluster, separate from the three previously known subgenotypes. Another previous study in the ROK reported six subgenotypes of SFTSV genotype B, suggesting that this virus is continually undergoing genetic mutations (Seo et al., 2021). However, discussions on SFTSV genotyping are ongoing (Liu et al., 2023), and other than B-1, B-2, and B-3, others remain unclassified in this study. In this study, we used only a portion (346 bp) of the S segment of SFTSV for genotype analysis. However, for more accurate genotyping, it is essential to analyze the whole genome. Consequently, there were limitations in the genotyping conducted in this study. Further research is needed to confirm the types and pathogenicity of these additional subgenotypes.
Among 34 positive serum samples, only one virus was successfully cultured in Vero E6 cells, and whole-genome and phylogenetic analyses revealed that this cultured SFTSV clustered with the B-2 genotype based on all viral segments. SFTSV has a wide host range and has been reported to exhibit similar genetic similarities regardless of the host (Han et al., 2023; Lee et al., 2018). Therefore, understanding the genetic characteristics of these isolated viruses is crucial for assessing their risks. Since the genetic variation of SFTSV has been recently reported, resulting in different biological characteristics, pathogenicity, and virulence, whole-genome analysis of SFTSV has become increasingly important (Lee et al., 2023; Liu et al., 2023). Furthermore, considering recent findings highlighting the effect of genetic variations in the L-segments on viral replication and virulence, the significance of conducting comprehensive investigations into virus quasi-species has been emphasized (Jeon et al., 2023). Consequently, this increases the necessity of further whole-genome studies in SFTSV research to attain a deeper and more nuanced understanding of transmission and pathogenicity. To our knowledge, this is the first report of a cultured SFTSV isolated from cattle in the ROK.
To identify SFTSV exposure, serological antibodies were evaluated, indicating an overall infection rate of 21.1% in cattle. There have been many investigations of SFTSV antibodies, with reported infection rates of 14.4% in goats (Yu et al., 2018), 13.9% in dogs (Lee et al., 2018), 51.0% in horses (Han et al., 2023), 28.8% in wild boars (Rim et al., 2021), and 30.3% in various poultry (Rim et al., 2023). These varying infection rates could be attributed to the differing probabilities of the sampled animals being exposed to ticks in outdoor environments. In Korea, cattle, unlike dogs raised as pets or goats primarily reared in cages, could be more frequently exposed to outdoor conditions, but less so than wild animals. This greater exposure to outdoor environments is presumed to contribute to their relatively higher infection rates. In this study, the infection rates based on SFTSV antibodies (21.1%) were much higher than those based on the SFTSV antigen (4.0%) in cattle in the ROK. This is consistent with a previous study in China showing that 442 of 842 cattle (60.4%) surveyed were positive for an SFTSV-specific antibody, whereas the SFTSV RNA detection rate was only 4.2% (35/842) (Niu et al., 2013). Whereas no experimental infection results have been reported in cattle, studies on goats inoculated with the virus revealed brief viremia detection in only 60% of infected animals at 3 DPI (days postinoculation), with antibodies persistently found from 4 to 8 DPI, indicating a natural tendency for higher prevalence rates based on SFTSV antibodies (Jiao et al., 2015). In addition, during experimental infection of dogs with SFTSV, viremia was observed only at 3 DPI, while serum neutralization antibodies continued to increase until the end of the experiments (24 DPI), suggesting that the detection rates of antibodies could be higher compared with antigens (Park et al., 2022).
Conclusion
This investigation revealed the occurrence of SFTSV in grazing cattle in the ROK based on the detection of both antigens and antibodies. This is the first study to document the isolation and whole-genome sequencing of this virus from cattle in the ROK. These results are expected to contribute to future research to understand SFTSV transmission dynamics in cattle.
Footnotes
Acknowledgments
The authors would like to thank the veterinarians and staff of the Veterinary Service Laboratories in Gyeonggi-do, Gyeongsangbuk-do, Incheon Metropolitan City, and Jeollanam-do in the ROK for their help with blood collections.
Authors’ Contributions
J.-B.C.: Formal analysis, visualization, and writing—original draft. J.-M.R.: Investigation and methodology. Y.-K.C.: Investigation, validation, and methodology. S.-W.H.: Investigation and validation. J.-G.K.: Conceptualization, resources, and supervision. J.-S.C.: Conceptualization, funding acquisition, resources, supervision, and writing—review and editing.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This research was funded by the
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.